
中华鳖对日本沼虾幼虾捕食作用的实验研究
2016-06-14 09:42袁向阳朱卫东韦庠有王志铮
淡水渔业 2016年3期
袁向阳,朱卫东,富 裕,韦庠有,王志铮
(1.浙江海洋大学水产学院,浙江舟山 316022;2.余姚市水产技术推广中心,浙江余姚 315400)
中华鳖对日本沼虾幼虾捕食作用的实验研究
袁向阳1,朱卫东2,富裕1,韦庠有1,王志铮1
(1.浙江海洋大学水产学院,浙江舟山316022;2.余姚市水产技术推广中心,浙江余姚315400)
摘要:以平均体重为(416.67±14.21)g的中华鳖(Trionyx sinensis)为捕食者,平均全长为(18.40±0.04) mm(A)、(27.77±0.07) mm(B)和(38.90±0.07) mm(C)三种规格的日本沼虾(Macrobrachium nipponensis)幼虾为猎物,采用种间捕食作用研究方法,开展了中华鳖对日本沼虾的捕食选择性和功能反应的研究,以期为进一步优化虾鳖混养模式参数和提高日本沼虾养殖效益提供理论依据。结果表明:(1)中华鳖的捕食活动时段为18:00~6:00,主要集中于0:00~4:00,该时间段中华鳖对日本沼虾幼虾的捕食量占总捕食量的80%;(2)当日本沼虾幼虾密度为30尾/m2时,中华鳖表现出对B规格的日本沼虾幼虾更具捕食选择倾向,当密度为60尾/m2和90尾/m2时,中华鳖则表现出对A规格的日本沼虾幼虾更具捕食选择倾向;(3)中华鳖对日本沼虾幼虾的捕食功能反应属Holling-Ⅱ型,中华鳖对A、B和C三种规格日本沼虾幼虾理论上最大日捕食量分别为 169、75和70尾。
关键词:中华鳖(Trionyx sinensis);日本沼虾(Macrobrachium nipponensis)幼虾;捕食选择性;功能反应
日本沼虾(Macrobrachiumnipponensis),俗称青虾、河虾,隶属于节肢动物门(Arthropoda)甲壳纲(Crustaeea)十足目(Deeapoda)长臂虾科(Palaemonidae)沼虾属(Macrobrachium),是我国一种重要的经济淡水养殖动物。中华鳖(Trionyxsinensis)具夜间捕食习性[1]以及食性广、对水体环境适应性强等生物学特征[2]。借助中华鳖对日本沼虾的种间捕食压力,有助于改善日本沼虾养殖池塘底质、切断种内疾病水平传播途径和提高日本沼虾种质质量。目前,国内外有关捕食效应的研究多集中在陆上有害昆虫的生物防治[3-6],而有关水生生物间捕食作用的研究仅见于缢蛏(Sinonovaculaconstricta)对牟氏角毛藻(Chaeroeerosmoelleri)和青岛大扁藻(Platymonassubcordiformis)的滤食效应[7]、日本鳗鲡(Anguillajaponica)对日本沼虾的捕食效应[8]和中华鳖对泥鳅(Misgurnusanguillicaudatus)幼鱼的捕食效应[9]。本实验研究了中华鳖对三种不同规格日本沼虾幼虾的捕食效应,以期为进一步优化虾鳖混养模式参数和提高日本沼虾养殖效益提供理论依据。
1材料与方法
1.1材料
1.1.1中华鳖
购自余姚市冷江鳖业有限公司。运回实验基地后,用20 mg/L的高锰酸钾溶液消毒15 min后,立即移入若干规格为6 m×4 m×1.2 m的水泥池内暂养。待其排空体内异物后,从中选取肢体健壮、规格均匀、体色正常、无伤残的健壮个体作为实验对象,平均体重(416.67±14.21)g、平均甲长(146.27±1.94)mm、平均甲宽(121.70±1.20)mm。
1.1.2日本沼虾幼虾
来自本实验基地,暂养于若干底部直径为0.7 m、桶口直径为0.9 m、桶高为0.8 m的塑料桶内。暂养期间,每天早晚各投饲1次,日投喂量为总体重的3%~5%,其中早上投喂量占日总投料量的40%,下午占60%。暂养时间为7 d,以其能适应环境、正常摄食为准。实验时从中选取反应敏捷、规格均匀、体表光洁、健壮的个体作为实验对象,按平均全长依次分为A(18.40±0.04)mm、B(27.77±0.07)mm和C(38.90±0.07)mm三种规格。
1.1.3实验条件
实验在室内自然光照条件下进行,实验容器为1.15 m(底部直径)×1.3 m(桶口直径)×0.8 m(桶高)的白色圆形塑料桶(实验实际水位约0.7 m),实验持续时间均为24 h,即以当天6:00作为实验开始时刻,以次日6:00作为实验结束时刻,实验期间全程充气。实验用水为经2 d自然曝气的自来水,水温27.1~30.5 ℃。
1.2试验方法
1.2.1中华鳖对日本沼虾幼虾的捕食选择性实验
日本沼虾幼虾设定3种不同密度:30尾/ m2、60尾/ m2和90尾/m2,每种密度按三种不同规格的不同配比进行组合(表1),共21个组合,每个组合重复3次。日本沼虾幼虾放养24 h后,每一个实验组均放入1尾中华鳖。连续观察中华鳖和日本沼虾幼虾间的捕食与逃避行为,24 h后统计各实验组中幼虾的存活数。
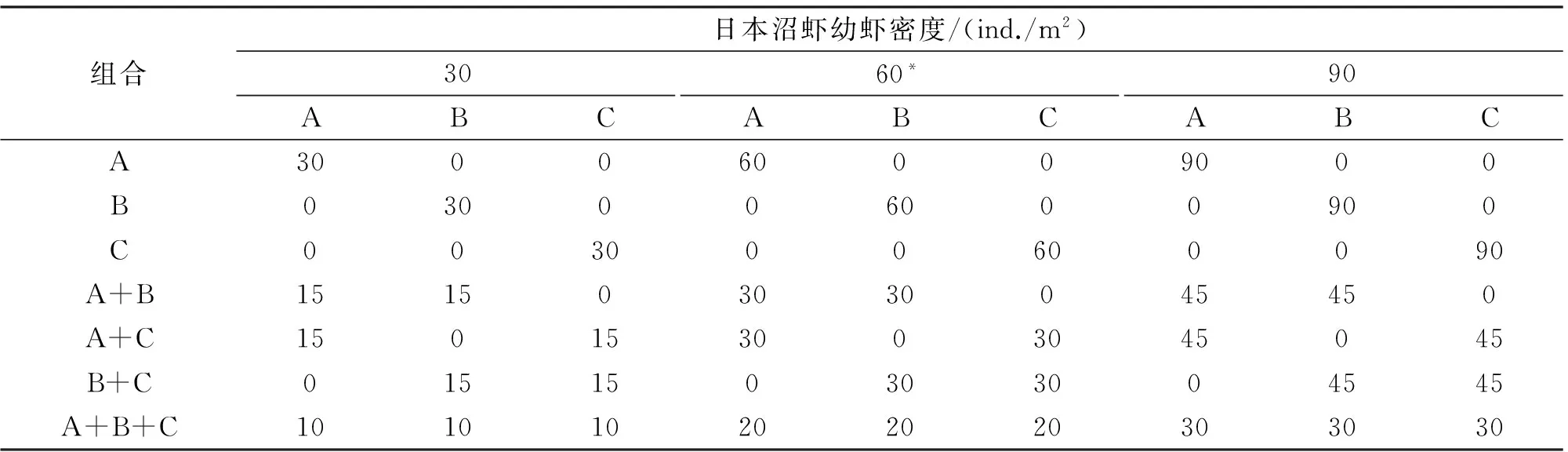
表1 日本沼虾幼虾密度及其不同规格间的组合方式
注:A、B、C分别表示平均全长为(18.40±0.04) mm、(27.77±0.07) mm、(38.90±0.07) mm的日本沼虾幼虾,下同;*表示以长江流域日本沼虾虾苗双季主养最高放养密度8.0×105ind./hm2为参照[10],设定实验组日本沼虾幼虾的数量。
1.2.2中华鳖捕食日本沼虾幼虾的功能反应实验
每一种规格日本沼虾幼虾的初始数量均按等差间距法,依次设置10、20、30、40、50、60、70、80和90尾九个梯度,共27个梯度组合,每一组合均重复3次。日本沼虾幼虾放养24 h后,每一实验组均放入1尾中华鳖。24 h后统计各实验组中日本沼虾幼虾的存活数。
1.2.3数据处理
整理实验所得数据,利用SPSS 19.0计算出日本沼虾幼虾被捕食量的平均值、标准误,同时借助LSD进行组内和组间差异性检验。在此基础上,分别计算出日本沼虾幼虾被捕食选择系数、日本沼虾幼虾的被捕食数量和日本沼虾幼虾的被捕食率,并对日本沼虾幼虾的被捕食数量进行Holling圆盘方程[11]拟合。
日本沼虾幼虾被捕食选择系数(S)[12]:
日本沼虾幼虾的被捕食数量(Na):
Na=N0-Ne
日本沼虾幼虾的被捕食率(K):
Holling圆盘方程:
式中:Mo、Me分别表示实验初始时刻实验组中某规格日本沼虾幼虾所占的数量百分比和实验结束时刻实验组中该规格日本沼虾幼虾被捕食量占总被捕食量的数量百分比,N0、Ne、a、T、Th分别表示日本沼虾幼虾初始数量(ind.)、实验结束时刻日本沼虾幼虾存活数量(ind.)、瞬时攻击率、日本沼虾幼虾暴露于中华鳖的总时间(d)、处置时间(d)。
2结果
2.1中华鳖对日本沼虾幼虾的捕食特征
白天,多数日本沼虾幼虾不均匀地静伏于桶底,个别在水体中做无规则游动;中华鳖静栖于桶壁底部,换气间隔约1 h,每次换气持续时间约2 min,无捕食意愿和行为。18:00起,多数日本沼虾幼虾俯卧于桶壁中上层,时常沿桶壁做圈状运动,部分个体做直线运动,极少数栖息于桶底呈均匀分散状态;中华鳖沿桶底部四周爬行,偶尔停止运动并伸出脖子凝视其周围猎物,表现出捕食意愿,但捕食活动及强度均较弱,捕食成功率几乎为零。20:00以后,大部分日本沼虾幼虾位于桶底,并缓慢地做圈状或直线运动,剩余个体做无规则运动;中华鳖静呆在桶底,仅伸出脖子观望位于其近前方的猎物,偶尔表现出捕食动作,但捕食成功率极低。22:00-24:00之间,日本沼虾幼虾分布于水体各层,但多数聚集于中下层;中华鳖开始游离水体底层并捕食,但捕食量仅占整个实验周期总捕食量的10%左右。0:00-4:00,中华鳖开始追赶猎物,捕食强度剧烈,且捕食成功率相对较高,捕食量约占整个实验周期总捕食量的80%。4:00以后,中华鳖捕食行为开始减弱,捕食量约占整个实验周期总捕食量的10%。6:00以后,各实验组中的实验对象均趋于平静状态,无捕食行为出现(见图1)。观察发现,18:00至翌日6:00为中华鳖的捕食活动时间,但主要集中在0:00-4:00,该时间段中华鳖对日本沼虾幼虾的捕食量占总捕食量的80%。

图1 中华鳖对日本沼虾幼虾的捕食节率
2.2中华鳖对日本沼虾幼虾的捕食选择性
由表2可见,在相同密度条件下,放养单一规格日本沼虾幼虾的3个组合之间,中华鳖对日本沼虾幼虾的捕食量无显著差异,即日本沼虾幼虾的被捕食量与其规格无关(Ⅰ—Ⅲ)。
当密度为30尾/m2时:(1)任意两种规格等数配比组合中,除Ⅵ组合内的两种规格之间无显著差异外,其它各组合内的两种规格之间均存在显著差异(P<0.05);(2)三种规格等数配比组合中,中华鳖对B规格日本沼虾幼虾的捕食量显著大于A和C(P<0.05;(3)除Ⅳ组合外,其它组合间日本沼虾幼虾的被捕食总量均无显著差异 (P>0.05)。结果表明,在密度为30尾/m2时,中华鳖对B规格日本沼虾幼虾更具捕食倾向性。
当密度为60尾/m2和90尾/m2时:(1)任意两种规格或三种规格等数配比组合中,中华鳖对三种规格日本沼虾幼虾的总捕食量呈现A>B>C,且差异显著(P<0.05);(2)除密度为60尾/m2的Ⅶ组合和90尾/m2的Ⅳ组合外,同等密度条件下其它组合间日本沼虾幼虾的总被捕食量均无显著差异 (P>0.05)。结果表明,在密度为60尾/m2、90尾/m2时,中华鳖对A规格日本沼虾幼虾更具捕食倾向性。

表2 中华鳖对日本沼虾幼虾的捕食选择性
注:数据右上标用“a、b、c”字母标注相同密度水平下组合间的差异性(P<0.05),字母相同的表示无差异;用“m、n、p”标注组合内不同规格间的差异性(P<0.05),字母相同的表示无差异;S表示日本沼虾被捕食选择系数。
由表3可知,(1)A+B组合中,随着日本沼虾幼虾密度的增加,A规格的被捕食率随之增大,而B规格的被捕食率则呈减小趋势,且差异显著(P<0.05);(2)A+C和B+C组合中,日本沼虾幼虾的被捕食率组间均无显著差异;(3)A+B+C组合中,A规格的被捕食率随日本沼虾幼虾密度的增加呈现上升趋势,而B规格的被捕食率则呈现下降趋势,且差异显著(P<0.05),C规格的被捕食率组间无显著差异。结果进一步表明,三种规格等数配比组合中,中华鳖更倾向于捕食规格较小的日本沼虾幼虾个体。
2.3中华鳖捕食日本沼虾幼虾的功能反应



表3 不同组合中日本沼虾幼虾的被捕食率
注:数据右上标用“a、b、c”字母标注同一组合内不同规格之间的差异性(P<0.05),字母相同的表示无差异;用“o、p、q、r、s”标注所有组别间差异性(P<0.05),字母相同的表示无差异;K表示日本沼虾幼虾的被捕食率。

表4 不同密度条件下中华鳖对不同规格日本沼虾幼虾的日捕食能力
注:数据右上标用“a、b、c、d、e、f、g”字母标注同一规格组间差异性(P<0.05),字母相同的表示无差异;用“m、n、p”字母标注相同密度水平下组间差异性(P<0.05),字母相同的表示无差异。

表5 中华鳖对日本沼虾幼虾的捕食功能模型及其检验

3讨论
3.1中华鳖对日本沼虾幼虾的捕食选择性
本研究中,中华鳖表现出对捕食作用的强时效性(主要捕食时段为0:00-4:00,其捕食量约占实验周期总捕食量的80%)(图1),这与袁向阳等[9]所述的中华鳖对泥鳅幼鱼的捕食主要时段集中在 0:00-4:00的情形完全吻合,也进一步证实中华鳖有夜间觅食的生活习性[13-14]。
实验结果表明,随着日本沼虾幼虾规格的增大,其逃逸能力也同步增大,从而导致其被捕食量逐渐减少;然而,随着日本沼虾幼虾密度的增大,规格较小的日本沼虾幼虾的空间拥挤度也随之增加,同样会增大中华鳖对其捕食率。因此,在不同密度条件下,中华鳖对日本沼虾幼虾呈现出不同捕食选择性的原因,主要由于日本沼虾幼虾的自身密度及其规格的差异造成。即当日本沼虾幼虾密度低于60尾/m2时,虽然A规格日本沼虾幼虾的逃逸能力较B、C的弱,但其拥有较低的空间拥挤度使其更不易被捕食,而C规格日本沼虾幼虾凭借较强的逃逸能力而不易被捕食,故中华鳖表现出对B规格日本沼虾幼虾更具捕食选择性。当日本沼虾幼虾密度大于60尾/m2时,较高的空间拥挤度增加了中华鳖对逃逸能力弱的小规格日本沼虾幼虾的捕食机率,故中华鳖表现出对A规格日本沼虾幼虾更具有捕食选择性(表2)。
实际生产上,在日本沼虾养殖池塘内套养中华鳖的放养量仅为10~20尾/亩[15],其种间作用不足以对日本沼虾养殖产生影响,故本实验未开展中华鳖数量对日本沼虾捕食效应的影响以及中华鳖与日本沼虾种间相互干扰反应的研究。
3.2中华鳖与日本沼虾间捕食效应的生态学意义
由表2可知,在三种规格日本沼虾幼虾等数配比组合中,随着日本沼虾幼虾密度的提升,A、B规格日本沼虾幼虾的被捕食量呈明显增大趋势,而C规格的被捕食量依旧较低,且在高密度条件下,三种规格日本沼虾幼虾的被捕食量呈现:A>B>C(P<0.05)。说明在高密度养殖条件下,当日本沼虾规格存在显著差异时,中华鳖更倾向于捕食规格较小的个体。因此,在高密度养殖日本沼虾池塘内套养中华鳖时,可充分利用日本沼虾繁殖期长(每年4-9月[16-17])、生长快、繁殖力强[18-19]等特点,采取“轮捕轮放”措施,即在定期捕获达上市规格虾的同时,向池塘中投放规格较小的日本沼虾,以此来维持规格较大的个体存活数,以期达到提高日本沼虾成虾产量及其产品品质的目的。
综上所述,合理优化日本沼虾养殖池塘中日本沼虾和中华鳖的配置模式,不仅有助于提高水产养殖对象的产品质量,而且对提升日本沼虾的种质质量也具有重要意义。
参考文献:
[1]王永森,毛乾盛.中华鳖的生态习性和人工养殖[J].海洋湖沼通报,1985,(1):42-45.
[2]刘连科,王自灿,鞠广胜,等.南四湖中华鳖的习性及人工孵化[J].海洋湖沼通报,1979,(2):65-67.
[3]张安盛,于毅,李丽莉,等.东亚小花蜻成虫对西花蓟马若虫的捕食功能反应与搜寻效应[J].生态学杂志,2007,26(8):1233-1237.
[4]何辉.黄蜻稚虫对大鳞副泥鳅仔稚鱼的捕食强度及其捕食选择性[J].淡水渔业,2015,45(6):108-112.
[5]Blaeser P,Sengonca C,Zegula T.The potential use of different predatory bug species in the biological control ofFrankliniellaoccidentalis[J].Eur J Forest Res,2004,77:211-219.
[6]Fan Y Q,Petitt F L.Functional response ofNeoseiulusbarkeriHughes on two-spotted spider mite[J].Exp Appl Acarol,1994,18(10):613-621.
[7]王伟定,王志铮,杨阳,等.黑暗条件下缢蛏对牟氏角毛藻和青岛大扁藻的滤食效应[J].海洋与湖沼,2008,39(5):523-528.
[8]王志铮,杨磊,朱卫东,等.日本鳗鲡对日本沼虾的捕食效应[J].海洋与湖沼,2011,42(1):107-113.
[9]袁向阳,邹李昶,申屠琰,等.中华鳖对泥鳅幼鱼的捕食效应[J].海洋与湖沼,2014,45(1):206-211.
[10]中华人民共和国农业部.NY/T 5258-2004 无公害青虾养殖技术规范[S].北京:中国标准出版社,2004.
[11]Holling C S.Some characteristics of simple type of predation and parasitism[J].Can Ent,1959,91:385-398.
[12]周集中,陈常铭.捕食者对猎物选择性的数量测定方法[J].生态学报,1987,7(1):50-55.
[13]李林春.中华鳖生态习性的探讨[J].信阳农专学报,1995,5(2):1-6.
[14]毛盼,胡毅,罗方兴.中华鳖生物学特性及其常见传染性疾病的研究[J].湖南饲料,2013,(5):17-19.
[15]沈爱苗,陈杰,施建军,等.南美白对虾池塘套养中华鳖技术研究[J].吉林农业,2011,(6):78-79.
[16]孙建贻,张道源,谭德清,等.洪湖日本沼虾种群生长的研究[J].湖泊科学,1999,11(2):149-154.
[17]屈忠湘.青虾的生物学观察[J].淡水渔业,1990,(1):3-6.
[18]刘军,龚世园,何绪刚,等.武湖日本沼虾食性的研究[J].淡水渔业,2005,35(1):25-28.
[19]杨永林.青虾养殖初见成效[J].淡水渔业,1982,(2):9.
(责任编辑:邓薇)
The experimental study on predatory effect of Trionyx sinensis on juvenile Macrobrachium nipponensis
YUAN Xiang-yang1,ZHU Wei-dong2,FU Yu1,WEI Xiang-you1,WANG Zhi-zheng1
(1.FisheryCollegeofZhejiangOceanUniversity,Zhoushan316022,Zhejiang,China;2.FisheryTechnologyExtensionCenterofYuyao,Yuyao315400,Zhejiang,China)
Abstract:In order to optimize the culture patten of intercropping of Trionyx sinensis and Macrobrachium nipponensis,and provide theoretical basis for improving output benefits of M.nipponensis,the selective predation and functional response of T.sinensis on juvenile M.nipponensis was studied based on the predator-prey relationship theory, T.sinensis[(416.67±14.21)g(average weight)] as the predator,A(18.40±0.04) mm,B(27.77±0.07) mm and C(38.90±0.07) mm of juvenile M.nipponensis as preys.The results showed that:(1) The active interval of T.sinensis was from 18:00 to 06:00,and the hunting time focused on 00:00~04:00.(2) The predatory functional response of T.sinensis to M.nipponensis belonged to the type of Holling-Ⅱ.The theoretical daily maximal preying quantity were 169ind(A),75ind(B)and 70 ind (C),respectively.(3) When the stocking density of M.nipponensis was 30ind/m2, T.sinensis showed a stronger tendency to prey on B.When the stocking densities of M.nipponensis were 60 ind/m2 and 90 ind/m2, T.sinensis showed a stronger tendency to prey on A.
Key words:Trionyx sinensis;juvenile Macrobrachium nipponensis;predatory selectivity;functional response
收稿日期:2015-11-06;
修订日期:2016-01-11
第一作者简介:袁向阳(1988-),男,硕士研究生,专业方向为水产养殖生态环境与疾病防控。E-mail:739226829@qq.com通讯作者:王志铮。E-mail:wzz_1225@163.com。
中图分类号:S968.22
文献标识码:A
文章编号:1000-6907-(2016)03-0076-06
资助项目:宁波市农业领域科技重大攻关择优委托项目,2012C10032 号;浙江海洋学院优秀硕士学位论文培育项目[浙海院学位(2013)7号];宁波市农业创新创业项目2014C91026
猜你喜欢
当代水产(2022年7期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2022年6期)2022-06-29
江苏农业科学(2022年4期)2022-03-11
当代水产(2021年8期)2021-11-04
宝藏(2021年6期)2021-07-20
科学养鱼(2021年3期)2021-04-16
科学养鱼(2020年5期)2020-06-18
山东冶金(2019年1期)2019-03-30
海洋与渔业(2017年11期)2018-01-26
