
成都市区白颊噪鹛繁殖行为及对城市环境的行为适应
2016-12-09 10:06严勇柴璐艳吴永杰窦亮冉江洪
四川动物 2016年6期
严勇, 柴璐艳, 吴永杰, 窦亮, 冉江洪
(四川大学生命科学学院,生物资源与生态环境教育部重点实验室,成都 610064)
成都市区白颊噪鹛繁殖行为及对城市环境的行为适应
严勇, 柴璐艳, 吴永杰, 窦亮, 冉江洪*
(四川大学生命科学学院,生物资源与生态环境教育部重点实验室,成都 610064)
2012—2013年繁殖季节,在成都市四川大学望江校区观察了白颊噪鹛Garrulaxsannio的繁殖行为,与已有的研究结果相比,结果显示:(1)白颊噪鹛巢材选择广泛,可就地利用多种材料,包括各种生活垃圾;(2)白颊噪鹛在市区主要营巢于高大乔木树种上,在市郊营巢于竹林或较低矮乔木树种;(3)影响其巢址选择的主要因素为胸径、巢位高度、树高;(4)孵卵时间为11~12 d,较1950年代(15~17 d)明显缩短;(5)育雏时间为12 d,但市区白颊噪鹛在卵质量、育雏期日喂食频次、出巢日雏鸟体质量和体长上均高于市郊;(6)市区白颊噪鹛繁殖成功率(73.3%)高于市郊(23.1%)。在城市化进程中,市区白颊噪鹛较市郊个体产生了行为的适应性变化,这些行为变化是其成为城市优势种的重要原因。
白颊噪鹛;繁殖行为;适应性变化;城市化;巢址选择;孵卵节律
随着当前城市化进程的加剧,有关城市鸟类行为适应性变化以及城市环境对鸟类的影响成为国内外研究的热点(Beissinger & Osborne,2010;Alberti,2015;叶淑英等,2015)。城市环境的改变使当地原有的一些鸟类减少或是区域灭绝,也出现了由于城市环境相似性导致的“鸟类的城市趋同”现象(Clergeauetal.,2006;Devictoretal.,2007)。一些鸟类,尤其是城市常见鸟类,如树麻雀Passermontanus、紫翅椋鸟Sturnusvulgaris、白头鹎Pycnonotussinensis等,其种群数量随着城市化的提升而增加,并逐渐成为优势种(Emlen,1974;于同雷等,2011)。与自然环境相比,城市环境具有噪音大、干扰多、适宜生境零散、食物资源相对集中等特点。城市鸟类通过调整原有生活史策略以适应复杂的城市环境,这些“城市适应者”在城市环境中产生了行为或生理适应性变化,如大山雀Parusmajor改变鸣声的频率以适应城市噪音(Slabbekoorn,2013);鸟类惊飞距离的长短是对不同人为干扰强度的响应(Blumstein,2003),城市中的红腹灰雀Pyrrhulapyrrhula较农村中的同类提升了解决问题和学习的技巧,变得更大胆,生理免疫力增强等(Audetetal.,2016)。
白颊噪鹛Garrulaxsannio是我国南方地区常见的鸟类,也是成都市常见的鸟类优势种(吴先智等,2005)。目前,对白颊噪鹛的研究涉及鸣声、取食生态位、繁殖行为、非繁殖季节行为节律等方面(李桂垣,1982;李明等,2009;朱峰等,2010;晏林波等,2012;曹长雷等,2013;柴璐艳等,2014),其中关于其繁殖行为的研究见于李桂垣(1982)1950年代在四川省成都市、朱峰等(2010)在四川省南充市郊对白颊噪鹛的繁殖行为研究报道,晏林波等(2012)对四川省南充市郊白腰文鸟Lonchurastriata、白颊噪鹛和白头鹎巢大小、结构及巢材的比较研究报道。2012年3—8月、2013年4—7月,本研究在成都市四川大学望江校区内对白颊噪鹛的繁殖行为进行了较系统的观察,以了解城市中白颊噪鹛的繁殖特点,丰富其生物学资料。通过与已有市郊研究结果的比较,了解白颊噪鹛在繁殖期适应市区环境而产生的行为适应性变化,为研究鸟类对城市环境的适应提供参考案例。这有助于了解城市鸟类在城市化进程中对城市环境变化的响应,进一步探究城市鸟类进化适应以及与环境变化的关系。
1 研究地区与研究方法
1.1 研究区域概况
四川大学望江校区位于成都市城区中心一环路九眼桥旁,占地面积1.73 km2,建筑面积1.3 km2。地理坐标为104°4′59.5″~104°5′54.3″E,30°37′63.2″~30°37′66.7″N,海拔480 m。属亚热带湿润季风气候,年平均降水量945.6 mm,年平均气温16.2 ℃。校园植被主要为人工绿化植被,植物种类多达461种,常见乔木树种有鸡爪槭Acerpalmatum、香樟Cinnamonumcamphora、小叶榕Ficusconcinna和银杏Ginkgobiloba等;常见灌木种类有栀子Gardeniajasminoides、小叶女贞Ligustrumquihoui、凤尾竹Bambusamultiplex等;常见草本植物有早熟禾Poaannua、吊兰Chlorophytumcomosum等(杨建欣,白洁,2010)。四川大学望江校区是成都市区著名的观鸟地点之一,常见的鸟类有白颊噪鹛、树麻雀、珠颈斑鸠Streptopeliachinensis、红头长尾山雀Aegithalosconcinnus、乌鸫Turdusmerula、白头鹎等(李偲等,2010)。
1.2 研究方法
2012年3—8月和2013年4—7月,持续对出现衔枝筑巢、警报鸣叫、衔虫跳窜飞行等行为的白颊噪鹛进行观察和追踪,在其活动频繁的区域寻找巢址。发现巢后,用直尺(精度为0.1 cm)测量并记录巢大小、巢材等参数和位置并编号。
待幼鸟出飞或亲鸟离巢后,对筑于乔木的巢测量其巢址的7个相关参数:胸径、树高、巢枝直径、巢枝夹角、巢位高度、隐蔽度、人为活动强度。其中隐蔽度分8个等级:在巢周围8个方向观察鸟巢,8个方向均无法见到鸟巢,定义为8级,7个方向均无法见到,定义为7级,以此类推分成1~8级;人为活动强度分5个等级:距巢树5 m范围内平均每分钟通过的人数>40人,定义为5级,30~39人,定义为4级,20~29人,定义为3级,10~19人,定义为2级,10人以下,定义为1级;分别在08∶00—09∶00、11∶00—12∶00、14∶00—15∶00、17∶00—18∶00测定距巢树5 m内的人流量,并在1个月内随机选择日期重复4次,不同位置的测定时间相同。
选择易于观察的巢对白颊噪鹛的繁殖行为进行持续观察。亲鸟产卵后,戴薄膜手套取出鸟卵,用电子秤和游标卡尺测量记录卵质量、大小等参数,并用水性笔对每枚卵进行标记编号。每天06∶30—19∶30用10×42倍望远镜连续观察记录孵卵期间亲鸟离巢次数、离巢时间等参数。
育雏期间,在亲鸟离巢觅食后,戴薄膜手套取出幼鸟,用彩色塑料壳套在跗趾上标记,每天11∶00—13∶00在亲鸟离巢时使用电子秤和游标卡尺测量巢中幼鸟的体质量、体长等生长参数,测完数据把幼鸟按原来位置放回巢中。每天06∶00—20∶00,利用8×42倍望远镜以及结合拍照、录像等手段在巢周围选取的隐蔽点连续观察亲鸟喂食幼鸟情况、记录离巢次数和时间、喂食频次等参数。
用主成分分析法对巢址选择的7个变量(胸径、树高、巢枝直径、巢枝夹角、巢位高度、隐蔽度、人为活动强度)进行主成分分析,确定白颊噪鹛对巢址选择的主要因素,相关统计分析在SPSS 17.0中完成。所有数据均用平均值±标准差(Mean±SD)表示。
2 结果与分析
2.1 巢及巢址
共发现30个白颊噪鹛的巢,其中27个位于树上,3个位于灌丛或竹丛中。巢分别位于小叶榕、黄葛树Ficusvirensvar.sublanceolata、香樟、棕竹Rhapisexcelsa和凤尾竹5种植物中,所占比例分别为70.00%、13.30%、6.67%、6.67%、3.33%。
对其中26个巢大小测量的结果为:外径13.59(±3.20) cm×11.71(±1.15) cm;内径8.69(±0.69) cm×7.49(±0.76) cm;巢高10.23(±1.51) cm;巢深5.14(±0.60) cm。
巢呈碗形,分内外2层:外层多为棕榈、小叶榕、鸡爪槭的叶片,小叶榕的气须根,以及草茎、细藤等;内层为小叶榕气须根、草茎等,也有一些碎纸、碎塑料袋(条)、破棉花等生活垃圾。巢外部颜色灰暗与周围环境融为一体,内部基本上为黑色,内外颜色对比鲜明。
2.2 卵及孵卵行为
白颊噪鹛每巢产卵2~4枚(n=5)。卵的平均大小为24.28(±1.34) mm×18.96(±1.76) mm,卵质量为5.40(±0.13) g(n=12)。孵卵期为11~12 d(n=4),雌、雄轮换孵卵。孵卵期,随孵化时间的延长,亲鸟离巢时间和离巢次数逐渐减少(表1)。
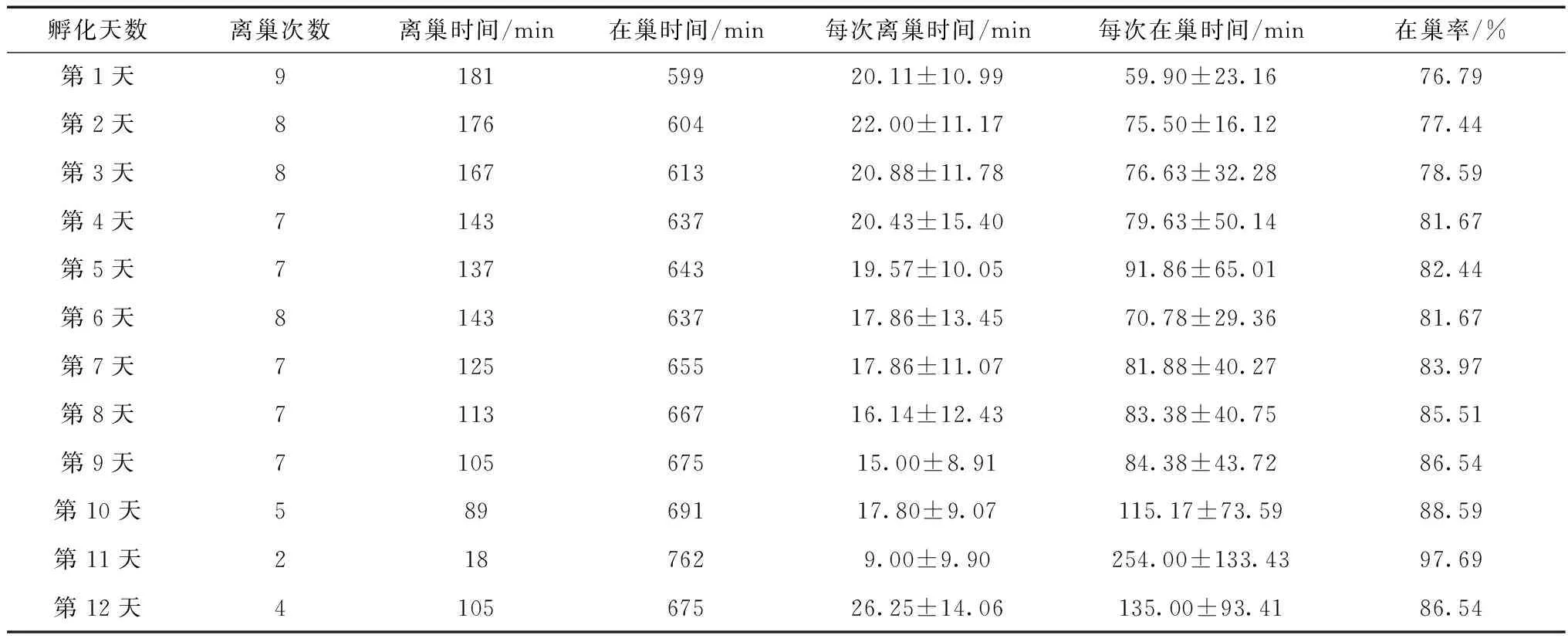
表1 白颊噪鹛孵卵节律
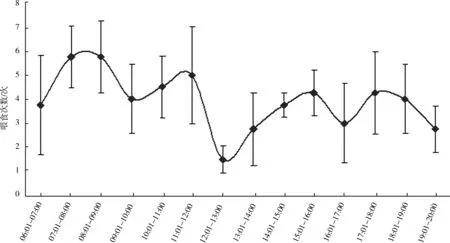
图1 白颊噪鹛亲鸟日喂食频次(平均值±标准差)
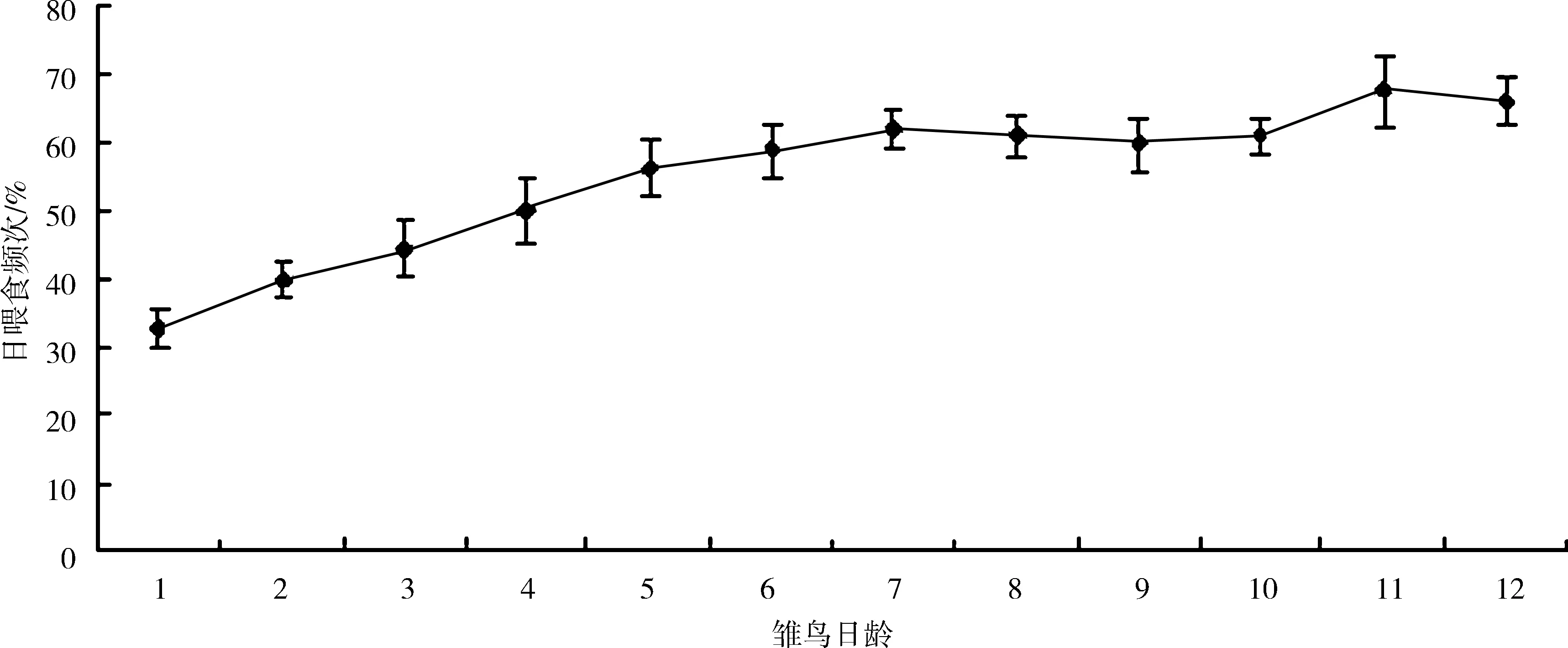
图2 白颊噪鹛亲鸟日喂食频次节律(平均值±标准差)
2.3 育雏行为
4巢育雏期亲鸟喂食节律的数据统计表明:亲鸟在一天之内有4个喂食高峰(图1),分别是07∶01—09∶00、11∶01—12∶00、15∶01—16∶00、17∶01—18∶00,相邻2次高峰出现的时间间隔为2~3 h。高峰期亲鸟的喂食频次可达每小时5.80次±1.28次/h。从日节律来看,上午的喂食频次高于下午和黄昏时分,12∶01—13∶00的喂食频次最低。从雏鸟日龄与喂食频次的关系来看(图2),1~7日龄亲鸟日喂食频次逐渐增加,由每日32.75次±2.87次增加到每日67.50次±5.26次,基本维持在每日喂食60次左右。亲鸟在育雏期间喂食给雏鸟的全为动物性食物。
雌雄亲鸟共同育雏,育雏期(雏鸟出壳到雏鸟离巢)为12 d(n=4),共监测5巢15个卵,出巢日共记录11只白颊噪鹛,繁殖成功率为73.3%。
2.4 巢树选择
对7个巢址选择数值型变量的主成分分析结果显示:前3个主成分的特征值均大于1,累积贡献率达70%(表1),提取前3个主成分并计算各变量特征向量(表2)。
由表2和表3可知:胸径、巢位高度、树高在第一主成分中贡献较高,这3个参数反映出白颊噪鹛对巢安全性的需求;巢枝夹角在第二主成分中相关系数较高;第三主成分中,巢枝直径相关系数较高。
2.5 城市与城郊繁殖行为的对比
本研究与李桂垣(1982)在四川成都市市郊(1956年的观察)、朱峰等(2010)在四川南充市城郊(2009年观察)、晏林波等(2012)在四川南充市郊(2011年观察)的研究相比较,发现白颊噪鹛的巢型和巢大小在4个研究中几乎无变化。但在1950年代的巢材上基本没有发现生活垃圾,而近几年的巢材中都出现了生活垃圾,而且巢材与城市绿化植物(营巢树、竹)的变化有关。
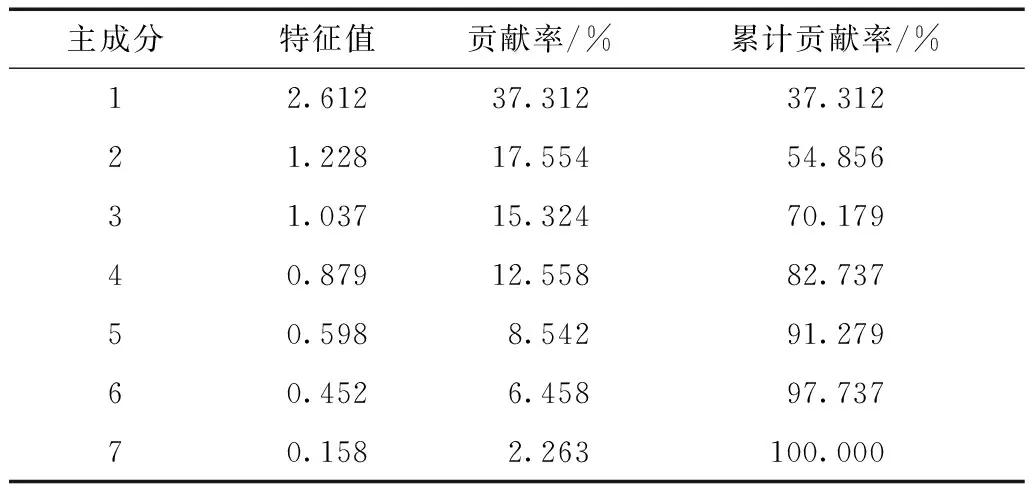
表2 白颊噪鹛巢址选择主成分分析(n=27)
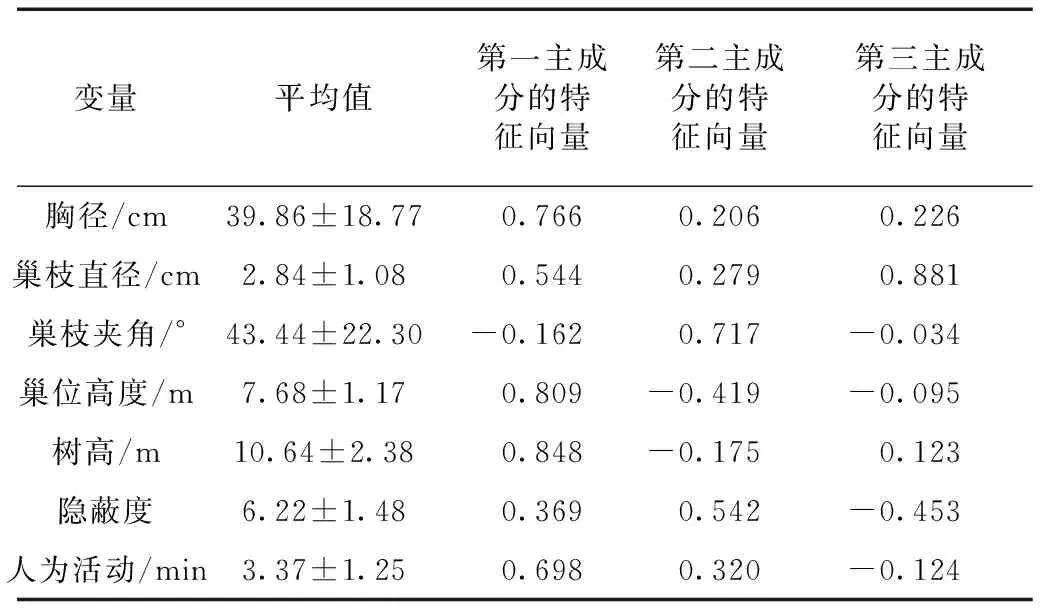
表3 白颊噪鹛巢址选择参数特征向量的转置矩阵
不同时期城区和郊区的白颊噪鹛在营巢植物的选择上出现了较大的差异(表4):朱峰等(2010)的研究表明白颊噪鹛在南充市郊主要营巢于竹林中,巢位较低;晏林波等(2012)的研究表明在市郊以天竺桂Cinnamomumjaponicum为优势种的生境中,白颊噪鹛巢全部位于其中,巢位较低;在城区白颊噪鹛主要营巢于常见绿化树种上,巢位较高。在巢高度上,城区的巢平均高度明显高于郊区的巢,本研究与市郊(朱峰等,2010)白颊噪鹛巢址选择主要因素的第一主成分基本相同:分别为巢位高、巢树(竹)高度、巢树胸径(竹直径)。
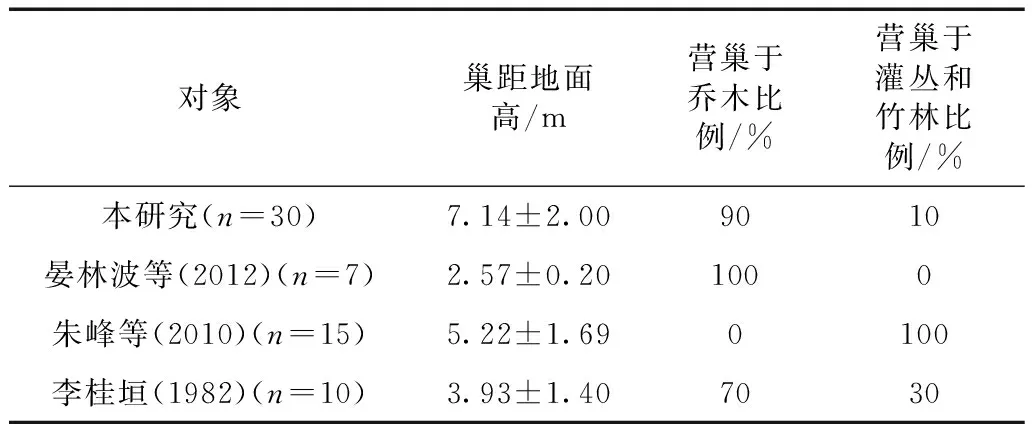
表4 不同研究中白颊噪鹛筑巢参数
3个研究中白颊噪鹛的产卵数、卵色、卵形无变化;本研究白颊噪鹛卵质量稍大于朱峰等(2010)在市郊的观察结果;在孵化时间上,本研究和朱峰等(2010)比李桂垣(1982)的观察结果明显缩短(表5)。

表5 不同时期和城区白颊噪鹛孵卵的差异
3个研究中白颊噪鹛的育雏时间均为12 d,但市区白颊噪鹛亲鸟的喂食频次、离巢日雏鸟的体质量、体长和繁殖成功率都高于市郊(成都市区为73.3%,南充市郊23.1%;表6)。

表6 白颊噪鹛雏鸟发育差异
3 讨论
城市化程度的提高影响了鸟类对巢材的选择,城市鸟类在筑巢中增加了对人类生活材料的利用(Wangetal.,2009)。与李桂垣(1982)1950年代对成都市白颊噪鹛巢材的观察结果相比,当前白颊噪鹛巢材发生了变化,不仅出现了人类生活垃圾,如碎纸、塑料袋等,而且巢材与城市绿化植物(营巢树、竹)的变化有关:1950年代白颊噪鹛多筑巢于柏树和竹林,巢材基本为松柏枝、草茎和竹叶;晏林波等(2012)研究表明白颊噪鹛全筑巢于天竺桂,巢材多为天竺桂叶、枇杷叶、草茎等。在城市化进程中,白颊噪鹛具有利用多种环境材料的能力,白颊噪鹛巢材的变化反映了其巢材的选择具有一定的灵活性,也从侧面反映出环境中存在大量而广泛的人类生活垃圾。
成都市区白颊噪鹛营巢树种中乔木比例达到90%,朱峰等(2010)、晏林波等(2012)在南充市郊的研究表明白颊噪鹛全部筑巢于乔木树种中,这说明白颊噪鹛能在城市中适应以乔木为主的绿化植物,其巢址选择具有一定的灵活性和适应性。在巢址选择上,本研究和朱峰等(2010)研究的第一主成分基本相同,分别为巢高、树(竹)高和大小,这反映了市区和城郊白颊噪鹛对巢址的安全性要求相同。本研究白颊噪鹛巢距地面的平均高度比朱峰等(2010)、晏林波等(2012)的5.22 m、2.57 m更高,城市白颊噪鹛提高巢位高度是对城区以高大乔木为主的绿化植被以及高频次人为活动干扰的适应,在城市高大乔木筑巢和在较低矮且隐蔽度高的竹林筑巢都能提高巢址的安全性。研究表明合适的巢址可以帮助鸟类将各种风险降到最低,而巢位高度和巢的隐蔽性则可有效抵御天敌和人为干扰的侵袭(Ricklefs,1969;Greggetal.,1994;Stahlschmidtetal.,2011)。市区白颊噪鹛在面对城市更高的人为干扰强度时调整筑巢策略,体现了城市鸟类筑巢策略对环境变化的适应。
本研究和朱峰等(2010)中白颊噪鹛的孵卵时间较之李桂垣(1982)研究结果缩短3~5 d,这可能是气候变化的结果(Yuta & Koizumi,2011)。在卵体积和质量近似的情况下,环境背景温度的升高有助于加速卵中胚胎的发育,因此造成卵孵化期的缩短(杨振才等,2002)。
本研究中育雏期间白颊噪鹛亲鸟喂食频次、离巢日雏鸟的体质量和体长均略高于南充市郊白颊噪鹛的,这说明市区白颊噪鹛面对食物资源更丰富的环境,通过提高喂食幼鸟的频次,使幼鸟获得充足的营养和良好的发育条件从而获得更高的繁殖成功率(Gonzalezetal.,2006)。城区白颊噪鹛的繁殖成功率高于郊区,这可能与市区内食物集中丰富、捕食风险较小有关。丰富的食物是促进雏鸟生长发育的重要条件,较小的捕食风险使白颊噪鹛获得较高的繁殖成功率,使其城区种群快速扩展进而成为城市环境的优势种。
曹长雷, 戴玄, 韩宗先, 等. 2013. 长江师范学院校园白颊噪鹛取食生态位的初步研究[J]. 湖北农业科学, 52(2): 402-404.
柴璐艳, 赵璐玲, 纪维雯, 等. 2014. 城市白颊噪鹛群体非繁殖季节的行为节律及时间分配[J]. 四川动物, 33(1): 66-70.
李桂垣. 1982. 白颊噪鹛繁殖习性的初步观察[J]. 动物学研究, 3(3): 245-250.
李明, 郭延蜀, 韩艳良, 等. 2009. 白颊噪鹛四川亚种繁殖期鸣声声谱分析[J]. 四川动物, 28(6): 913-916.
李偲, 张敦房, 黄科. 2010. 四川大学江安校区鸟类调查[J]. 四川动物, 29(2): 249-254.
吴先智, 杨靖, 朱章顺, 等. 2005. 成都市区公共绿地野生鸟类调查初报[J]. 四川动物, 24(4): 568-574.
晏林波, 周材权, 袁施彬, 等. 2012. 南充市同域分布的白腰文鸟白颊噪鹛和白头鹎巢和巢材比较研究[J]. 内江师范学院学报, 27(12): 43-47.
杨建欣, 白洁. 2010. 四川大学望江校区校园植物配置[J]. 中国城市林业, 8(2): 25-27.
杨振才, 牛翠娟, 孙儒泳. 2002. 温度对中华鳖卵孵化和胚胎发育的影响[J]. 动物学报, 48(6): 716-724.
叶淑英, 郭书林, 路纪琪. 2015. 中国城市鸟类生态学研究进展与展望[J]. 河南教育学院学报(自然科学版), 24(3): 47-53.
于同雷, 符建荣, 郭延蜀. 2011. 四川南充地区白头鹎种群数量增长原因分析[J]. 信阳师范学院学报(自然科学版), 24(4): 495-498.
朱峰, 周材权, 杨志松, 等. 2010. 四川南充白颊噪鹛的繁殖行为观察[J]. 动物学杂志, 45(4): 150-155.
Alberti M. 2015. Ecoevolutionary dynamics in an urbanizing planet[J]. Trends in Ecology & Evolution, 30(2): 114-126.
Audet JN, Ducatez S, Lefebvre L. 2016. The town bird and the country bird: problem solving and immunocompetence vary with urbanization[J]. Behavioral Ecology, 27(2): 637-644.
Beissinger SR, Osborne DR. 2010. Effects of urbanization on avian community organization[J]. The Condor, 84(1): 75-83.
Blumstein DT. 2003. Flight-initiation distance in birds is dependent on intruder starting distance[J]. Journal of Wildlife Management, 67(4): 852-857.
Clergeau P, Croci S, Jokimaki J,etal. 2006. Avifauna homogenisation by urbanization: analysis at different European latitudes[J]. Biological Conservation, 127(3): 336-344.
Devictor V, Julliard R, Couvet D,etal. 2007. Functional homogenization effect of urbanization on bird communities[J]. Conservation Biology, 21(3): 741-751.
Emlen JT. 1974. An urban bird community in Tucson, Arizona: derivation, structure, regulation[J]. The Condor, 76(2): 184-197.
Gonzalez LM, Margalida A, Sánchez R,etal. 2006. Supplementary feeding as an effective tool for improving breeding success in the Spanish imperial eagle (Aquilaadalberti)[J]. Biological Conservation, 129(4): 477-486.
Gregg MA, Crawford JA, Drut MS,etal. 1994. Vegetational cover and predation of sage grouse nests in Oregon[J]. The Journal of Wildlife Management, 58(1): 162-166.
Ricklefs RE. 1969. An analysis of nesting mortality in birds[M]. Washington, D.C.: Smithsonian Institution Press: 1-48.
Slabbekoorn H. 2013. Songs of the city: noise-dependent spectral plasticity in the acoustic phenotype of urban birds[J]. Animal behavior, 85: 1089-1099.
Stahlschmidt ZR, Brashears J, DeNardo DF. 2011. The role of temperature and humidity in python nest site selection[J]. Animal Behaviour, 81(5): 1077-1081.
Wang YP, Chen SH, Blair RB,etal. 2009. Nest composition adjustments by Chinese bulbulsPycnonotussinensisin an urbanized landscape of Hangzhou[J]. Acta Ornithologica, 44(2): 185-192.
Yuta T, Koizumi I. 2011. Long breeding season and high frequency of multiple brooding in great tits in northern Japan[J]. Ardea-Wageningeningen, 100(2): 197-201.
Breeding Behavior ofGarrulaxsannioin Chengdu and Its Adaptation to the Urban Environments
YAN Yong, CHAI Luyan, WU Yongjie, DOU Liang, RAN Jianghong*
(Key Laboratory of Eco-resource and Eco-environment of Ministry of Education, College of Life Sciences,Sichuan University, Chengdu 610064, China)
The breeding behavior of the white-browed laughingthrush (Garrulaxsannio) was systematically studied from 2012 to 2013 in the Wangjiang campus of Sichuan University, which is located in Chengdu urban area. Compared with the previous studies ofG.sannioin different periods and sites, we found that: (1)G.sanniowould use diversified nest materials, including the human garbage; (2) They mostly nested in high trees of urban area while nesting in bamboos or low trees of suburban area; (3) The main factors affecting the nest-site selection were tree breast-height diameter, nest height, and tree height; (4) The contemporary incubation time ofG.sanniowas shortened to 11~12 d compared to 15~17 d in 1950s; (5) The mean feeding phase ofG.sanniowas 12 d, but the egg weight, daily feeding frequency, weight and length of nestling on leaving-nest day of the birds in urban area were higher than those in suburban area; (6)The breeding success rate was 73.3% in urban area compared with 23.1% in suburban area. Compared with those living in suburban area,G.sannioof urban area evolved adaptive changes of breeding behavior that helped them become the dominant urban species in the process of urbanization.
Garrulaxsannio; breeding behavior; adaptive changes; urbanization; nest-site selection; incubation rhythm
2016-08-16 接受日期:2016-10-19
严勇(1992—), 男, 硕士研究生, 研究方向为保护生物学, E-mail:396566617@qq.com
*通信作者Corresponding author, E-mail:rjhong-01@163.com
10.11984/j.issn.1000-7083.201600221
Q959.7; Q958.1
A
1000-7083(2016)06-0936-06
猜你喜欢
学与玩(2022年9期)2022-10-31
野生动物学报(2021年4期)2021-11-14
文苑(2020年12期)2020-04-13
广东第二课堂·小学(2019年5期)2019-06-14
小太阳画报(2019年1期)2019-06-11
森林与人类(2018年6期)2018-12-03
幼儿画刊(2018年10期)2018-10-27
中国畜禽种业(2018年12期)2018-01-18
小学生必读(低年级版)(2017年5期)2017-08-12
中外玩具制造(2016年5期)2016-11-06
