
燕麦资源生物学性状多样性分析
2017-01-06 02:21穆志新刘龙龙张丽君马名川周建萍崔林
山西农业科学 2016年12期
穆志新,刘龙龙,张丽君,马名川,周建萍,崔林
(山西省农业科学院农作物品种资源研究所,农业部黄土高原作物基因资源与种质创制重点实验室,山西太原030031)
燕麦资源生物学性状多样性分析
穆志新,刘龙龙,张丽君,马名川,周建萍,崔林
(山西省农业科学院农作物品种资源研究所,农业部黄土高原作物基因资源与种质创制重点实验室,山西太原030031)
为了扩充山西省燕麦育种材料遗传基础,收集289份燕麦种质资源,并对22个生物学性状进行分析评价。结果表明,供试燕麦种质间各性状遗传差异较大,具有较丰富的遗传多样性。根据种质间各性状的遗传差异,对供试材料进行聚类分析,选取68份具有代表性的燕麦材料在距离系数为3.8时划分为6类,其中,种质群Ⅰ包括9份材料,种质群Ⅱ包括12份材料,这2类可作为燕麦矮化育种亲本;种质群Ⅲ包括22份材料,可作为培育饲草品种的亲本;种质群Ⅳ包括5份材料,可作为高产品种的育种亲本;种质群Ⅴ包括8份材料,无突出优异性状;种质群Ⅵ包括12份材料,可作为以株高和穗长为育种目标的燕麦亲本。
燕麦;资源;生物学性状;聚类分析
燕麦是禾本科燕麦属1年生草木植物,是一种重要的粮饲兼用作物,分为皮燕麦和裸燕麦2种[1],主要产区集中在北半球的温带地区。我国栽培的燕麦以裸燕麦为主[2],主要种植在内蒙古、河北、山西、甘肃,种植面积约占全国的90%。山西省燕麦种植主要集中在太行山和吕梁山区[3]。
燕麦有食疗兼备的功能,具有抗旱、耐瘠薄、耐适度盐碱的优点[4]。近年来,由于燕麦新品种的不断更新以及栽培措施的提高,燕麦的产量和品质得到了明显的提升。但是,随着育种进程的加快,育种基础材料的遗传背景越来越窄,选择具有特异的生物学性状的亲本材料是研究工作的重点和难点[5-6]。多数生物学性状除了遗传因素以外,受到外界环境因素的影响比较大,增加了选择的难度。因此,对燕麦生物学性状遗传关系的研究和探讨是十分有必要的。
本研究针对山西省燕麦育种基础材料日渐狭隘的现状,从国内外收集燕麦种质资源共289份,在山西省阳高县进行生物学性状的种植调查,利用遗传变异和聚类分析的方法研究燕麦种质资源的生物学性状之间的遗传关系[7-11],提高这些燕麦种质资源利用中性状选择的效率。
1 材料和方法
1.1 材料
供试材料为289份燕麦种质,均为裸燕麦;聚类分析所用的68份燕麦资源如表1所示。
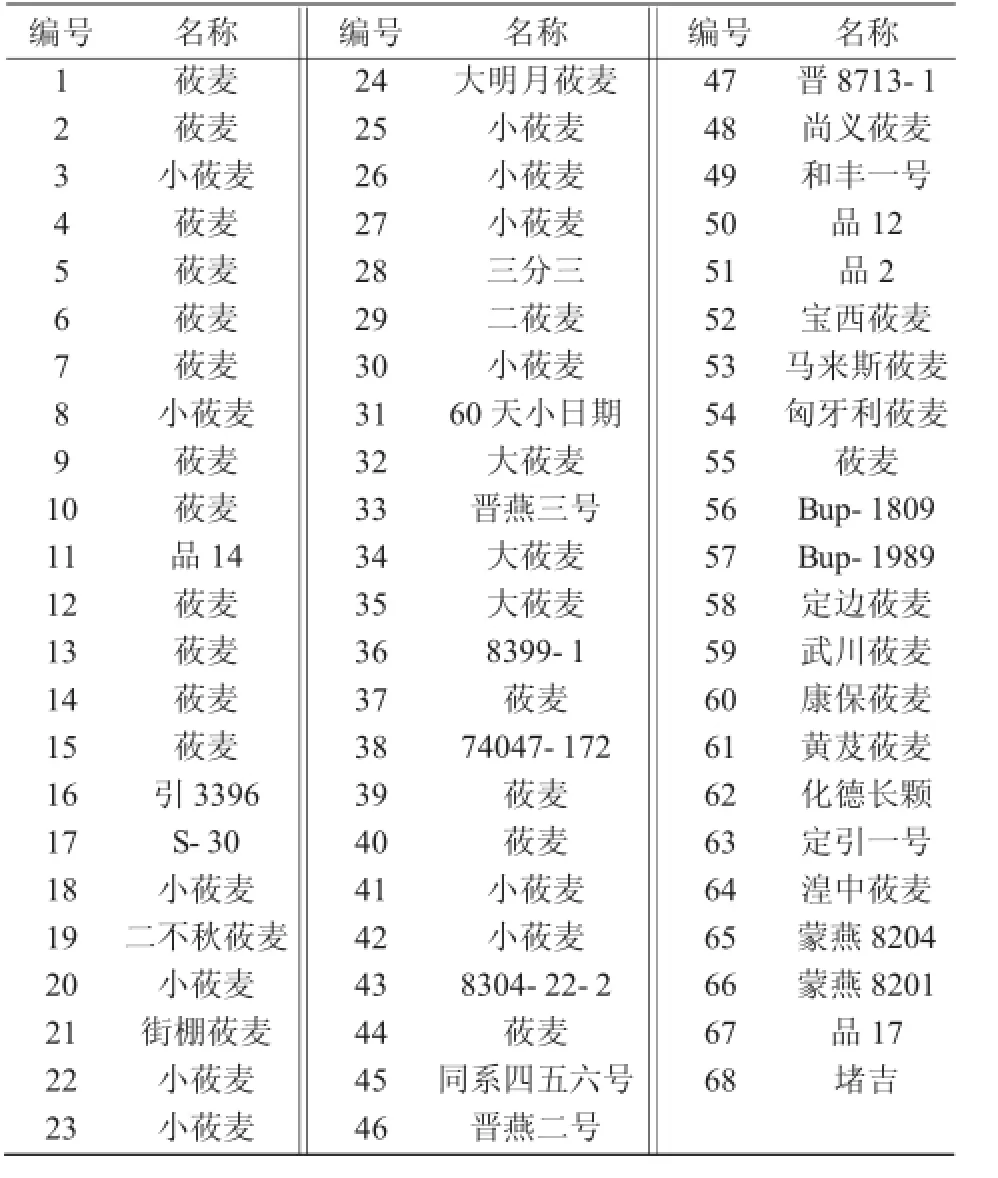
表1 燕麦材料编号、名称
1.2 试验地概况
试验于2015年在山西省大同市阳高县良种研发中心基地进行,该地区位于山西省北部,地理坐标为N40°22′,E113°46′,海拔1 050.3 m。四季分明,日夜温差大。春季多风少雨,温度回升快;夏季炎热多雨。年平均气温7.0℃,年平均降水量333 mm,无霜期约125d。4月17日播种,每小区8行,行长2m,行距25 cm,常规田间管理,燕麦生长期间浇水3次。
1.3 性状调查
田间调查穗色、芒性、芒型、芒色、小穗型、茎节数和茎秆颜色;收获时每小区随机取样10株进行室内考种,调查株高、穗长、穗下茎长、穗下茎长/株高、穗叶距、主穗小穗数、主穗粒数、单株粒质量、穗轮层数、粒型、籽粒颜色、籽粒茸毛、籽粒饱满度、内稃色和外稃色。
性状的统计分析分为2个部分:第1部分是质量性状的分析,分析各类别性状的频率分布和多样性指数,包括穗色、芒型、芒性、芒色、小穗型、内稃色、外稃色、茎秆颜色、粒型、籽粒颜色、籽粒茸毛和籽粒饱满度12个性状;第2部分是数量性状的分析,计算最大值、最小值、平均值、标准差、变异系数、极差和多样性指数,包括株高、穗长、穗下茎长、穗下茎长/株高、穗叶距、主穗小穗数、主穗粒数、单株粒质量、穗轮层数和茎节数10个性状。
1.4 数据处理
数据采用Excel和SPSS 16.0软件进行统计分析。多样性指数的计算采用Shannon-Weaver信息指数,计算公式如下。

其中,Pi为某一性状第i个级别出现的概率[12]。为了便于数量化和统计分析,对数量性状进行分级,质量性状予以赋值。在聚类分析中,种质间遗传距离采用欧氏距离平方法,用离差平方和法进行聚类分析,从而获得聚类树。
2 结果与分析
2.1 燕麦资源生物学性状多样性
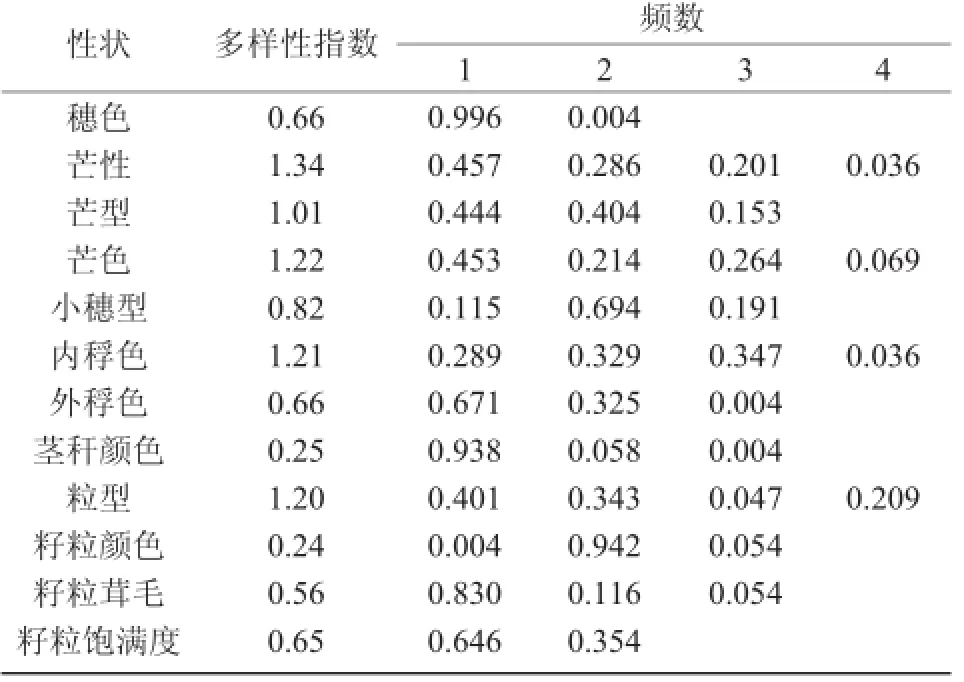
表2 燕麦资源质量性状的遗传多样性
对参试燕麦资源的质量性状分析表明,穗色以白色为主,频数为0.996;芒性以无芒为主,频数为0.457;芒型除无芒外,以直芒为主,频数为0.404;芒色除无芒外,以黄色和白色为主,频数分别为0.214和0.264;小穗型以串铃型为主,频数为0.694;内稃色以黄色和褐色为主,频数分别为0.329和0.347;外稃色以白色为主,频数为0.671;茎秆颜色以黄色为主,频数为0.938;粒型以纺锤型和长筒型为主,频数分别为0.401和0.343;籽粒颜色以白色为主,频数为0.942;籽粒茸毛以少毛为主,频数为0.830;籽粒饱满度以中度饱满为主,频数为0.646。在这些性状中,芒性、芒色、内稃色、粒型和芒型的多样性指数较高,分别为1.34,1.22,1.21,1.20和1.01,说明这5种性状的遗传差异较大(表2)。
由表3可知,主穗粒数的多样性指数为1.29,在10个数量性状中最高;其次是单株粒质量、主穗小穗数和穗下茎长/株高;茎节数最低,为0.23。10个数量性状的多样性指数平均值为0.84。不同性状的变异系数存在很大差异。单株粒质量的变异系数最大,为69.78%,变异范围为0.23~13.52 g,表明参试材料间在该性状上存在丰富的变异,改良的潜力大;其次是主穗小穗数和主穗粒数,变异系数分别为35.23%和35.39%,变异范围为5.50~43.50,9.50~103.50个;茎节数的变异系数最小,为6.34%,说明此性状在各材料间差异较小,较稳定。参试燕麦资源数量性状的统计分析结果说明,各生物学性状差异较大,参试材料具有丰富的多态性,可为燕麦遗传改良提供丰富的材料。

表3 燕麦资源数量性状的多样性
2.2 聚类分析
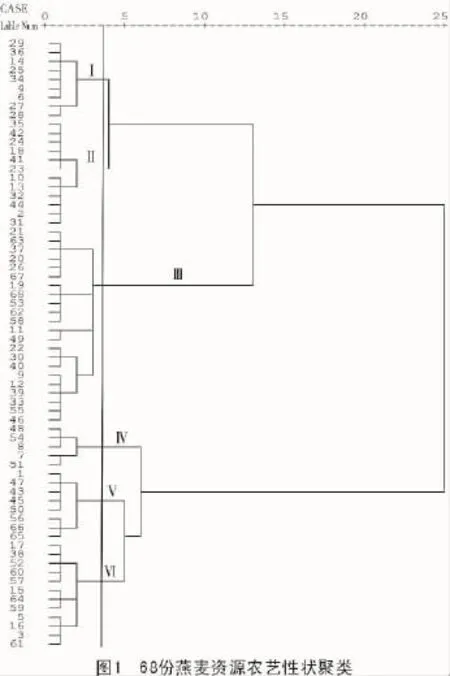
对供试的289份燕麦种质资源的22个性状数据进行初步聚类后,在所得的5类种质群中按照比例随机选取68个燕麦资源进行下一步聚类。在欧式距离系数为3.8时,68份材料被归为6个种质群(图1)。各类群特征列于表4。
种质群Ⅰ有包括五寨县二莜麦等在内的9份材料。这类种质群株高较低,穗长较短,穗下茎长较短,穗下茎长与株高比中等,穗叶距中等,主穗小穗数较少,主穗粒数较少,单株粒质量最小,穗轮层数较少,茎节数较少,芒性为无芒、多芒和少芒,芒型为直芒,小穗型为串铃型和纺锤型,茎秆颜色为黄色,粒型为纺锤型、长筒型和椭圆型,籽粒颜色为白色,籽粒茸毛为少毛和中毛。
种质群Ⅱ有包括神池县莜麦等在内的12份材料。这类种质群株高较低,穗长较短,穗下茎长较短,穗下茎长与株高比较高,穗叶距中等,主穗小穗数较少,主穗粒数较少,单株粒质量较小,穗轮层数较少,茎节数较少,内稃色为白色和黄色,茎秆颜色为黄色,籽粒颜色为白色,籽粒茸毛为少毛和中毛。
种质群Ⅲ有包括昔阳县小莜麦等在内的22份材料。这类种质群株高较高,穗长较长,穗下茎长较长,穗下茎长与株高比较高,穗叶距较大,主穗小穗数中等,主穗粒数中等,单株粒质量较小,穗轮层数中等,茎节数较少,茎秆颜色为黄色,籽粒颜色为白色,籽粒茸毛为少毛和中毛。
种质群Ⅳ有包括锡盟农科所尚义莜麦等在内的5份材料。这类种质群株高较高,穗长较长,穗下茎长较短,穗下茎长与株高比较低,穗叶距较小,主穗小穗数较多,主穗粒数最多,单株粒质量较大,穗轮层数较多,芒性为无芒、少芒,芒型为直芒,小穗型为鞭炮型和串铃型,内稃色为白色和黄色,粒型为纺锤型和长筒型,籽粒茸毛为少毛和中毛。
种质群Ⅴ有包括五寨县莜麦等在内的8份材料。这类种质群株高较低,穗长较短,穗下茎长较短,穗下茎长与株高比中等,穗叶距较小,主穗小穗数中等,主穗粒数较多,单株粒质量中等,穗轮层数较多,茎节数较多,小穗型为鞭炮型和串铃型,茎秆颜色为黄色和白色,粒型为纺锤型、长筒型和椭圆型。
种质群Ⅵ有包括S-30等在内的12份材料。这类种质群株高较高,穗长较长,穗下茎长较长,穗下茎长与株高比中等,穗叶距中等,主穗小穗数较多,主穗粒数较多,单株粒质量较大,穗轮层数较多,茎节数较多,茎秆颜色为黄色和白色,籽粒茸毛为少毛和中毛。
穗色、外稃色和籽粒饱满度在6类种质群中没有表现出明显差异。
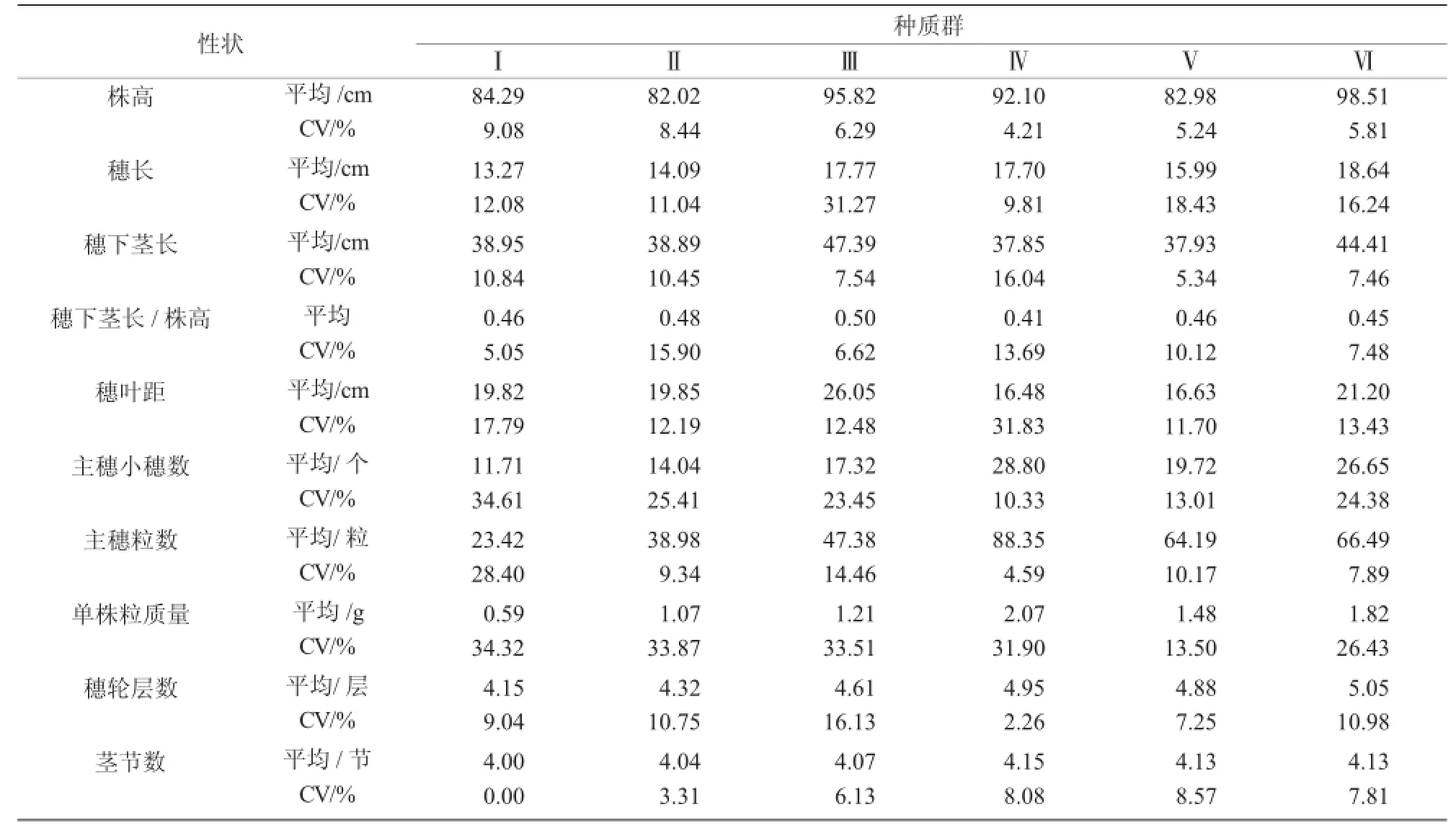
表4 燕麦种质资源各类群的生物学性状
3 讨论
种质资源的遗传多样性是育种工作的基础。育种进程的不断加快使得种质资源相对集中[13-15],为了防止新育成品种主要性状的变异幅度持续变窄,需要对育种材料的遗传多样性进行提高[16-18]。本试验对289份燕麦种质的22个生物学性状进行了遗传变异分析。其中,芒性、芒型、内稃色和粒型等外观农艺性状以及对于燕麦产量起关键作用的主穗粒数、单株粒质量和主穗小穗数的遗传差异较大,遗传多样性更加丰富。因此,在改良燕麦产量性状上,这些种质资源具有较大的潜力。
依据22个性状对68份具有代表性的供试燕麦资源进行聚类分析。根据性状相近原则,整体将68份燕麦资源划分为6类。聚类分析结果可明确不同燕麦资源有哪些优良性状,结合实际育种目标,选择性状亲本配置组合,大大提高选配的合理性。
聚类得到的6类种质群根据育种目标可分为不同类型。种质群Ⅰ,Ⅱ可作为燕麦矮化品种亲本,但这2类种质群产量及其他性状较差。种质群Ⅲ可作为培育抗旱和饲草品种的基因库。种质群Ⅳ可作为改良产量性状的基因库。种质群Ⅴ无突出优异性状。种质群Ⅵ可作为以株高和穗长为育种目标的燕麦亲本。在选择区分种质群性状中,穗色、外稃色和籽粒饱满度表现较差,不可作为区分种质群的性状。
由于生物学性状受到遗传、自然环境和人为等多方面的作用,单从外观农艺性状很难对种质资源间的遗传变异作出准确的分析。因此,依据燕麦资源的表型差异,利用细胞生物学和分子生物学等方法开展更深入的研究,发掘燕麦资源中的优良基因,进行基因定位,为分子辅助育种提供标记,从而加快高产、优质燕麦新品种的选育进程。
[1]杨海鹏,孙泽民.中国燕麦[M].北京:农业出版社,1989.
[2]赵世锋,田长叶,王志刚,等.我国燕麦生产和科研现状及未来发展方向[J].杂粮作物,2007,27(6):428-433.
[3]刘龙龙,崔林,刘根科,等.山西省燕麦产业现状及技术发展需求[J].山西农业科学,2010,38(8):3-5,12.
[4]马艳明,刘志勇,白玉亭,等.新疆燕麦地方品种资源多样性分析[J].新疆农业科学,2006,43(6):510-513.
[5]Zhou X,Jellen E N,Murphy J P.Progenitor germplasm of domesticated hexaploid oat[J].Crop Science,1999,39:1208-1214.
[6]Jin H.Combined AFLP and RFLP mappingin twohexaploid oat recombinant inbred populations[J].Genome,2000,43:94-101.
[7]周竹青,朱旭彤,王维金.影响小麦粒质量的农艺性状、生理指标的主成分分析[J].生物数学学报,2002,17(1):92-96.
[8]王亚娟,张秋芳,任志龙,等.小麦优异种质资源农艺性状综合鉴定与评价[J].麦类作物学报,2004,24(4):119-122.
[9]张极英,张丽娟.大豆种质资源的分类鉴定研究[J].中国油料作物学报,2002,24(1):33-37.
[10]杨守臻,李初英,陈怀珠,等.广西春大豆地方品种农艺性状鉴定及聚类分析[J].广西农业科学,2005,36(1):71-74.
[11]沈强云,许志斌.宁夏春小麦品种主要农艺性状的遗传相关及主成分分析[J].宁夏农林科技,1997(4):7-9.
[12]郝黎仁,樊元,郝哲欧,等.SPSS实用统计分析[M].北京:中国水利水电出版社,2002:280-285.
[13]庄萍萍,李伟,魏育明,等.波斯小麦农艺性状相关性及主成分分析[J].麦类作物学报,2006,26(4):11-14.
[14]孔德伟,陈德全,周良强,等.杂交水稻几个重要农艺及产量性状的主成分分析[J].中国农学通报,2006,21(8):117-119.
[15]韩秉进,潘相文,金剑,等.大豆农艺及产量性状的主成分分析[J].大豆科学,2008,27(1):67-73.
[16]Wang L,Guan Y,Guan R,et al.Establishment of Chinese soybean Glycine max core collections with agronomic traits and SSR markers[J].Euphytica,2006,151(2):215-223.
[17]刘三才,郑殿升,曹永生,等.中国小麦选育品种与地方品种的遗传多样性[J].中国农业科学,2000,33(4):20-24.
[18]周建萍,刘龙龙,崔林.山西省燕麦育种现状及资源特点[J].山西农业科学,2010,38(11):6-9.
Diversity Analysis on Biological Characters of Oat Resources
MUZhixin,LIULonglong,ZHANGLijun,MAMingchuan,ZHOUJianping,CUI Lin
(Institute ofCrop GermplasmResources,Shanxi AcademyofAgricultural Sciences,KeyLaboratoryofCrop Gene Resources and GermplasmEnhancement on Loess Plateau,MinistryofAgriculture,Taiyuan 030031,China)
To enrich the genetic basis of oat breeding materials in Shanxi province,the diversity of 22 biological characters of 289 oat germplasm was investigated.The results showed that the genetic diversity index of every character was bigger,and the diversity of germplasms was much abundant.According to genetic difference of every characteristic among varieties,cluster analysis indicated that the 68 germplasm which had representation for the 289 germplasm could be classified into 6 clusers at distance coefficients of 3.8,9 germplasm were classified intoⅠcluster and 12 germplasm were classified intoⅡcluster,these two clusers could be as parent for breeding dwarfted varieties.22 germplasm were classified intoⅢcluster and this cluster could be as parent for breeding forage grass.5 germplasm were classified intoⅣcluster and this cluster could be as parent for breeding high yield oat.8 germplasm were classified intoⅤcluster,but the beneficial traits were not be observed.12 germplasm were classified intoⅥcluster and this cluster could be as parent ofplant height and spike length breedingobjectives.
oat;resource;biological characters;cluster analysis
S512.6
A
1002-2481(2016)12-1751-05
10.3969/j.issn.1002-2481.2016.12.02
2016-10-24
国家燕麦荞麦产业技术体系(CARS-08-A-1);山西省科技基础条件平台建设项目(2014091003-0103);山西省重点研发计划项目(201603D221004-1)
穆志新(1974-),男,山西浑源人,副研究员,主要从事农作物种质资源研究工作。
猜你喜欢
中老年保健(2022年5期)2022-11-25
今日农业(2022年13期)2022-09-15
现代畜牧科技(2021年10期)2021-11-19
今日农业(2021年7期)2021-07-28
中国麻业科学(2018年6期)2018-04-09
延河(2017年7期)2017-07-19
初中生世界·八年级(2017年3期)2017-03-24
西南农业学报(2016年5期)2016-05-17
中学生数理化·七年级数学人教版(2016年6期)2016-05-14
广西林业科学(2016年3期)2016-03-16
