
毛竹快速生长过程中碳水化合物的时空变化
2017-04-17 01:43程路芸马丹丹李丹丹许馨露张汝民
浙江农林大学学报 2017年2期
程路芸,温 星,马丹丹,李丹丹,许馨露,高 岩,张汝民
(浙江农林大学 林业与生物技术学院,浙江 临安311300)
毛竹快速生长过程中碳水化合物的时空变化
程路芸,温 星,马丹丹,李丹丹,许馨露,高 岩,张汝民
(浙江农林大学 林业与生物技术学院,浙江 临安311300)
为了探讨毛竹Phyllostachys edulis快速生长过程碳水化合物的变化,采用试剂盒和3,5-二硝基水杨酸比色法,分别测定黄昏(0 h),黄昏后4 h和8 h等3个时间的毛竹笋竹上部、中部、下部和竹蔸中蔗糖、葡萄糖、果糖、淀粉和纤维素质量分数。结果表明:毛竹快速生长过程中不同部位碳水化合物质量分数存在差异,并随时间有不同程度的变化。黄昏开始经过8 h后,各部位淀粉质量分数均显著下降,竹兜和笋竹中部分别下降了28.7%和61.5%;竹蔸的蔗糖、葡萄糖和果糖分别下降29.4%,39.2%和21.4%;笋竹中部蔗糖、葡萄糖和果糖分别下降36.9%,54.3%和26.6%; 4 h后笋竹下部纤维素质量分数增加25.5%。竹蔸、下部和中部中淀粉质量分数与蔗糖、葡萄糖和果糖之间均存在显著正相关,上部淀粉与葡萄糖和果糖质量分数之间存在显著负相关;下部纤维素质量分数与淀粉、蔗糖、葡萄糖和果糖之间均存在显著负相关。黄昏后笋竹下部纤维素沉积较多,中部伸长生长快,消耗可溶性糖较多;笋竹上部伸长生长较慢,对碳水化合物的需求量小;竹蔸作为储藏淀粉的重要部位,为笋竹快速生长提供大量的碳水化合物。图5表4参30
植物生理学;毛竹;碳水化合物;快速生长;时空差异;相关性
碳水化合物是自然界分布最广的物质之一,在植物生命活动中可以构成碳骨架、提供能量、作为运输物质、进行渗透调节和充当信号物质[1]。目前,在很多植物生长过程中碳水化合物的变化已有一定研究。对杂草稻Oryza sativa f.spontanea中胚轴伸长过程的研究发现,发芽后3 d杂草稻可溶性糖迅速增加,发芽后6 d变化趋缓[2];马铃薯Solanum tuberosum萌芽过程中还原糖下降[3];葡萄糖和果糖随甜高粱Sorghum bicolor‘Moench’茎秆节间提高呈两端高中间低的变化规律,蔗糖无明显变化[4]。夜间拟南芥Arabidopsis thaliana植株淀粉逐渐降低;发现拟南芥植株生长过程中的淀粉降解受到生物钟调控[5]。毛竹Phyllostachys edulis是浙江地区重要的经济竹种,在建筑、造纸、手工艺品、食品等领域有广泛应用。竹具有其独特的生长方式即 “爆发式生长”,又称快速生长[6]。前人已从组织结构[7]、光合作用[8]、酶活性[9]、内源激素[10]、蛋白组学[11]、microRNA表达[12]和基因[13]等方面对毛竹快速生长的机制进行了研究和探讨。碳水化合物能为毛竹笋竹快速生长提供结构材料和能量,人们对毛竹中的碳水化合物已有一定探索[14],也有观察表明夜间毛竹笋竹的生长量更大[15],但毛竹笋竹快速生长过程中碳水化合物的变化却鲜见研究。本研究以浙江省临安市的毛竹为对象,从笋竹体内碳水化合物代谢入手,研究夜间笋竹快速生长过程中葡萄糖、果糖、蔗糖、淀粉和纤维素的变化,探究毛竹笋竹快速生长中碳水化合物代谢的生理过程,为研究毛竹快速生长提供理论依据。
1 材料与方法
1.1 供试材料
1.1.1 样地概况 供试材料毛竹采自浙江省临安市浙江农林大学示范基地,地理位置为29°56′~30°27′N,118°51′~119°52′E,属中亚热带季风气候区,四季分明,温暖湿润,年平均气温为16.4℃,气候特点为春多雨、夏湿热、秋气爽、冬干冷,年平均降水量为1 628.6 mm,年平均日照时数为1 847.3 h,森林覆盖率为76.5%。采样地土质为红壤[9]。
1.1.2 实验设计 在2015年4-5月毛竹笋竹快速生长时期,分别于18:00,22:00和02:00(为了更加直观地表示夜间糖质量分数的变化情况,将18:00,22:00和02:00分别定为黄昏0 h,黄昏后4 h和黄昏后8 h),选择长势良好、高度(2.5±0.2)m,大小(基径15.0±2.0)cm相近的毛竹笋竹,从靠近地面的基部伐倒,并挖出竹蔸,将地上笋体部分按照生长高度平均分成3段,由下到上分别记为笋竹的下部、中部和上部。按照生长高度找到位于每段最中间的一节间,从各节间基部1/3处进行取样,并对竹蔸中间位置进行取样。采伐5株·次-1(即5个重复),1株1个独立实验。将毛竹样品迅速放进液氮中冷冻,在-80℃条件下储存。
1.2 测定方法
1.2.1 可溶性糖质量分数测定 取植物样品0.5 g,研磨,加蒸馏水8 mL,80℃恒温水浴30 min,冷却,定容10 mL。离心(3 000 r·min-1,5 min),上清液为可溶性糖提取液。蔗糖、葡萄糖和果糖等3种可溶性糖采用试剂盒(南京建成科技有限公司)进行测定[16]。
1.2.2 淀粉质量分数测定 将提取可溶性糖后残渣用蒸馏水冲洗2遍,去上清液。加入蒸馏水3 mL,搅匀,再加6 mol·L-1的盐酸2 mL,沸水浴30 min,定容25 mL。取上清液,采用3,5-二硝基水杨酸法,在540 nm波长下测其吸光度并计算[17]。
1.2.3 纤维素质量分数测定 将提取淀粉后残渣用蒸馏水冲洗2遍,去上清液,加入体积分数为80%硫酸2 mL,冰浴30 min,再加氢氧化钠调至pH 7.0,定容25 mL。取上清液,用3,5-二硝基水杨酸法,在540 nm波长下测其吸光度并计算[17]。
1.3 数据处理
所有的数据均为5次重复的平均值±标准误差,利用Origin 8软件进行统计分析和作图。统计方法采用One-Way ANOVA,对同一时间不同部位和同一部位不同时间的各种碳水化合物质量分数两两之间分别进行Tukey比较,对不同部位不同碳水化合物的质量分数进行相关性分析。
2 结果与分析
2.1 碳水化合物质量分数的变化
2.1.1 淀粉质量分数的变化 快速生长的毛竹笋竹淀粉质量分数的变化见图1。黄昏时(0 h)竹蔸淀粉质量分数为17.75 mg·g-1,分别比笋竹下部、中部和上部高18.8%,58.6%和61.5%(P<0.05)。8 h后,笋竹竹蔸、下部、中部与上部淀粉质量分数分别比0 h下降了28.7%,47.4%,45.4%与38.5%(P<0.05)。此时,竹蔸淀粉质量分数比笋竹下部、中部及上部高60.9%,107.1%及87.2%(P<0.05)。
2.1.2 蔗糖质量分数变化 如图2,黄昏时竹蔸蔗糖质量分数为19.99 mg·g-1,比笋竹下部低10.4%(P<0.05),与中部相比无显著差异,比上部高出46.4%(P<0.05)。4 h后笋竹竹蔸、下部和中部蔗糖质量分数与黄昏时相比,分别降低了25.2%,35.4%和15.3%,上部不变;黄昏后8 h,中部蔗糖比黄昏后4 h时显著(P<0.05)下降25.5%,其他部位不变。8 h,各相邻部位之间蔗糖质量分数均无显著差异。

图1 笋竹淀粉质量分数的变化Figure 1 Change of bamboo shoots starch content
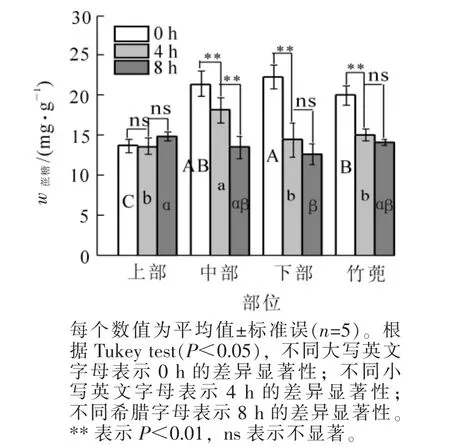
图2 笋竹蔗糖质量分数的变化Figure 2 Change of bamboo shoots sucrose content
2.1.3 葡萄糖质量分数 黄昏0 h,竹蔸中的葡萄糖为18.37 mg·g-1,比笋竹下部低7.2%(P<0.05),与中部无显著差异,比上部高44.8%(P<0.05)。8 h后,笋竹竹蔸、下部和中部葡萄糖质量分数分别下降39.2%,30.8%和54.3%,上部增加22.6%(P<0.05)。这时,竹蔸葡萄糖比笋竹下部和上部分别高出18.5%和28.2%(P<0.05),比中部低28.4%(P<0.05)(图3)。
2.1.4 果糖质量分数 图4显示了快速生长的毛竹笋竹果糖含量的变化。黄昏时候,竹蔸果糖为21.01 mg·g-1,比下部高39.2%(P<0.05);笋竹下部、中部与上部果糖均无显著差异。8 h后,竹蔸、笋竹下部和中部果糖分别降低21.4%,24.5%和26.6%(P<0.05),上部不变。此时,竹蔸果糖与上部无显著差异,分别比笋竹下部和中部分别高出44.9%和64.4%(P<0.05)。
2.1.5 纤维素质量分数 图5中,黄昏时竹蔸纤维素为11.13 mg·g-1,分别是笋竹下部、中部与上部的2.2,4.5与5.8倍(P<0.05)。4 h后笋竹下部纤维素质量分数增加25.5%(P<0.05),黄昏后4 h到8 h变化不显著;黄昏过去8 h后,竹蔸、笋竹的中部和上部纤维素质量分数均无明显变化。黄昏过去8 h后,竹蔸纤维素质量分数分别为下、中和上部的1.6,4.0和5.0倍(P<0.05)。
2.2 相关性分析
2.2.1 笋竹竹蔸碳水化合物的相关性 笋竹竹蔸碳水化合物之间的相关性见表1。淀粉质量分数与蔗糖、葡萄糖和果糖之间均存在显著正相关;蔗糖与葡萄糖和与果糖之间,存在显著正相关;葡萄糖与果糖间存在显著正相关;纤维素质量分数与淀粉、蔗糖、葡萄糖和果糖之间均不存在显著相关性。
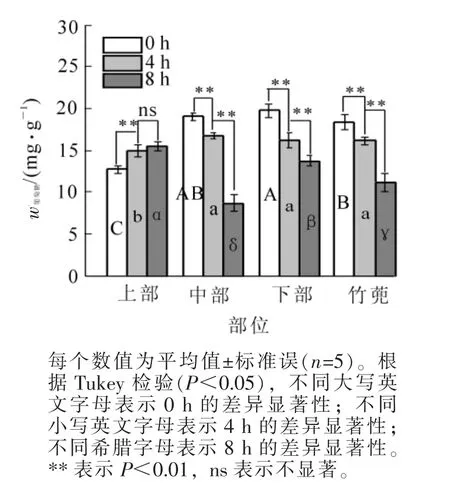
图3 笋竹葡萄糖质量分数的变化Figure 3 Change of bamboo shoots glucose content
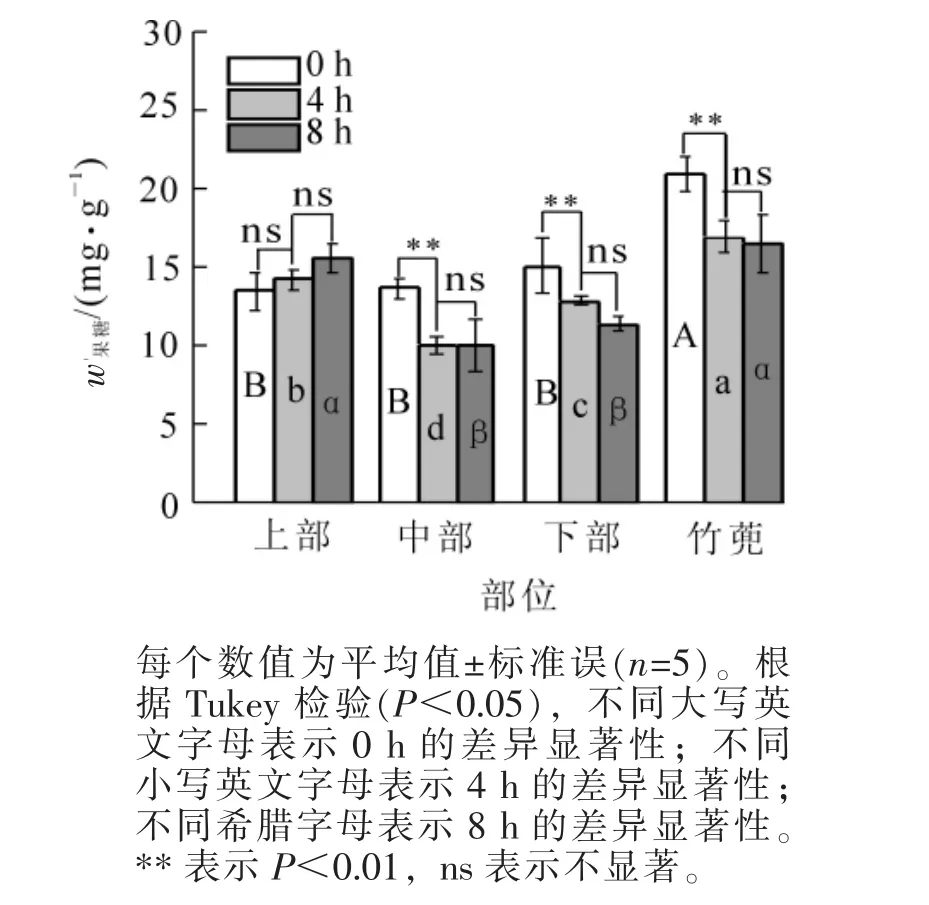
图4 笋竹果糖质量分数的变化Figure 4 Change of bamboo shoots fructose content
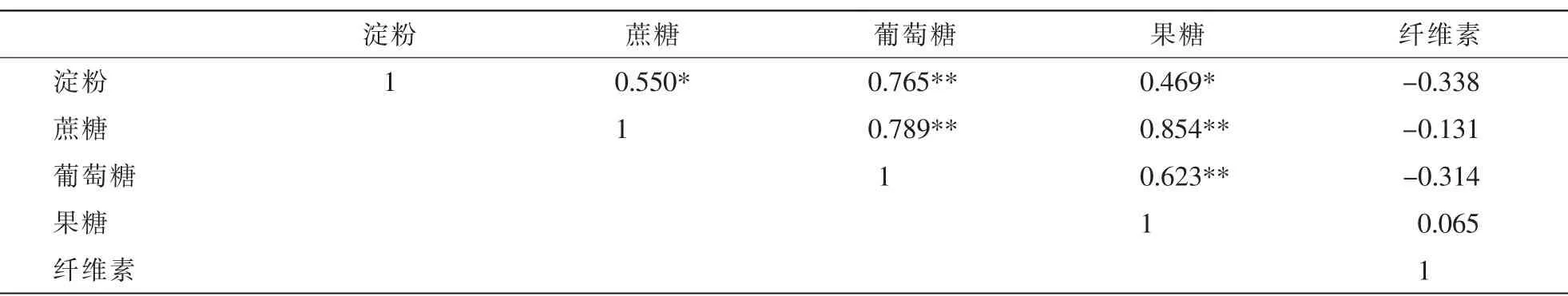
表1 笋竹竹蔸碳水化合物质量分数的相关性Table 1 Correlation of bamboo shoot carbohydrate content in bamboo stump
2.2.2 笋竹下部碳水化合物质量分数的相关性 笋竹下部淀粉与蔗糖、葡萄糖和果糖之间,蔗糖与葡萄糖和果糖,以及葡萄糖与果糖之间均存在显著正相关。纤维素质量分数与淀粉、蔗糖、葡萄糖和果糖之间均存在显著负相关(表2)。

表2 笋竹下部碳水化合物质量分数的相关性Table 2 Correlation of bamboo shoot carbohydrate content in the lower
2.2.3 笋竹中部碳水化合物质量分数的相关性 笋竹中部碳水化合物质量分数的相关性见表3。淀粉与蔗糖、葡萄糖和果糖之间,蔗糖与葡萄糖和果糖之间,已及葡萄糖与果糖之间均存在显著正相关;纤维素质量分数与淀粉、蔗糖、葡萄糖和果糖之间均不存在显著相关性。
2.2.4 笋竹上部碳水化合物质量分数的相关性 淀粉与葡萄糖和果糖质量分数之间存在显著负相关;蔗糖与葡萄糖和果糖之间不存在显著相关;葡萄糖和果糖之间存在显著正相关;纤维素与淀粉质量分数之间存在显著负相关,与葡萄糖质量分数之间存在显著正相关(表4)。
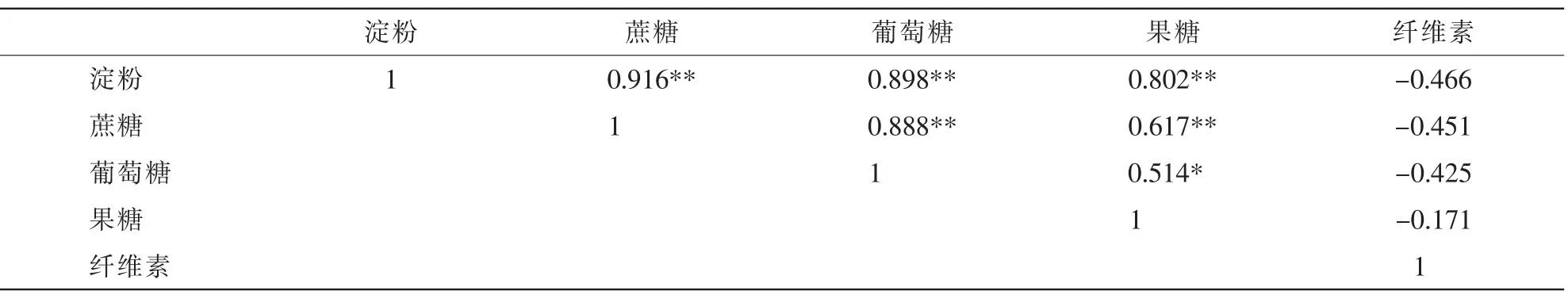
表3 笋竹中部碳水化合物质量分数的相关性Table 3 Correlation of bamboo shoot carbohydrate content in the middle

表4 笋竹上部碳水化合物质量分数的相关性Table 4 Correlation of bamboo shoot carbohydrate content in the upper
3 讨论
碳水化合物是植物光合作用产物和呼吸作用的底物,它为植物生长发育提供能量和碳构架。植物体内碳水化合物组分水平通常可反映植物整体的碳供应水平,表征植物生长状况及其对外界干扰、胁迫的缓冲能力与适应策略[18]。淀粉因其不可移动性通常被视为储备物质,可溶性糖可随时被调动参与植物的各种生理代谢活动[20]。WANG等[21]观察到烟草Nicotiana tabacum幼苗叶片白天合成淀粉,夜晚分解淀粉;GRAF等[5]发现拟南芥叶片中夜间淀粉逐渐降低,并且在黎明时几乎消耗殆尽。本研究中,黄昏时竹蔸淀粉质量分数显著高于笋竹下部、中部和上部,夜间竹蔸和笋竹淀粉均随时间呈下降趋势,与GRAF等[5]的研究结果相似。淀粉在夜间受到糖信号和生物钟的共同调控而逐渐降解[22]。
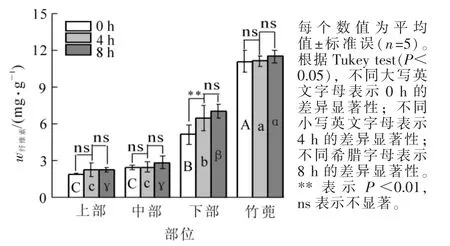
图5 笋竹纤维素质量分数的变化Figure 5 Change of bamboo shoots cellulose content
MATSUDA等[23]发现:夜间番茄Solanum lycopersicum叶片中可溶性糖含量几乎不变;MARTINS等[24]研究发现:拟南芥叶片在黄昏后蔗糖水平下降;刘洋等[4]发现:甜高粱茎秆不同节间之间的蔗糖含量无明显差异,位置靠近中间的节间葡萄糖和果糖含量比位置更靠上部和下部的节间更低。本实验中,黄昏时笋竹中、下部蔗糖和葡萄糖水平比竹蔸和上部高,而笋竹下部、中部和上部的果糖水平显著低于竹蔸;黄昏后竹蔸中蔗糖和己糖的质量分数均随时间下降。竹蔸淀粉与蔗糖、葡萄糖和果糖均显著正相关,蔗糖与葡萄糖和果糖均呈显著正相关,说明竹蔸中淀粉大量降解不断生成不参与运输的葡萄糖和果糖在竹蔸中合成蔗糖后被运输到笋竹[25-26],或者用于维持竹蔸生命。黄昏后,笋竹处于显著伸长期的中、下部可溶性糖质量分数显著减少;下部和中部淀粉与蔗糖、葡萄糖和果糖含量均呈显著正相关,蔗糖与葡萄糖和果糖均呈显著正相关,说明在笋竹中、下部蔗糖被分解为己糖,己糖被大量利用,以供中、下部快速伸长的需要,并为纤维素合成提供底物[27]。笋竹上部的蔗糖和果糖无明显变化,葡萄糖略有积累。上部淀粉与葡萄糖和果糖呈负相关,与蔗糖无显著相关性,表明上部淀粉主要分解生成葡萄糖,转化为果糖,上部可能不进行蔗糖积累。上部主要进行初生生长,细胞伸长较慢[28],对可溶性糖的需求量小。
纤维素是毛竹细胞壁的重要组成成分,被誉为 “细胞壁的骨骼”。在毛竹笋竹快速生长的过程中,纤维素的合成必不可少。本研究中,竹蔸纤维素质量分数变化不明显,竹蔸中纤维素与其他糖类也均无显著相关性,可能由于竹蔸组织较为成熟,纤维素不再进行积累;黄昏后下部纤维素质量分数显著增加。相关性分析的4个部位中,只有笋竹下部纤维素与淀粉、蔗糖、葡萄糖和果糖质量分数均存在显著负相关,这表明黄昏后笋竹下部淀粉降解生成可溶性糖,下部消耗大量可溶性糖,进行次生壁纤维素沉积[19];中部纤维素变化均不显著,纤维素与其他糖类均不存在显著相关性,中部细胞所消耗可溶性糖主要用于伸长生长,仅有初生壁形成,纤维素沉积较少[30],上部纤维素质量分数几乎不变,纤维素与淀粉质量分数显著负相关,与葡萄糖显著正相关,上部主要进行初生生长,纤维素质量分数几乎不变,但略有增加趋势,跟淀粉降解趋势相反,上部淀粉分解产生的葡萄糖可以为纤维素合成提供底物。
综上所述,黄昏后毛竹笋竹下部纤维素沉积较多,下部纤维素沉积与淀粉降解和可溶性糖的消耗有关;中部伸长生长快,消耗可溶性糖较多;笋竹上部伸长生长较慢,对碳水化合物的需求量小,上部淀粉主要分解生成、转化为葡萄糖、果糖;竹蔸作为储藏淀粉的重要部位,为笋竹快速生长提供大量的碳水化合物。
[1] CHEN Liqing,CHEUNG L S,FENG Liang,et al.Transport of sugars[J].Annu Rev Biochem,2015,84:865-894.
[2] 马殿荣,孔德秀,刘晓亮,等.杂草稻中胚轴伸长动态及其与籽粒淀粉酶活性和可溶性糖含量的关系[J].中国水稻科学,2014,28(1):97-102. MA Dianrong,KONG Dexiu,LIU Xiaoliang,et al.Mesocotyl elongation of weedy rice and its relationship with grain amylase activities and soluble sugar contents[J].Chin J Rice Sci,2014,28(1):97-102.
[3] ZHANG Huiling,HOU Juan,LIU Jun,et al.Amylase analysis in potato starch degradation during cold storage and sprouting[J].Potato Res.2014,57(1):47-58.
[4] 刘洋,赵香娜,岳美琪,等.甜高粱茎秆不同节间糖分累积与相关酶活性的变化[J].植物遗传资源学报,2010,11(2):162-167. LIU Yang,ZHAO Xiangna,YUE Meiqi,et al.Sugar metabolism and change of related activities of enzymes in internode of sweet sorghum[J].J Plant Genetic Res,2010,11(2):162-167.
[5] GRAF A,SCHLERETH A,STITT M,et al.Circadian control of carbohydrate availability for growth in Arabidopsis plants at night[J].Proc Nat Acad Sci,2010,107(20):9458-9463.
[6] 袁佳丽,温国胜,张明如,等.毛竹快速生长期的水势变化特征[J].浙江农林大学学报,2015,32(5):722-728. YUAN Jiali,WEN Guosheng,ZHANG Mingru,et al.Water potential with Phyllostachys edulis in its fast-growth periods[J].J Zhejiang A&F Univ,2015,32(5):722-728.
[7] 丁雨龙,LIESE W.竹节解剖构造的研究[J].竹子研究汇刊,1995,14(1):24-32. DING Yulong,LIESE W.On the nodal structure of bamboo[J].J Bamboo Res,1995,14(1):24-32.
[8] 刘琳,王玉魁,王星星,等.毛竹出笋后快速生长期茎秆色素含量与反射光谱的相关性[J].生态学报,2013,33(9):2703-2711. LIU Lin,WANG Yukui,WANG Xingxing,et al.Correlation between pigment content and reflectance spectrum of Phyllostachys pubescens stems during its rapid growth stage[J].Acta Ecol Sin,2013,33(9):2703-2711.
[9] 王星星,刘琳,张洁,等.毛竹出笋后快速生长期内茎秆中光合色素和光合酶活性的变化[J].植物生态学报,2012,36(5):456-462. WANG Xingxing,LIU Lin,ZHANG Jie,et al.Changes of photosynthetic pigment and photosynthetic enzyme activity in stems of Phyllostachys pubescens during rapid growth stage after shooting[J].Chin J Plant Ecol,2012,36(5):456-462.
[10] 方楷,杨光耀,杨清培,等.毛竹成竹过程中内源激素动态变化[J].江西农业大学学报,2011,33(6):1107-1111. FANG Kai,YANG Guangyao,YANG Qingpei,et al.Dynamic changes of endogenesis hormone in bamboo formation course(Phyllostachys edulis)[J].Acta Agric Univ Jiangxi,2011,33(6):1107-1111.
[11] CUI Kai,HE Caiyun,ZHANG Jianguo,et al.Temporal and spatial profiling of internode elongation-associated protein expression in rapidly growing culms of bamboo[J].J Proteome Res,2012,11(4):2492-2507.
[12] HE Caiyun,CUI Kai,ZHANG Jianguo,et al.Next-generation sequencing-based mRNA and microRNA expression profiling analysis revealed pathways involved in the rapid growth of developing culms in moso bamboo[J].BMC Plant Biol,2013,13(1):119.doi:10.1186/1471-2229-13-119.
[13] PENG Zhenhua,LU Ying,LI Lubin,et al.The draft genome of the fast-growing non-timber forest species moso bamboo(Phyllostachys heterocycla)[J].Nature Genetics,2013,45(4):456-461.
[14] 刘骏,杨清培,杨光耀,等.厚壁毛竹非结构性碳水化合物分配格局[J].江西农业大学学报,2011,33(5):924-928. LIU Jun,YANG Qingpei,YANG Guangyao,et al.Allocation pattern of non-structural carbohydrates of Phyllostachys edulis‘Pachyloen’[J].Acta Agric Univ Jiangxi,2011,33(5):924-928.
[15] 郑进烜,董文渊,陈冲,等.海子坪天然毛竹种群生长规律研究[J].竹子研究汇刊,2008,27(2):32-37. ZHENG Jinxuan,DONG Wenyuan,CHEN Chong,et al.Studies on the growth and rhythm of natural moso bamboo population in Haiziping[J].J Bamboo Res,2008,27(2):32-37.
[16] TIFFANY T O,JANSEN J M,BURTIS C A,et al.Enzymatic kinetic rate and end-point analyses of substrate,by use of a GeMSAEC fast analyzer[J].Clin Chem,1972,18(8):829-840.
[17] BERNFELD P.Amylases,α and β[J].Meth Enzymol,1955,1:149-158.
[18] CHANTUMA P,LACOINTE A,KASEMSAP P,et al.Carbohydrate storage in wood and bark of rubber trees submitted to different level of C demand induced by latex tapping[J].Tree Physiol,2009,29(8):1021-1031.
[19] MYERS J A,KITAJIMA K.Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest[J].J Ecol,2007,95(2):383-395.
[20] LASTDRAGER J,HANSON J,SMEEKENS S.Sugar signals and the control of plant growth and development[J].J Exp Bot,2014,65(3):799-807.
[21] WANG Yan,YU Bingjie,ZHAO Jinping,et al.Autophagy contributes to leaf starch degradation[J].Plant Cell, 2013,25(4):1383-1399.
[22] STITT M,ZEEMAN S C.Starch turnover:pathways,regulation and role in growth[J].Curr Opin Plant Biol,2012,15(3):282-292.
[23] MATSUDA R,OZAWA N,FUJIWARA K.Leaf photosynthesis,plant growth,and carbohydrate accumulation of tomato under different photoperiods and diurnal temperature differences[J].Sci Hortic,2014,170(3):150-158.
[24] MARTINS M C,HEJAZI M,FETTKE J,et al.Feedback inhibition of starch degradation in Arabidopsis leaves mediated by trehalose 6-phosphate[J].Plant Physiol,2013,163(3):1142-1163.
[25] LIU D D,CHAO W M,TURGEON R.Transport of sucrose,not hexose,in the phloem[J].J Exp Bot,2012,63(11): 4315-4320.
[26] 张懿,张大兵,刘曼.植物体内糖分子的长距离运输及其分子机制[J].植物学报,2015,50(1):107-121. ZHANG Yi,ZHANG Dabing,LIU Man.The molecular mechanism of long-distance sugar transport in plants[J]. Chin Bull Bot,2015,50(1):107-121.
[27] 张春玲.毛竹笋—竹生长发育过程系统分析与生长素相关基因研究[D].北京:中国林业科学研究院,2014. ZHANG Chunling.The Comprehensive Analysis of Shoot-Culm and Study of Auxin-related Genes of Phyllostachys edulis[D].Beijing:Chinese Academy of Forestry,2014.
[28] 董丽娜.毛竹秆茎高生长的发育解剖研究[D].南京:南京林业大学,2007. DONG Lina.Studies on Developmental Anatomy of Elongated Growth about Bamboo Culms[D].Nanijing:Nanjing Forestry University,2007.
[29] 柴静,张会,姚丽丽,等.蔗糖合酶在植物生长发育中的作用研究[J].生命科学,2012,24(1):81-88. CHAI Jing,ZHANG Hui,YAO Lili,et al.The function of sucrose synthase in plant growth and development[J]. Chin Bull Life Sci,2012,24(1):81-88.
[30] 刘波.毛竹发育过程中细胞壁形成的研究[D].北京:中国林业科学研究院,2008. LIU Bo.Formation of Cell Wall in Developmental Culms of Phyllostachys pubescens[D].Beijing:Chinese Academy of Forestry,2008.
Spatial and temporal change of carbohydrates during rapid growth processes of Phyllostachys edulis
CHENG Luyun,WEN Xing,MA Dandan,LI Dandan,XU Xinlu,GAO Yan,ZHANG Rumin
(School of Forestry and Biotechnology,Zhejiang A&F University,Lin’an 311300,Zhejiang,China)
To discuss the change of carbohydrates of Phyllostachys edulis in the process of rapid growth,kits and 3,5-dinitrosalicylic acid colorimetry were used to measure the contents of starch,sucrose,glucose,fructose and cellulose in the bamboo stump,the lower,middle and upper of Ph.edulis bamboo shoot at dusk(0 h),4 h and 8 h after dusk,respectively.Results showed that the carbohydrates mass fractions among different parts in Ph.edulis in the progress of rapid growth existed differences and had different degree of change over time.The starch mass fractions,decreased significantly in all the parts of bamboo stump 8 h after dusk,dropped by 28.7%and 61.5%in the bamboo stump and the middle,respectively;the mass fractions of sucrose,glucose and fructose in the bamboo stump dropped by 29.4%,39.2%and 21.4%,respectively;in the middle of bamboo shoot,sucrose,glucose and fructose mass fractions dropped by 36.9%,54.3%and 26.6%;4 h after dusk cellulose mass fraction in the lower of bamboo stump increased by 25.5%.There were significant positive correlations in mass fractions between the starch with any of sucrose,glucose or fructose in the bamboo stump,the lower and middle of Ph.edulis bamboo shoot,and there were significant negative correlations in mass fractions between starch with any of glucose or fructose in the upper.There were significant negative correlations in mass fractions between cellulose with any of starch,sucrose,glucose or fructose in the lower.After dark,cellulose in the lower of bamboo shoots had more deposition than the other parts,the elongation in the middle was fast,andit consumed more soluble sugar.The upper growth was slowly,and consumed little soluble sugar.Bamboo stump as an important part reserving starch,provided plenty of carbohydrates for rapid growth of bamboo shoots.[Ch,5 fig.4 tab.30 ref.]
plant physiology;Phyllostachys edulis;carbohydrates;rapid growth;spatial and temporal disparities;correlation
S718.43;Q945.1
A
2095-0756(2017)02-0261-07
10.11833/j.issn.2095-0756.2017.02.009
2016-03-24;
2016-05-09
国家自然科学基金资助项目(31570686,30972397,31270497);浙江省省院合作项目(2014SY16)
程路芸,从事植物生理学研究。E-mail:zgrcly@163.com。通信作者:马丹丹,讲师,博士,从事植物生理生态学等研究。E-mail:madandan@gmail.com
猜你喜欢
中国药学药品知识仓库(2022年1期)2022-03-23
健康博览(2021年10期)2021-12-10
少儿科技(2021年8期)2021-01-02
意林·少年版(2020年13期)2020-08-02
东坡赤壁诗词(2020年2期)2020-06-04
婚姻与家庭·性情读本(2019年11期)2019-11-18
食品界(2019年8期)2019-10-07
保健与生活(2019年7期)2019-07-31
阅读与作文(小学高年级版)(2019年3期)2019-04-20
中国药业(2019年5期)2019-03-01
