
乳酸杆菌的表面特性及其黏附能力的研究
2016-09-12 01:05向鑫玲张英春张兰威李少慧
食品工业科技 2016年7期
向鑫玲,张英春,,*,马 放,张兰威,李少慧
(1.哈尔滨工业大学 化工学院食品科学与工程系,黑龙江哈尔滨 150090;2.哈尔滨工业大学 市政环境工程学院,黑龙江哈尔滨 150090)
乳酸杆菌的表面特性及其黏附能力的研究
向鑫玲1,张英春1,2,*,马放2,张兰威1,李少慧1
(1.哈尔滨工业大学 化工学院食品科学与工程系,黑龙江哈尔滨 150090;2.哈尔滨工业大学 市政环境工程学院,黑龙江哈尔滨 150090)
乳酸杆菌是肠道益生菌,主要通过其表面的黏附因子定殖于肠道而发挥益生作用。为了研究乳酸杆菌表面性质与黏附能力之间的关系,选择五种乳酸杆菌,分别进行自聚集能力、表面疏水性及与致病菌大肠杆菌ATCC25922和枯草芽孢杆菌K的共聚能力测定。同时,利用氯化锂和高碘酸钠分别处理乳酸杆菌后再与致病菌进行共聚作用,研究乳酸杆菌表面参与黏附的活性物质。结果表明,约氏乳酸杆菌F0421和副干酪乳酸杆菌M5-L具有良好的表面性质和黏附特性。乳酸杆菌与枯草芽孢杆菌K的共聚作用较好。同时,氯化锂和高碘酸钠处理前后,乳酸杆菌对致病菌的聚集能力有所下降,表明菌株表面蛋白及多糖参与了黏附过程。
乳酸杆菌,自动聚集,共聚作用,疏水性,表面特性
乳酸杆菌是人和动物胃肠道中的正常菌群,其在肠道中存活的数量越多、定殖的时间越长,对人及动物的健康就会越有利[1]。乳酸杆菌对肠道表面的黏附能力能够为其提供竞争优势,是其在消化道中长期存在的必要条件。而乳酸杆菌的表面特性是决定其黏附能力的重要影响因素,因此研究乳酸杆菌的表面性质对了解其益生能力具有重要的作用[2-4]。益生菌的黏附分为非特异性黏附和特异性黏附[5]。非特异性黏附通过疏水相互作用、静电作用等进行,而特异性黏附则为菌体表面的黏附素与特定受体间的结合[6]。乳酸杆菌的凝集作用与表面性质密切相关,也赋予了乳酸杆菌许多益生功能。自聚集作用使益生菌形成阻碍致病菌在肠道定殖的屏障,从而抑制致病菌对人体肠道的侵袭,有助于促进人体健康。而与肠道致病菌的凝集可使致病菌更容易从肠道环境中排出,同时,产生抑菌物质的乳酸杆菌与致病菌的凝集对于宿主抵抗致病菌感染具有重要的意义[7-10]。
本文综合研究了乳酸杆菌的非特异性黏附和特异性黏附,为全面了解乳酸杆菌的黏附特性及进一步研究乳酸杆菌的黏附机制奠定了基础。实验同时综合了两种常见的致病菌如大肠杆菌和枯草芽孢杆菌,进一步分析了不同乳酸杆菌作用不同致病菌能力的差异,为特异性的应用乳酸杆菌抑制致病菌提供了可能。本实验的目的在于,通过对益生菌的表面特性、黏附能力的测定,分析研究影响益生菌黏附作用的因素,对制定功能性食品、认识微生态学的基本规律有着重要意义。
1 材料与方法
1.1材料与仪器
副干酪乳酸杆菌M5-L(L.paracasei subp.paracasei M5-L,M5-L)源于新疆哈萨克牧民自制的马奶酒;鼠李糖乳酸杆菌J10-L(L.rhamnosus J10-L,J10-L)分离于甘肃兰州传统的发酵蔬菜汁;干酪乳酸杆菌Q8-L(L.casei Q8-L,Q8-L)分离于青海牧区自制的牦牛奶;约氏乳酸杆菌F0421(L.johnsonii F0421,F0421)分离于健康人的粪便;鼠李糖乳酸杆菌GG(L.rhamnosus GG,LGG)标准参照菌株,哈尔滨工业大学食品系保存;大肠杆菌ATCC25922(Escherichia coli ATCC25922,Ec)、枯草芽孢杆菌K(Bacillus subtilis K,K)由黑龙江省临床检测中心提供。
乳酸细菌培养基(De Man,Rogosa,Sharpe,MRS)用于培养乳酸杆菌,广东环凯微生物科技有限公司;胰酪胨大豆肉汤培养基(Trypticase Soy Broth,TSB)用于培养致病菌,广东环凯微生物科技有限公司;高碘酸钠、氯化锂、甲醇天津市科密欧化学试剂有限公司,分析纯。
UV-5100型紫外可见分光光度计上海广谱仪器有限公司;XK96-A型漩涡震荡仪姜堰市新康医疗器械有限公司;LDZX-40CI型立式自动电热压力灭菌锅上海申安医疗器械厂;SW-CJ-1F型单人双面净化工作台上海蓝豹实验仪器有限公司;TGL-16G型台式离心机上海安亭科学仪器厂。
1.2细菌培养及菌数的调整
乳酸杆菌M5-L、J10-L、Q8-L、F0421、LGG活化2代后接种于MRS 培养液,37℃厌氧培养18 h。致病菌大肠杆菌Ec和枯草芽孢杆菌K接种于TSB培养液中37℃培养16~18 h备用。
使用平板计数法得出五种乳酸杆菌菌落数均为1×109cfu/mL,两种致病菌菌落数均为2×109cfu/mL。实验中将乳酸杆菌菌落数调整为108cfu/mL,即取100 μL于MRS液体培养基中培养的菌液,离心收集菌体,使用PBS(pH7.2)洗涤2次后于1 mL PBS中重悬,将菌数调整为108cfu/mL。将致病菌菌落数调整为108cfu/mL,即取50 μL于TSB液体培养基中培养的菌液,离心收集菌体,使用PBS(pH7.2)洗涤2次后于1 mL PBS中重悬,将菌数调整为108cfu/mL。
1.3乳酸杆菌的自凝能力测定
乳酸杆菌在MRS培养基中培养24 h,离心收集菌体(4500×g,15 min),使用PBS(pH7.2)洗涤2次后于PBS中重悬,将菌数调整为108cfu/mL,4 mL等量的细胞菌悬液装入5 mL的离心管中,混匀后静置于室温下。自聚测定的时间为0、2、4、6、8 h,每次取0.5 mL的上部菌悬液,加入到1.5 mL的PBS中,混匀,测定600 nm下的吸光值,以PBS作为空白对照。凝集率(A%)计算公式[11]:
式中:A0-为初始时间时600 nm下的吸光度; At-为不同时间的吸光光度值。
1.4乳酸杆菌的表面疏水能力测定
采用微生物黏附碳氢化合物法(Bacteria Adhesion To Hydrocarbons,BATH)对菌株表面疏水特性进行测定[12]。细菌培养物经4500×g 15 min离心,然后用PBS(pH7.2)洗脱2次,重新悬浮于PBS中,调整菌数为108cfu/mL。此时从其中吸取3mL与0.5mL二甲苯涡旋混合60 s,停顿10 s后振荡60 s,室温下静置60 min分层。取下层水相,以缓冲液为空白对照,在波长600 nm处测量吸光光度值并记录。表面疏水率计算公式:
式中:A0-为与二甲苯混合前600 nm的吸光度;A-为与二甲苯混合后600 nm的吸光度。
1.5乳酸杆菌与致病菌的共聚能力测定
乳酸杆菌在MRS培养基中培养24 h,离心收集菌体(4500×g,15 min),使用PBS(pH7.2)洗涤2次后于PBS中重悬,将菌数调整为108cfu/mL。同时,致病菌采用同样的方法,重悬于PBS中,并且调整菌数为108cfu/mL。分别吸取2mL待测菌菌悬液和致病菌菌悬液于试管中,漩涡震荡10 s混匀,分别在0、2、4、6、8 h测定其在波长为600 nm下的吸光值[13]。对致病菌的凝集率通过下列公式计算:
式中:Amix-为致病菌与待测菌混合液在各个时间点的吸光值;A0-为初始时间时混合液的吸光值。
1.6乳酸杆菌表面活性分子的研究
1.6.15 mol/L氯化锂法参考文献[14]的方法,乳酸杆菌连续活化3代后,接入10 mL的MRS培养基中,培养18 h后,离心(4500×g,10 min,4℃),沉淀物用灭菌的PBS(4℃,pH7.2)洗两次,将全部沉淀物悬浮于2 mL的5 mol/L LiCl中,在37℃的摇床培养箱(200 r/min)培养60 min。然后离心收集菌体(4500×g,15 min),后于PBS中重悬,将菌数调整为108cfu/mL。同时,将致病菌采用相同的方法,调整菌数为108cfu/mL。分别吸取2 mL待测菌菌悬液和致病菌菌悬液于试管中,漩涡震荡10 s混匀,分别在0、2、4、6、8 h测定其在波长为600 nm下的吸光值。对致病菌的凝集率通过下列公式计算:
式中:Amix-为致病菌与待测菌混合液在各个时间点的吸光值;A0-为初始时间时混合液的吸光值。
1.6.2高碘酸钠法参考文献[15]的方法,乳酸杆菌连续活化3代后,接入10 mL的MRS培养基中,培养18 h后,离心(4500×g,10 min,4℃),沉淀物用灭菌的PBS(4℃,pH7.2)洗两次,将所有菌体沉淀悬浮于2 mL的0.05 mol/L的高碘酸钠溶液中(溶解于0.1 mol/L柠檬酸-磷酸缓冲液,pH4.5),在37℃的摇床培养箱(200 r/min)培养60 min。然后离心收集菌体(4500×g,15 min),后于PBS中重悬,将菌数调整为108cfu/mL。测定方法同1.6.1氯化锂处理后测定方法。
1.7数据统计
实验结果以平均值±标准偏差表示,所有实验均重复三次。采用Origin 8.6对数据进行计算并绘图。运用SPSS 18.0软件,采用独立样本t检验进行数据分析(p<0.05)。
2 结果与分析
2.1乳酸杆菌的自凝能力测定结果
菌株的自凝集是指同一种菌间相互凝集形成多细胞簇的现象。乳酸杆菌的自凝集结果见图1。五株乳酸杆菌均具有自凝能力,并且在静置凝集的8 h过程中,凝集率随着时间的延长均有所上升。其中M5-L菌株的自凝能力最强,在8 h时达到85.58%,自凝能力最差的是J10-L,8 h后的凝集率为37.46%。而在实验测定的8 h内,F0421菌株也表现出了高于标准菌株LGG的自凝能力,其凝集率仅次于M5-L菌株。
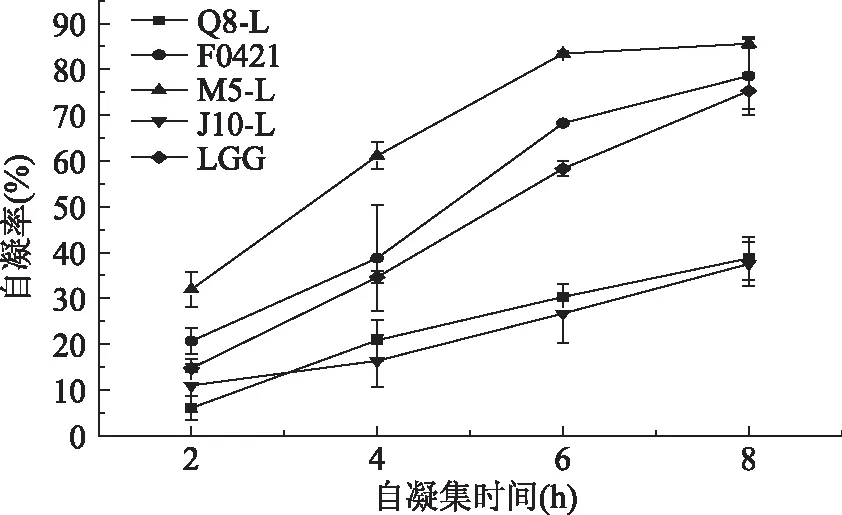
图1 五株乳酸杆菌自凝集实验结果比较Fig.1 The autoaggregation ability result of five strains of Lactobacillus
2.2乳酸杆菌的表面疏水能力测定结果
在衡量乳酸杆菌的表面疏水性能力时,通常将菌株对碳氢化合物的黏附能力作为重要的衡量指标。在本实验中将各个菌株对二甲苯的黏附能力作为评价菌体表面疏水能力的指标,菌株的疏水性如图2所示,M5-L的疏水能力最大为64.48%(p<0.05),Q8-L的疏水能力最小为3.73%,而另外三株乳酸杆菌F0421、J10-L、LGG的疏水率分别为11.87%(p<0.05)、4.80%、5.03%。其中Q8-L、J10-L和LGG三种菌株的疏水能力相差不大。
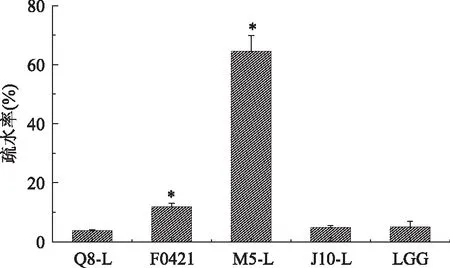
图2 五株乳酸杆菌的疏水能力测定结果Fig.2 Cell surface hydrophobicity of five strains of Lactobacillus注:*代表与标准菌株相比差异显著(p<0.05),图3~图8同。
2.3乳酸杆菌与致病菌的共聚能力测定结果
根据乳酸杆菌的自凝能力、表面疏水能力测定结果,选择自凝性和疏水性较好的菌株M5-L、F0421,并用LGG菌株作为标准菌株进行后续实验测定。测试的三株乳酸杆菌和两种致病菌的共聚实验结果见图3和图4。在室温下静置共聚8 h后,受试的3株乳酸杆菌对人体肠道致病菌均显示出凝集能力。并且对致病菌的凝集率随益生菌、致病菌的不同和共聚时间的变化而有所不同。结果表明,三种乳酸杆菌对两种致病菌的共聚能力具有时间依赖性,即随着时间的增加,凝集率也发生增加。由图3可以看出,在4 h内,标准菌株LGG对大肠杆菌的共聚作用强于M5-L和F0421,但在6~8 h内,M5-L对大肠杆菌的凝集率发生显著的增加,强于其他两种乳酸杆菌菌株,但相比于LGG没有显著性差异。而图4可以看出,在6 h内,M5-L相比其他两种菌株对枯草芽孢杆菌的共聚作用具有显著的优势,而在8 h时,F0421的共聚能力发生显著的提高,在三种乳酸杆菌中对枯草芽孢杆菌具有最好的共聚能力,且共聚效果显著高于LGG。同时,对比图3和图4可知,在各个时间内,三种乳酸杆菌均表现出与枯草芽孢杆菌(K)的共聚能力强于大肠杆菌(Ec)的情况。
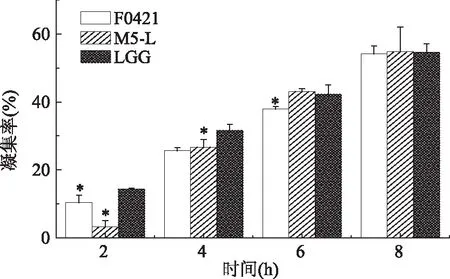
图3 三株乳酸杆菌对大肠杆菌(Ec)的凝集率Fig.3 Aggregation rate of three strains of Lactobacillus with Ec

图4 三株乳酸杆菌对枯草芽孢杆菌(K)的凝集率Fig.4 Aggregation rate of three strains of Lactobacillus with K
2.4乳酸杆菌表面活性分子的研究
研究表明,乳酸杆菌的黏附作用与其细菌的表面蛋白、多糖等存在一定关系。采用氯化锂处理菌体能够破坏乳酸杆菌表面的S-层蛋白,而采用高碘酸钠处理菌体则破坏乳酸杆菌表面的多糖。将经处理后的乳酸杆菌与致病菌进行共聚能力测定,来判断三种乳酸杆菌黏附作用的主要位点是S-层蛋白还是多糖。氯化锂处理后三株乳酸杆菌对致病菌的凝集率见图5和图6。结果表明,氯化锂处理后,静置凝聚8 h,三株乳酸杆菌对大肠杆菌(Ec)、枯草芽孢杆菌(K)仍能表现出一定的共聚能力,并且随着时间的延长,三种乳酸杆菌对两种致病菌的凝集率均不断增加。其中,M5-L菌株对Ec和K的共聚能力均为最高。通过与2.3的结果比较可知,F0421菌株经氯化锂处理后,其与两种致病菌的凝集率均发生显著降低,而氯化锂处理对于M5-L菌株及LGG菌株对两种致病菌的共聚作用没有显著影响,反而使其与致病菌的共聚能力发生了一定程度的提高。

图5 氯化锂处理后三株乳酸杆菌对大肠杆菌(Ec)的凝集率Fig.5 Copolymerization rate of three strains of Lactobacillus with Ec after LiCl treatment

图6 氯化锂处理后三株乳酸杆菌对枯草芽孢杆菌(K)的凝集率Fig.6 Copolymerization rate of three strains of Lactobacillus with K after LiCl treatment
高碘酸钠处理后三株乳酸杆菌分别对两种致病菌的凝集率见图7和图8。高碘酸钠处理后,静置凝集8 h,三株乳酸杆菌对大肠杆菌(Ec)、枯草芽孢杆菌(K)仍能表现出一定的共聚能力,并且随着时间的延长,三种乳酸杆菌对两种致病菌的凝集率均不断增加。其中,F0421菌株对Ec和K的共聚能力均为最高。通过与2.3的结果比较可知,高碘酸钠的处理使得M5-L和LGG菌株与两种致病菌的凝集率均发生显著降低,而高碘酸钠的处理相对于F0421菌株却未发生显著影响。

图7 高碘酸钠处理后三株乳酸杆菌对大肠杆菌(Ec)的凝集率Fig.7 Copolymerization rate of three strains of Lactobacillus with Ec after sodium periodate treatment

图8 高碘酸钠处理后三株乳酸杆菌对枯草芽孢杆菌(K)的凝集率Fig.8 Copolymerization rate of three strains of Lactobacillus with K after sodium periodate treatment
3 讨论
菌株的凝集包括同一菌株之间的凝集现象(如自凝集)及不同菌间的凝集现象(共凝集)。菌株凝集作用与益生菌对致病菌的抑制能力相关。很多研究结果已证明,乳酸菌的凝集能力与其黏附宿主细胞的能力和与病原菌共凝集的能力具有相关性,具有较强凝集能力的菌株可能定殖能力更好,也能更有效借助共凝集作用清除病原菌[16-17]。同时,乳酸菌与致病菌的共聚作用能够掩盖致病菌的表面物质,使致病菌不能够与宿主细胞表面的分子受体结合,进而干扰致病菌对宿主细胞的入侵作用[18]。Golowczyc等[19]分别研究了能发生共聚作用的菌株L.kefir CIDCA8321和不能发生共聚作用的菌株L.kefir CIDCA83113对Salmonella的作用,结果表明,CIDCA8321在抑制Salmonella入侵Caco-2细胞及TC-7细胞方面具有显著的效果,而在CIDCA83113却未发现这种作用。本实验通过对乳酸杆菌自凝能力及与致病菌共聚能力的测定,结果表明M5-L和F0421都具有很好的自凝能力以及与致病菌的共聚能力,且两种乳酸菌对枯草芽孢杆菌K的共聚性强于大肠杆菌(Ec),这可能与两种致病菌不同的表面结构有关。
细菌的黏附与菌体表面性质存在很大的相关性。研究菌体的表面性质不仅可以快速筛选具有黏附性质的益生菌,还可以用于判断菌体的黏附机制。研究表明,细菌表面疏水性的强弱与细胞表面蛋白、菌毛、脂磷壁酸、多糖等有关,其被认为与细菌的特异性黏附相关[20-21]。为了进一步研究乳酸杆菌表面疏水性与黏附性的关系,利用BATH法测定了乳酸杆菌的表面疏水能力。M5-L显示出了很好的表面疏水性,F0421的表面疏水能力仅次于M5-L。通过进一步的研究分析,对两种菌株分别使用氯化锂和高碘酸钠处理,分别破坏其表面的S-层蛋白和多糖,而后再与两种致病菌进行共聚作用,结果表明氯化锂处理能够显著降低F0421菌株的共聚能力,而高碘酸钠处理则显著降低M5-L菌株的共聚能力,说明F0421表面的S-层蛋白是其发挥黏附作用的活位点,而M5-L的粘附活位点则是多糖。而实验中经氯化锂处理后的M5-L的共聚能力相比于未处理的菌株有所上升,推测可能是其表面覆盖的S-层蛋白对多糖的黏附位点有一定的掩盖作用,经氯化锂去除后,将多糖位点暴露,使得M5-L的共聚能力有所上升,我们推测位于F0421表面黏附位点的S-层蛋白是参与乳酸杆菌黏附的主要物质。陈臣等[22]利用荧光标记法来检测植物乳杆菌ST-Ⅲ对Caco-2细胞的黏附机理,利用化学法或酶法去除菌体的S-层蛋白,进行黏附性实验。结果表明,去除S-层蛋白的L.plantarum ST-Ⅲ对Caco-2细胞的黏附能力显著降低,而将除去S-层蛋白的菌体与提取的表面蛋白进行孵育后,再进行黏附性实验,其黏附率从4.82%提高至8.46%,结论也证明了表面蛋白参与黏附过程。
乳酸杆菌的S-层蛋白是其细菌表面的一种蛋白成分,以非共价键与细胞壁结合的包被结构。研究证实,用提取剂去除S-层蛋白后,菌体能够重新组装其结构[23]。在本实验中,经氯化锂处理后的乳酸菌体对致病菌的共聚能力随着时间的延长而增加,也再次证实了其表面的S-层蛋白的再生能力。而经高碘酸钠处理的菌体,其与致病菌的共聚能力也随时间延长而增强,可能也是由于乳酸杆菌表面的多糖发生了再生。
4 结论
本研究表明,五株乳酸杆菌中,自聚能力、疏水性较好的均为M5-L和F0421,同时,M5-L及F0421与K的共聚效果强于其与Ec的共聚作用,并且乳酸杆菌与致病菌的共聚能力与共聚时间成正相关。研究证实,F0421菌株表面的主要活性物质为S-层蛋白,而M5-L菌株表面的主要活性物质则为多糖。实验结果表明不同的乳酸菌菌株具有不同的表面性质,这对于开发具有潜力的益生菌,并应用于功能性食品、发酵等行业具有重要意义。
[1]张英春,韩雪,张兰威,等.益生菌抑制致病菌作用的机制研究进展[J].微生物学通报,2012,39(9):1306-1313.
[2]Kleerebezem M,Bernard E.The extracellular biology of the lactobacilli[J].Fems Microbiology Reviews,2010,34(2):199-230.
[3]汪海锋,章文明,汪以真,等.乳酸杆菌与肠道黏附相关表面因子及其机制研究进展[J].动物营养学报,2011,23(2):179-186.
[4]肖荣,王远亮,李宗军.益生性乳酸菌黏附性研究进展[J].食品与发酵工业,2008,34(5):134-137.
[5]佟盼盼,刘莉,杨桂连,等.乳酸杆菌表面结构和粘附特性的研究进展[J].中国微生态学杂志,2012,24(10):941-947.
[6]李清,刘小莉,王英,等.植物乳杆菌表面性质及对Caco-2细胞的黏附[J].食品科学,2015,36(09):97-101.
[7]García-cayuela T,Korany A M,Bustos I,et al.Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype[J].Food Research International,2014,57:44-50.
[8]Tuo Y F,Yu H L,Ai L Z,et al.Aggregation and adhesion properties of 22 Lactobacillus strains[J].Journal of Dairy Science,2013,96(7):4252-4257.
[9]白洁,李卫芬,黄琴,等.几株益生乳酸菌对Caco-2细胞的黏附及其对致病菌黏附的影响[J].动物营养学报,2012,24(10):1992-1998.
[10]Liu X M,Liu W Y,Zhang Q X,et al.Screening of lactobacilli with antagonistic activity against enteroinvasive Escherichia coli[J].Food Control,2013,30(2):563-568.
[11]Del R B,Sgorbati B,Miglioli M,et al.Adhesion,autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum[J].Letters in Applied Microbiology,2000,31:438-442.
[12]Bernet M F,Brassar T D,Neeser J R,er al.Adhesion of Human Bifidobacterial Strains to Cultured Human Intestinal Epithelial Cells and Inhibition of Enteropathogen-Cell Interactions[J].Applied Environmental Microbiology,1993,59(12):4121-4128.
[13]Collado M C,Meriluoto J,Salminen S.Adhesion and aggregation properties of probiotic and pathogen strains[J].European Food Research and Technology,2008,226:1065-1073.
[14]Lortal S V,Heijenoort J,Gruber K,et al.S-layer of Lactobacillus helveticus ATCC 12046:isolation,chemical characterization and re-formation after extraction with lithium chloride[J].Journal of General Microbiology,1992,139:611-618.
[15]Greene J D,Klaenhammer T R.Factors involved in adherence of lactobacilli to human Caco-2 cells[J].Applied and Environmental Microbiology,1994,60:4487-4494.
[16]Boris S,Suarez J E,Barbes C,et al.Characterization of the aggregation promoting factor from Lactobacillus gasseri,a vaginal isolate[J].Journal of Applied Microbiology,1997,83(4):413-420.
[17]Mastromarino P,Brigidi P,Macchia S,et al.Characterization and selection of vaginal Lactobacillus strains for the preparation of vaginal tablets[J].Journal of Applied Microbiology,2002,93(5):884-893.
[18]Kos B,Suskovic J,Vukovic S,et al.Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92[J].Applied Microbiology and Biotechnology,2003,94:981-987.
[19]Golowczyc M A,Mobili P,Garrote G L,et al.Protective action of Lactobacillus kefir carrying S-layer protein against Salmonella enterica serovar Enteritidis[J].International Journal of Food Microbiology,2007,118:264-273.
[20]Remus D M,Kleerebezem M,Bron P A.How probiotic lactobacilli communicate with the host[J].European Journal of Pharmacology,2011,668:s33-s42.
[21]Ren D Y,LI C,Qin Y Q,et al.Inhibition of Staphylococcus aureus adherence to Caco-2 cells by lactobacilli ang cell surface properties that influence attachment[J].Anaerobe,2012,18:508-515.
[22]陈臣,周方方.荧光标记法初探植物乳杆菌ST-Ⅲ对 Caco-2 细胞的黏附机理[J].微生物学通报,2010,37(3):355-361.
[23]李宗军,杨秀华,刘元元,等.乳杆菌S-层蛋白的多样性及其研究方法[J].食品与生物技术学报,2009,28(6):721-725.
Surface properties and adhesion ability of Lactobacilus
XIANG Xin-ling1,ZHANG Ying-chun1,2,*,MA Fang2,ZHANG Lan-wei1,LI Shao-hui1
(1.Department of Food Science and Engineering in School of Chemical Engineering and Technology,Harbin Institute of Technology,Harbin150090,China; 2.School of Municipal and Environmental Engineering,Harbin Institute of Technology,Harbin 150090,China)
Lactobacillus are the intestinal probiotics and they usually play an important role in intestinal through the ability of adhesion and colonization.In order to realize the relationship between surface properties and adhesion,the autoaggregation ability,copolymerization ability with Escherichia coli ATCC25922 or Bacillus subtilis K and cell surface hydrophobicity of five Lactobacillus strains from different sources were assessed.At the same time,the compounds associated with the adhesion process were investigated by using LiCl-treated and sodium periodate-treated.The results showed that L.paracasei subp.paracasei M5-L and L.johnsonii F0421 showed better surface properties and adhesion ability.When Lactobacillus coaggregated with Bacillus subtilis K had much better copolymerization.Meanwhile,when treated with LiCl or sodium periodate,the autoaggregation and adhesion of Lactobacillus were decreased,indicating that the surface layer protein and polysaccharide play an important role in the process of adhesion.
Lactobacilus;autoaggregation;hydrophobicity;copolymerization;surface property
2015-08-07
向鑫玲(1992-),女,在读硕士,研究方向:乳酸杆菌及其活性产物研究,E-mail:xiangxinling1992@163.com。
张英春(1975-),女,博士,副教授,研究方向:乳酸杆菌及其活性产物研究,E-mail:zyc229@163.com。
国家自然科学基金项目(31301515);中国博士后基金特别资助项目(2013T60382);中国博士后基金项目(2012M510093);黑龙江省自然科学基金项目(C201433)。
TS201.3
A
1002-0306(2016)07-0126-06
10.13386/j.issn1002-0306.2016.07.017
猜你喜欢
当代水产(2022年1期)2022-04-26
新疆有色金属(2021年2期)2021-05-22
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年3期)2016-11-15
郑州大学学报(医学版)(2015年2期)2015-02-27
心脑血管病防治(2014年3期)2014-08-18
西北农林科技大学学报(自然科学版)(2014年10期)2014-03-26
食品工业科技(2014年9期)2014-03-11
大理大学学报(2014年4期)2014-02-13
中国神经精神疾病杂志(2012年8期)2012-04-19
