
罗马洋甘菊HMGR基因的克隆与表达分析
2017-01-17 09:57孟想想闫佳萍刘晓梦廖咏玲
华北农学报 2016年6期
孟想想,闫佳萍,刘晓梦,廖咏玲,常 杰,许 锋
(1.长江大学 园艺园林学院,湖北 荆州 434025;2.靶向抗肿瘤药物湖北省协同创新中心,湖北 荆门 448000;3.荆楚理工学院 化工与药学院,湖北 荆门 448000)
罗马洋甘菊HMGR基因的克隆与表达分析
孟想想1,闫佳萍1,刘晓梦1,廖咏玲1,常 杰2,3,许 锋1
(1.长江大学 园艺园林学院,湖北 荆州 434025;2.靶向抗肿瘤药物湖北省协同创新中心,湖北 荆门 448000;3.荆楚理工学院 化工与药学院,湖北 荆门 448000)
3-羟基-3-甲基戊二酰-CoA还原酶(3-hydroxy-3-methyl glutaryl coenzyme A reductase,HMGR)是植物萜类化合物甲羟戊酸合成途径中的关键限速酶。为研究HMGR基因在洋甘菊萜类化合物合成代谢中的功能,根据前期罗马洋甘菊转录组注释HMGR的Unigene序列设计引物,以罗马洋甘菊cDNA为模板,采用RT-PCR方法,从罗马洋甘菊中克隆得到一个长为1 856 bp的HMGR基因,命名为CnHMGR(GenBank登录号为KU589282)。CnHMGR基因序列包含1个1 746 bp长的开放阅读框,编码582个氨基酸,生物信息学软件预测蛋白质分子量和等电点分别为62.5 kDa和6.80。氨基酸序列多重比对结果显示,CnHMGR蛋白质与其他植物的HMGR蛋白具有高度相似性,并包含HMGR蛋白质家族中的2个HMG-CoA结合基序:TTEGCLUA、EMPVGYVQIP以及2个NADP(H)结合基序:TVGGGT、DAMGMNM,表明CnHMGR属千罗马洋甘菊HMGR家族成员。组织表达分析显示CnHMGR在罗马洋甘菊的不同组织均有表达,在花中表达量最高,在茎中表达量最低。通过对CnHMGR基因的克隆及表达特性分析,为后续深入研究其在洋甘菊倍半萜合成途径中的功能奠定了理论基础。
罗马洋甘菊;CnHMGR;基因克隆;组织表达
罗马洋甘菊(Chamaemelumnobile)为菊科母菊属草本植物,富含挥发性芳香油[1]。研究表明,洋甘菊挥发油的主要成分为倍半萜类化合物,包括α-甜没药醇、α-甜没药醇氧化物A和B以及母菊薁等活性成分,具有消炎、抑菌、抗氧化性和抗癌等功效,被广泛用于药品、化妆品和香料等领域。尽管洋甘菊挥发油主要从花中提取,但花中仅含有1%~2%的挥发性油[2-3],远不能满足市场需求。因此,提高洋甘菊挥发油中倍半萜类化合物含量,是目前洋甘菊栽培育种研究领域中的重要方向之一。
植物倍半萜类物质生物合成中的C5单位主要来自甲羟戊酸(MVA)途径。MVA途径是合成倍半萜的主要来源,两分子乙酰CoA(Acetyl-CoA)在乙酰CoA转运酶(AACT)作用下形成乙酰CoA(Acetoacetyl-CoA),经羟甲基戊二酰CoA合酶(HMGS)催化形成3-羟基-3-甲基戊二酰CoA(HMG-CoA),再在HMGR催化下形成甲羟戊酸(MVA),之后逐步合成甲羟戊酸-5-磷酸(MVAP)、甲羟戊酸-5-二磷酸(MVAPP)及异戊烯基焦磷酸(IPP),或其异构体二甲丙烯基焦磷酸(DMAPP)[4]。IPP和DMAPP缩合形成牻牛儿基焦磷酸(GPP);GPP与1个IPP 单元结合生成法呢基焦磷酸(FPP)为倍半萜的前体物质,FPP经异构、环化、络合等方式最终形成倍半萜[5]。在MVA途经中,3-羟基-3-甲基戊二酸单酰CoA还原酶(HMGR)催化3-羟基-3-甲基戊二酸单酰辅酶A(HMG-CoA)生成甲羟戊酸(MVA),这一催化过程被认为是控制MVA途径前期反应的关键限速步骤[6],因此HMGR被认为是MVA途径中的关键限速酶之一。大量研究表明:倍半萜类物质的合成与HMGR活性呈正相关,提高植物体内HMGR基因的表达水平可显著提高倍半萜类物质含量[7]。如Chappell和Nable[8]通过外源真菌诱导烟草悬浮细胞的HMGR酶活性,进而促进了倍半萜类化合物的积累。此外,甜椒HMGR2基因表达水平也受外源真菌诱导,同时倍半萜环化酶及法尼基焦磷酸合酶的活性也随之提高,暗示HMGR2基因参与调控甜椒倍半萜类化合物的生物合成[9]。
近年来,科学家已先后从银杏(Ginkgobiloba)[10]、乐昌含笑(Micheliachapensis)[11]、丹参(Salviamiltiorrhiza)[12]、龙眼(Dimocarpuslongan)[13]、蒲公英(Taraxacummongolicum)[14]、露水草(Cyanotisarachnoidea)[15]、积雪草(Centellaasiatica)[16]等植物中分离克隆出HMGR基因,但迄今为止在罗马洋甘菊中尚未见HMGR基因的报道。鉴于HMGR是倍半萜类化合物合成途径中的关键限速酶,因此,本研究以罗马洋甘菊为试材,利用RT-PCR技术克隆CnHMGR基因,并进行了该基因的生物信息学分析,通过实时荧光定量PCR技术检测CnHMGR在不同组织中的表达水平,以期为进一步开展罗马洋甘菊CnHMGR基因的表达调控和定向调节倍半萜类化合物的生物合成奠定理论基础。
1 材料和方法
1.1 材料和试剂
试验材料为种植于长江大学植物园的罗马洋甘菊。本试验中引物核苷酸和PCR产物测定序列工作均委托上海生工生物工程公司完成。切胶回收试剂盒(Agarose Gel DNA purifieation Kit Ver.4.0)、RNA提取试剂盒(MiniBEST Plant RNA Extraction kit)、反转录试剂盒(PrimeScriptTM1st Strand cDNA Synthesis Kit)、实时定量试剂盒(PrimeScriptTMRT reagent Kit Perfect Real Time)、pMD18-T载体、dNTP、RNA酶和TaqDNA聚合酶均购自宝生物工程(大连)有限公司。
1.2 罗马洋甘菊CnHMGR基因的克隆
取罗马洋甘菊的花组织样品2~3 g,使用液氮研磨成粉状,并采用TaKaRa MiniBEST Plant RNA Extraction kit试剂盒提取总RNA,采用PrimeScriptTM1st Strand cDNA Sythesis Kit(TaKaRa,大连)试剂盒将提取RNA反转录成cDNA。根据许锋课题组前期罗马洋甘菊转录组注释的HMGRUnigene序列,设计一对特异引物DFHMGR(5′-CCCACCCCTTCCT
TTGAAAA-3′)和DRHMGR(5′-GGATCAGAAGATGA
TGCCCACA-3′),利用反转录PCR技术(RT-PCR)扩增HMGR基因。PCR扩增产物经过凝胶电泳初步检测后切胶回收,进一步纯化后与pMD-18T载体进行连接,转化进大肠杆菌株DH5α,挑选阳性克隆送上海生工生物工程公司测序。
1.3 生物信息学分析
经测序成功的HMGR基因序列,利用NCBI网站上的在线软件Blast P(http://www.ncbi.nlm.nih.gov/BLAST/)进行DNA序列相似性比较及蛋白质序列分析,利用软件Vector NTI 11.5预测CnHMGR基因的开放读码框(ORF)。采用在线工具ExPASy(http://web.expasy.org)预测CnHMGR编码蛋白的理化性质。利用软件Vector NTI 11.5将CnHMGR蛋白与其他植物HMGR的蛋白质进行多重比对,通过Clustal X2.0和MEGA5构建HMGR蛋白质的系统进化树。
1.4 实时荧光定量PCR分析
分别提取罗马洋甘菊根、茎、叶、花的总RNA,反转录成cDNA。根据CnHMGR的cDNA序列设计实时荧光定量PCR特异引物,上游引物为5′-AGAAGAAACCACCTCCAACTATCG-3′,下游引物为5′-CTTCCAGAGGCAATCCTGATAGAG-3′;内参基因选用18S基因,基因上游引物为5′-TTGGTCTCC CGTGCTAATGG-3′,下游引物为5′-CGAAGCGTC ATCCTAAGACAACA-3′。参照TaKaRa公司的SYBR®Premix ExTaqⅡ(TliRNaseH Plus)试剂盒说明书,在Bio-Rad CFX荧光定量PCR仪上进行实时荧光定量PCR。反应程序为95 ℃,30 s;95 ℃,5 s,60 ℃,30 s,共40个循环,加溶解曲线程序,每个部位均进行3次重复,基因相对表达量值通过2-ΔΔCt法计算[17],各组织表达水平以茎中的表达量为对照(CK)。

起始密码子和终止密码子通过方框标出;引物通过下划线标出。The initial codon and the stop codon are highlighted in square box;The primer sequences are underlined.
2 结果与分析
2.1CnHMGR全长cDNA克隆和序列分析
根据本课题组前期罗马洋甘菊转录组注释的HMGRUnigene序列设计引物,利用RT-PCR扩增得到长为1 856 bp的cDNA序列。经NCBI网站的Blast程序在线比对分析显示,该cDNA序列与其他植物的HMGR序列高度同源,表明克隆的cDNA序列为罗马洋甘菊HMGR基因,因此,将该cDNA序列命名为CnHMGR,GenBank登录号为KU589282。CnHMGR基因的cDNA序列全长1 856 bp,包含1 746 bp的开放阅读框,编码582个氨基酸(图1)。2.2 CnHMGR蛋白质分析ExPASy在线分析结果显示,CnHMGR基因编码的蛋白质分子量为62.5 kDa,理论等电点为6.80。利用软件Vector NTI11.5将CnHMGR蛋白质序列与其他植物HMGR蛋白序列进行多重比对,结果显示CnHMGR与其他植物的HMGR蛋白序列高度同源(图2)。如表1所示,CnHMGR与苍术(Atractylodeslancea)、三七(Panaxnotoginseng)、甜瓜(Cucumismelo)、荔枝(Litchichinensis)、橡胶树(Heveabrasiliensis)、莲花(Nelumbonucifera)、葡萄(Vitisvinifera)、黄瓜(Cucumissativus)、蓖麻(Ricinuscommunis)HMGR蛋白质的相似度分别为85%,78%,80%,79%,79%,78%,78%,77%,77%,进一步验证了CnHMGR为罗马洋甘菊HMGR基因家族成员之一。进一步的序列分析结果显示,CnHMGR蛋白包含4个保守的结构域,分别为2个NADP(H)结合域:TVGGGT、DAMGMNM,以及2个HMG-CoA结合域:TTEGCLVA、EMPVGYVQIP,暗示CnHMGR也具有与其他植物HMGR蛋白酶相似的催化功能[18]。
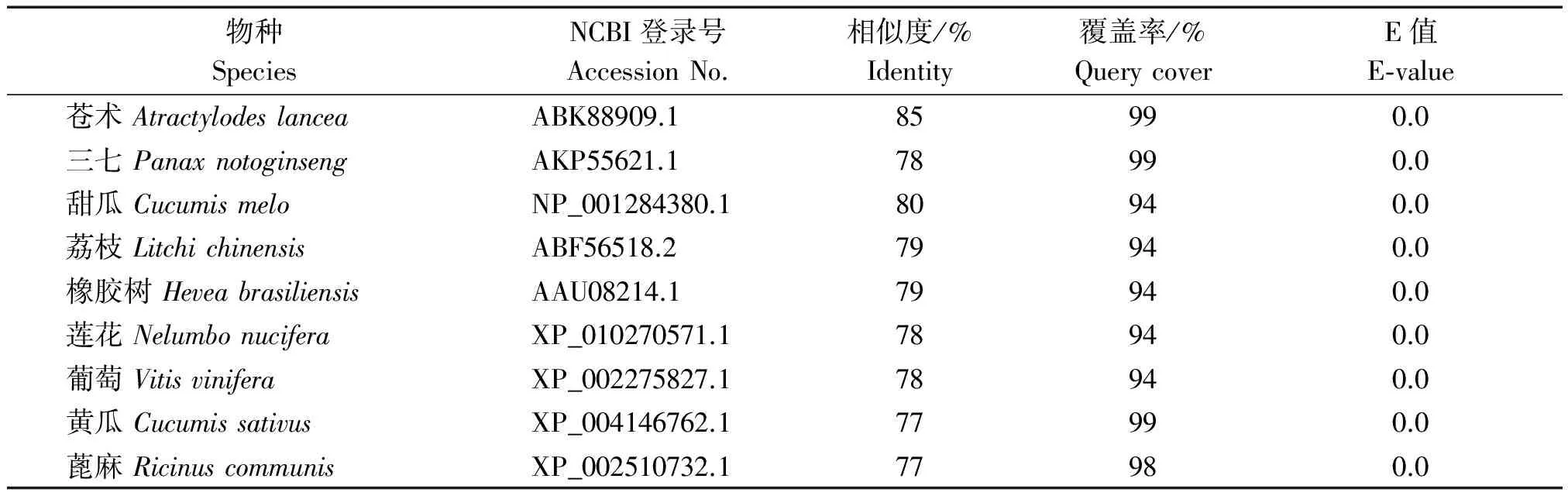
表1 CnHMGR与其他植物HMGR蛋白质序列的相似性比对Tab.1 Protein sequence of CnHMGR similarity to HMGRs of other plant species


保守的HMG-CoA结合基序与NADP(H)结合基序用黑框标出。The conserved NADP(H)-binding motifs and HMG-CoA-binding domains is highlighted in black square box.
2.3 CnHMGR系统进化分析
利用软件ClustalX 2.0和MEGA 6.0通过邻接法(Neighbor-joining)构建了HMGR的系统进化树。HMGR系统进化树分成两大分支,单子叶植物禾本科和双子叶植物,双子叶植物又被分为菊科、桔梗科、蓝果树科、五加科、茄科、玄参科、唇形科、十字花科、无患子科和大戟科。在HMGR系统进化上,罗马洋甘菊与双子叶植物的亲缘关系较单子叶植物近,与菊科植物万寿菊(Tageteserecta)亲缘关系最近、与桔梗(Platycodongrandiflorus)、喜树(Camptothecaacuminata)、三七(Panaxnotoginseng)和人参(Panaxginseng)也有很高的亲缘性(图3),这一方面反映了CnHMGR基因在双子叶植物中进化保守,另一方面也反映了植物HMGR基因在进化上的多样性,这与植物形态学分类情况是一致的。
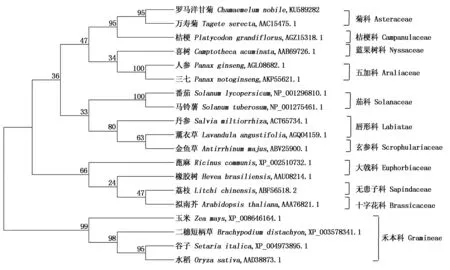
图中各分支数值代表置信度。The number shown at each branch indicates the bootstrap values.
2.4CnHMGR组织表达分析
提取罗马洋甘菊的根、茎、叶、花的总RNA,利用实时荧光定量PCR技术(qRT-PCR)测定了罗马洋甘菊各器官中CnHMGR基因的表达水平(图4)。结果表明,CnHMGR基因在罗马洋甘菊的根、茎、叶、花中均有表达,其中在花中的表达量最高,其次是根和叶,而在茎中表达量最低。
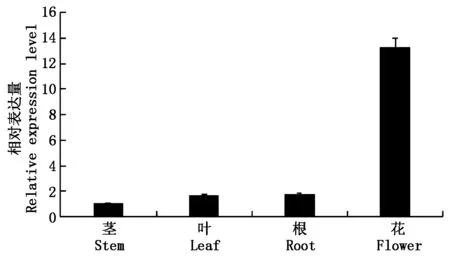
图4 CnHMGR在罗马洋甘菊不同组织的表达水平Fig.4 Expression pattern of CnHMGR among different tissues of Chamaemelum nobile
3 讨论
倍半萜在植物体内多以醇、酮、苷或内酯的形式存在,罗马洋甘菊中的倍半萜化合物主要有α-甜没药醇及其氧化物,以及母菊薁、金合欢烯等,药理学研究表明倍半萜类物质具有抗菌、抗氧化、消炎、抗痉挛、抗癌、抗肿瘤、抗神经毒性、保肝等生物活性,是医药、食品、化妆品工业的重要原料[11,19]。研究表明倍半萜类化合物生物合成主要通过甲羟戊酸(MVA)途径,HMGR是甲羟戊酸(MVA)合成途径中的关键基因,对甲羟戊酸代谢“碳流”的调控起重要作用,改变HMGR活性可有效调控萜类化合物含量。利用转基因技术将HMGR基因在植物中超表达可提高倍半萜类化合物含量[7,20-21]。例如马铃薯HMGR2和HMGR3的超表达可使与倍半萜植保素含量增加[22-23],长春花HMGR基因在黄花蒿中过表达,可使倍半萜类物质青蒿素含量提高22.5%[24]。进一步研究发现,MVA途径控制了黄花蒿萜类代谢途径中80%的碳源,且HMGR作为该途径的主要限速酶,其酶活性的调节直接影响了细胞中甲羟戊酸的储蓄和青蒿素的积累[25]。鉴于此,本研究克隆了罗马洋甘菊中倍半萜合成途径中的关键基因HMGR基因,并分析了该基因的组织表达模式,可为利用基因工程技术提高罗马洋甘菊倍半萜类化合物含量提供基因资源与理论基础。
生物信息学表明CnHMGR蛋白与其他植物的HMGR蛋白具有高度的同源性,并包含4个保守基序,即NADPH结合基序:TVGGGT、DAMGMNM和HMG-CoA结合基序:TTEGCLVA、EMPVGYVQIP。CnHMGR蛋白的HMG-CoA结合基序中TTEGCLVA中的谷氨酸(Glu)可能在HMGR的催化功能发挥着重要作用。研究表明,HMG-CoA结合基序(EMPVGYVQIP)在各植物之间的多样性对于HMGR催化底物的选择可能发挥着重要作用[18,26]。CnHMGR包含该保守基序,推测也具有底物选择的功能。通过进化树分析结果可见,罗马洋甘菊与万寿菊亲缘关系最近,推测CnHMGR与万寿菊HMGR蛋白具有相似的催化活性,罗马洋甘菊与其他双子叶、及单子叶植物也具有亲缘关系,反映了植物HMGR基因在进化上的多样性,同时也表明HMGR是萜类生物合成途径中1个十分保守的基因。利用邻接法构建的HMGR系统进化树虽然只能在一定程度上反映各物种之间的进化关系,但对物种间亲缘关系的判定仍具有一定的参考价值。
研究表明,HMGR基因在不同植物组织表达模式呈现出较大的差异,例如,在南非醉茄(Withaniasomnifera)中表达量最高,花中表达量最少[27];在人参花中表达量最高,茎中表达量最低[28];在露水草中根和叶表达丰富[15];在积雪草节点表达最高,在根部表达最低[16]。HMGR基因的特异性表达通常与类固醇或倍半萜积累相关,如在丹参[12]、银杏[10]、黄花蒿[24]、白木香[29]中,HMGR的表达量分别与银杏内酯和白果内酯、青蒿素、沉香倍半萜、丹参酮的含量呈正相关。本研究中实时定量PCR检测结果显示CnHMGR在花中的表达量最高。该结果表明CnHMGR在罗马洋甘菊中的表达具有组织特异性,该表达模式可能和罗马洋甘菊倍半萜的合成与积累部位有关。已有文献报道罗马洋甘菊的花器官中富含精油及倍半萜类化合物,其含量显著高于其他组织[30],CnHMGR在花中的表达水平最高,可能与倍半萜类化合物的合成呈正相关,暗示了HMGR可能具有调控倍半萜化合物的功能。
本课题组前期的转录组测序结果显示罗马洋甘菊HMGR基因家族中存在众多成员,本研究克隆的CnHMGR基因仅为其家族中的一个成员。因此,在后续研究中,需进一步对罗马洋甘菊HMGR基因家族的其余成员进行克隆及功能分析,并研究它们在催化功能上的异同点,为后期开展罗马洋甘菊HMGR基因家族调控倍半萜类化合物生物合成的分子机制研究提供基础理论。
[1] Lemberkovics E, Kery A, Marczal G, et al. Phytochemical evaluation of essential oils,medicinal plants and their preparations[J]. Acta Pharmaceutica Hungarica, 1998, 68(3): 141-149.
[2] Carnat A, Carnat A P, Fraisse D, et al. The aromatic and polyphenolic composition of Roman chamomile tea[J]. Fitoterapia, 2004, 75(1): 32-38.
[3] Srivastava J K, Shankar E, Gupta S. Chamomile: A herbal medicine of the past with a bright future (Review)[J]. Molecular Medicine Reports, 2010, 3(6): 895-901.
[4] 张长波, 孙红霞, 巩中军, 等. 植物萜类化合物的天然合成途径及其相关合酶[J]. 植物生理学通讯, 2007, 43: 779-786.
[5] 何云飞, 高 伟, 刘塔斯, 等. 二萜合酶的研究进展[J]. 药学学报, 2011, 46( 9): 1019.
[6] Doblas V G, Amorim-Silva V, Posé D, et al. The SUD1 gene encodes a putative E3 ubiquitin ligase and is a positive regulator of 3-hydroxy-3-methylglutaryl coenzyme A reductase activity in Arabidopsis[J]. Plant Cell, 2013, 25(2): 728-743.
[7] Chappell J, Wolf F, Proulx J, et al. Is the reaction catalyzed by 3-hydroxy-3-methylglutaryl coenzyme A reductase a rate-limiting step for isoprenoid biosynthesis in plants[J]. Plant Physiology, 1995, 109(4):1337-1343.
[8] Chappell J, Nable R. Induction of sesquiterpenoid biosynthesis in tobacco cell suspension cultures by fungal elicitor[J]. Plant Physiology, 1987, 85(2):469-473.
[9] Ha S H, Kim J B, Hwang Y S, et al. Molecular characterization of three 3-hydroxy-3-methylglutaryl-CoA reductase genes including pathogen-induced Hmg2 from pepper(Capsicumannuum)[J]. Biochimica et Biophysica acta, 2003, 1625(3): 253-260.
[10] Shen G, Pang Y, Wu W, et al. Cloning and characterization of a root-specific expressing gene encoding 3-hydroxy-3-methylglutaryl coenzyme A reductase fromGinkgobiloba[J]. Molecular Biology Reports, 2006, 33(2): 117-127.
[11] Cao X Y, Li C G, Miao Q, et al. Molecular cloning and expression analysis of a leaf- specific expressing 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) reductase gene fromMicheliachapensisDandy[J]. Journal of Medicinal Plant Research, 2011, 5(16): 3868-3875.
[12] Dai Z, Cui G, Zhou S F, et al. Cloning and characterization of a novel 3-hydroxy-3-methylglutaryl coenzyme A reductase gene fromSalviamiltiorrhizainvolved in diterpenoid tanshinone accumulation[J]. Journal of Plant Physiology, 2011, 168(2): 148-157.
[13] Xia R, Lu W J, Wang Z H, et al. Cloning and characterisation of two genes for 3-hydroxy-3-methylglutaryl coenzyme A reductase and their possible roles during fruit development inDimocarpuslongan[J]. The Journal of Horticultural Science and Biotechnology, 2011, 86(1): 25-30.
[14] Van D N, Bachmann A L, Schmidt T, et al. Molecular cloning of mevalonate pathway genes fromTaraxacumbrevicorniculatumand functional characterisation of the key enzyme 3-hydroxy-3-methylglutaryl-coenzyme A reductase[J]. Molecular Biology Reports, 2012, 39(4): 4337-4349.
[15] Wang Q J, Zheng L P, Zhao P F, et al. Cloning and characterization of an elicitor-responsive gene encoding 3-hydroxy-3-methylglutaryl coenzyme A reductase involved in 20-hydroxyecdysone production in cell cultures ofCyanotisarachnoidea[J]. Plant Physiology and Biochemistry, 2014, 84: 1-9.
[16] Kalita R, Patar L, Shasany A K, et al. Molecular cloning, characterization and expression analysis of 3-hydroxy-3-methylglutaryl coenzyme A reductase gene fromCentellaasiaticaL.[J]. Molecular Biology Reports, 2015, 42(9): 1431-1439.
[17] Livak K J, Schmittgen T D. Analysis of relative gene expression data using Real-time quantitative PCR and the 2-ΔΔCtmethod[J]. Methods, 2001, 25(4): 402-408.
[18] Wang Y, Darnay B G, Rodwell V W. Identification of the principal catalytically important acidic residue of 3-hydroxy-3-methylglutaryl coenzyme A reductase[J]. The Journal of Biological Chemistry, 1990, 265(35): 21634-21641.
[19] 朴英花, 朴惠顺. 倍半萜类化合物生物活性研究进展[J]. 职业与健康, 2012, 28(18): 2291-2293.
[20] Chappell J. Biochemistry and molecular biology of the isoprenoid biosynthetic pathway in plants[J]. Annual Review of Plant Biology, 1995, 46(1): 521-547.
[21] Darabi M, Masoudi-Nejad A, Nemat-Zadeh G. Bioinformatics study of the 3-hydroxy-3-methylglotaryl-coenzyme A reductase (HMGR) gene in Gramineae[J]. Molecular Biology Reports,2012,42(9):1243-1243.
[22] Yang Z, Park H, Lacy G H, et al. Differential activation of potato 3-hydroxy-3-methylglutaryl coenzyme A reductase genes by wounding and pathogen challenge[J]. The Plant Cell, 1991, 3(4): 397-405.
[23] Choi D, Ward B L, Bostock R M. Differential induction and suppression of potato 3-hydroxy-3-methylglutaryl coenzyme A reductase genes in response to Phytophthora infestans and to its elicitor arachidonic acid[J]. The Plant Cell, 1992, 4(10): 1333-1344.
[24] Aquil S, Husaini A M, Abdin M Z, et al. Overexpression of the HMG-CoA reductase gene leads to enhanced artemisinin biosynthesis in transgenicArtemisiaannuaplants[J]. Planta Medica, 2009, 75(13): 1453-1458.
[25] Alam P, Abdin M Z. Over-expression of HMG-CoA reductase and amorpha-4,11-diene synthase genes in Artemisia annua L. and its influence on artemisinin content[J]. Plant Cell Reports, 2011, 30(10): 1919-1928.
[26] Ruiz-Albert J, Cerdá-Olmedo E, Corrochano L M. Genes for mevalonate biosynthesis in Phycomyces[J]. Molecular Genetics and Genomics, 2002, 266(5): 768-777.
[27] Akhtar N, Gupta P, Sangwan N S, et al. Cloning and functional characterization of 3-hydroxy-3-methylglutaryl coenzyme A reductase gene fromWithaniasomnifera:an important medicinal plant[J]. Protoplasma, 2013, 250(2): 613-622.
[28] 罗红梅, 宋经元, 李雪莹, 等. 人参皂苷合成生物学关键元件HMGR基因克隆与表达分析[J].药学学报, 2013, 48(2): 219-227.
[29] 徐艳红, 杨 欣, 张 争, 等. 白木香3-羟基-3-甲基戊二酰辅酶A还原酶基因-AsHMGR2的克隆及表达分析[J]. 药学学报, 2013, 48(6): 953.
[30] Agatonovic-Kustrin S, Ortakand D B, Morton D W. Migraine headaches:Feverfew or Chamomile leaves[J]. Modern Chemistry Applications, 2015, 3(169): 2.
Cloning and Expression Analysis of HMGR Gene from Chamaemelum nobile
MENG Xiangxiang1,YAN Jiaping1,LIU Xiaomeng1,LIAO Yongling1,CHANG Jie2,3,XU Feng1
(1.College of Horticulture and Landscape Architecture,Yangtze University,Jingzhou 434025,China;2.Hubei Collaborative Innovation Center of Targeted Antitumor Drug,Jingmen 448000,China;3.College of Chemical Engineering and Pharmacy,Jingchu University of Technology,Jingmen 448000,China)
3-hydroxy-3-methyl-glutaryl-CoA reductase (3-hydroxy-3-methyl glutaryl coenzyme A reductase,HMGR) is one of the key rate-limiting enzymes in mevalonic (MVA) pathway of terpenoid biosynthesis.To analysis the function ofHMGRgene in terpenoid biosynthesis inChamaemelumnobile,aHMGRgene (designated asCnHMGR,GenBank accession number KU589282) was cloned fromC.nobileusing RT-PCR method.The full-length cDNA ofCnHMGRgene was 1 856 bp and contained an open reading frame (ORF) of 1 746 bp,which encodes a 582 amino-acid protein.The theoretical molecular weight and PI of theCnHMGRwere 62.5 kDa and 6.80,respectively.Multi-alignment comparison analysis showed the protein sequence of CnHMGR had high similarity with those of HMGR proteins from other plants.Furthermore,CnHMGR has two HMG-CoA-binding motifs:TTEGCLVA,EMPVGYVQIP,and two NADP(H)-binding motifs:TVGGGT,DAMGMMM,suggesting CnHMGR is one of HMGR family mumbers inC.nobile.The results of tissue expression analysis showed thatCnHMGRwas constituently expressed indifferent tissues ofC.nobile,with highest expression level in flowers and lowest expression level in stems.The present study cloned,characterized and analyzed the tissue expression pattern ofCnHMGRinC.nobile,providing basic data for further studying the function ofCnHMGRin sesquiterpene biosynthesis ofC.nobile.
Chamaemelumnobile;CnHMGR;Gene cloning;Tissue expression
2016-06-05
国家自然科学基金项目(31400603);大学生创新创业训练计划项目(201207014)
孟想想(1991-),女,安徽蚌埠人,在读硕士,主要从事植物分子生物学研究。
许 锋(1979-),男,湖北武汉人,教授,博士,主要从事药用植物次生代谢研究。
Q78;S682.03
A
1000-7091(2016)06-0068-08
10.7668/hbnxb.2016.06.011
猜你喜欢
中国化妆品(2021年12期)2021-12-23
世界科学技术-中医药现代化(2021年12期)2021-04-19
中成药(2020年2期)2020-05-12
环球时报(2020-03-23)2020-03-23
中成药(2018年12期)2018-12-29
中成药(2017年10期)2017-11-16
中南大学学报(自然科学版)(2016年2期)2017-01-19
椰城(2016年10期)2016-11-07
合成化学(2015年1期)2016-01-17
——青蒿素
中国学术期刊文摘(2015年21期)2015-12-24
