
侧金盏大小孢子发生及雌雄配子体发育1)
2017-08-01 00:06孙颖王蕾杨雪何淼
东北林业大学学报 2017年7期
孙颖 王蕾 杨雪 何淼
(东北林业大学,哈尔滨,150040)
侧金盏大小孢子发生及雌雄配子体发育1)
孙颖 王蕾 杨雪 何淼
(东北林业大学,哈尔滨,150040)
用石蜡切片技术对早春开花植物侧金盏(AdonisamurensisRegel et Radde.)大、小孢子的发生及雌、雄配子体的发育进行了研究。结果表明: 侧金盏花药壁由外向内分为表皮、药室内壁、2层中层和绒毡层,其发育方式为典型的双子叶型;绒毡层类型为腺质绒毡层。 小孢子母细胞减数分裂过程中的胞质分裂方式为同时型,四分体排列方式为左右对称形或正四面体形。 成熟花粉粒大多数为二细胞型,少数为三细胞型,具有三条萌发沟。 子房为单心皮,单室,横生型胚珠,厚珠心,双珠被。 大孢子母细胞由造孢细胞发育而来,多数情况下合点端第一个大孢子分化为功能大孢子。成熟胚囊为8细胞8核的蓼型胚囊。
侧金盏;大小孢子发生;雌雄配子体发育
AdonisamurensisRegel et Radde; Mega-and microsporogenesis; Female and male gametophyte
侧金盏属于毛茛科侧金盏属(AdonisL.),别名顶冰花、福寿草或冰凉花,是东北地区一种常见多年生草本植物。主要生长在东北山区的阔叶林下或山坡草地中,喜欢富含腐殖质和土层深厚的土壤环境[1-2]。侧金盏花色明亮艳丽、花形美观可爱,开花时间极早,植株抗寒性极强,是一种不可多得的野生花卉资源。
目前,关于侧金盏药用成分[3]、生物学[4-6]、分类学[7-9]的研究报道较多,而有关侧金盏的胚胎学研究却鲜有报道。侧金盏为代表性的早春类短命植物,本试验对侧金盏大、小孢子发生及雌、雄配子体的发育过程进行观察研究,能为此类植物的生殖生物学、遗传育种等提供科学依据。
1 材料与方法
试验材料为采自东北林业大学园林学院苗圃的侧金盏。待侧金盏植株地上部分的茎叶逐渐凋亡,地下部分的根状茎的顶端有白色的小芽冒出的时候,在2014年6月初至10月末,每3~5 d取材一次,经FAA固定液固定,对其进行抽气处理,使材料整体下沉,然后转至冰箱4 ℃冷藏保存。爱氏苏木精整体染色12~15 d,蓝化后,常规石蜡切片法制片,切片厚度8~12 μm,在LEICA DM2500显微镜下观察并照相。
2 结果与分析
2.1 小孢子发生和雄配子体的发育
2.1.1 花药壁的发育
侧金盏的花为两性花,每朵花具70~120枚雄蕊,花药4室(图1A、B)。幼小花药外侧为一层原表皮,内侧为一团分生组织(图1C),后其4个角隅突起,成为近四边形(图1D)。孢原细胞在原表皮下的第一层细胞中形成,经平周分裂(图1E),形成初生壁细胞和初生造胞细胞(图1F)。初生壁细胞经平周分裂和垂周分裂,形成2层次生壁细胞(图1G)。近表皮的次生壁细胞经平周分裂,形成了药室内壁和中层,内层直接分化为绒毡层。小孢子母细胞时期,花药壁已基本分化完全,由外到内依次为表皮、药室内壁、2层中层、绒毡层(图1H)。故侧金盏花药壁的发育方式为典型的双子叶型。
表皮细胞在二细胞花粉期趋于解体,外壁不规则突起,具明显角质层结构(图1K);药室内壁细胞在小孢子母细胞减数分裂期开始径向延长并液泡化(图1I),后细胞体积不断增大(图1J),二细胞花粉期发生多数斜纵向条纹状的“U”形纤维化加厚(图1K);中层为短命结构,小孢子母细胞减数分裂期,中层细胞被药室内壁细胞挤压(图1I),小孢子单核靠边期,中层趋于解体(图1J),二细胞花粉期已完全退化,只留下一些痕迹(图1K);绒毡层细胞在小孢子母细胞减数分裂期开始时逐渐解体(图1I)。
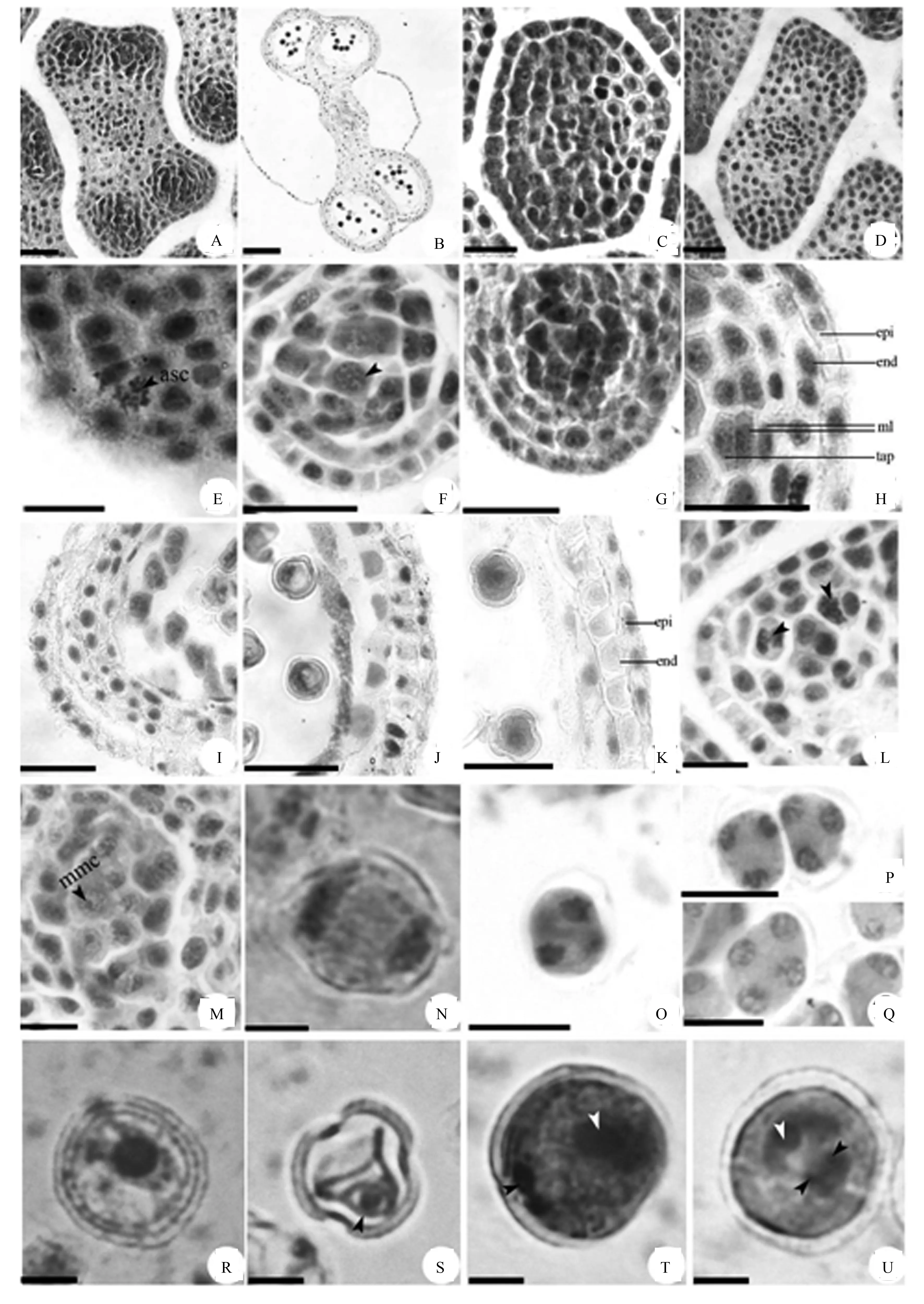
asc.孢原细胞;end.药室内壁;epi.表皮;ml.中层;mmc.小孢子母细胞;tap.绒毡层;A.花药四室;B.成熟花药;C.幼小花药;D.四边形花药;E.孢原细胞平周分裂;F.孢原细胞发育成初生壁细胞和初生造孢细胞;G.初生壁细胞形成两层次生壁细胞;H.5层花药壁;I.小孢子母细胞减数分裂期的花药壁;J.小孢子单核靠边期的花药壁;K.二细胞花粉期的花药壁;L.初生造孢细胞有丝分裂形成次生造孢细胞;M.小孢子母细胞;N.减数第一次分裂后期;O.减数第二次分裂后期;P.正四面体型四分体;Q.左右对称型四分体;R.释放到药室中的小孢子;S.单核靠边期小孢子;T.新形成的二细胞花粉;U.三细胞花粉(A、C、D、F-K.Bar=20 μm;B.Bar=80 μm;E、L、M、O、Q.Bar=10 μm;N、R-U.Bar=4 μm)。
图1 花药壁发育和小孢子的发生及雄配子体发育
2.1.2 小孢子发生
花药壁发育的同时,初生造孢细胞经数次的有丝分裂,形成次生造孢细胞(图1L),次生造孢细胞进一步发育成小孢子母细胞(图1M)。小孢子母细胞进入减数第一次分裂时期(图1N),此次分裂没有形成细胞壁结构,并且细胞质也不发生分裂。
随后两个子细胞进行减数第二次分裂(图1O),此次分裂伴随着细胞质的分裂,形成四分体。侧金盏四分体的排列方式大多数为正四面体型(图1P),也有少数为左右对称型(图1Q),胞质分裂为同时型。在减数分裂过程中,小孢子母细胞的细胞壁内逐渐积累形成胼胝质,后来形成的四分体被胼胝质壁所包围,而且同一个四分体的各个小孢子也都被胼胝质包围。四分体胼胝质壁随后被溶解,四分体分开,形成单核小孢子(图1R),至此小孢子形成。
观察发现,在同一朵花内,不同花药之间和同一花药内不同药室中的小孢子母细胞的减数分裂表现为明显的不同步,一般相差2~4个时期,但在同一个药室内,小孢子母细胞的减数分裂基本保持一致。
2.1.3 雄配子体发育
从四分体中释放出的小孢子,形状不规则,细胞壁微微的收缩,为收缩期小孢子,后细胞质液泡化,发育为单核居中期小孢子(图1R)。小孢子体积逐渐增大,椭球形,细胞壁加厚,中央大液泡形成且体积逐渐扩大,将细胞核逐渐挤压靠近细胞壁的一侧,同时形成萌发沟,可见3个萌发孔,成为单核靠边期小孢子(图1S)。小孢子的细胞核发生有丝分裂并生成2个子核,随后发生不均等的胞质分裂,形成两个细胞,大的为营养细胞,小的靠近花粉壁呈透镜状的为生殖细胞(图1T)。随着雄配子体的发育,两细胞之间的细胞壁消失,生殖细胞经有丝分裂形成2个精子。入冬前侧金盏花粉发育成熟,大部分成熟花粉为二细胞型,少数为三细胞花粉。三细胞花粉外观圆球形,3个萌发沟,营养细胞核位于雄配子体的一侧,和两个精细胞排列成三角形(图1U)。在散粉前,花粉基本已是三细胞型。
2.2 大孢子的发生和雌配子体发育
2.2.1 胚珠的发育
侧金盏的子房聚合在一个隆起的花托上,单心皮,单室,厚珠心,横生型胚珠。子房壁的珠心组织发育成胚珠原基(图2A)。珠孔端珠心表皮下的一个薄壁细胞特化成为孢原细胞,孢原细胞经平周分裂(图2C)生成1个周缘细胞和1个造孢细胞,周缘细胞经1次平周分裂和数次垂周分裂,使大孢子母细胞位于珠心顶端4层细胞之下,而造孢细胞进一步发育后行使大孢子母细胞的功能(图2D)。所以,侧金盏的珠心属于厚珠心。同时,胚珠原基的基部发生多次有丝分裂逐渐衍生出内外2层珠被组织,外层组织由6~7层细胞组成,称为外珠被,内层组织由3~4层细胞组成,称为内珠被,内、外珠被共同形成珠孔结构。胚珠的基部与子房相连的部分发育为珠柄,珠柄进入胚珠处为合点,在胚珠的发育过程中,由于珠柄和其他部位的生长速度不均等,使胚珠的珠心和珠被的纵轴与珠柄垂直,形成横生型胚珠(图2B)。
2.2.2 大孢子发生
大孢子母细胞体积增大,随后开始进行减数分裂,经历了较长时间的前期Ⅰ,后进入中期Ⅰ、后期Ⅰ、末期Ⅰ(图2E),形成二分体(图2F)。与小孢子母细胞减数分裂过程的不同点是,在大孢子母细胞减数分裂的末期Ⅰ中细胞核生成的同时伴随着胞质分裂。从其雌配子发育过程推测,一般保留位于合点端的第一个大孢子,并最终发育成为功能大孢子进而发育为单核胚囊,而位于珠孔端的3个大孢子逐渐退化。
2.2.3 雌配子体发育
随着功能大孢子的体积不断增大,出现液泡,功能大孢子发育为单核胚囊(图2G)。大孢子发育到一定程度连续进行3次有丝分裂,第1次分裂,形成2个子核的分别向珠孔端和合点端方向移动,形成二核胚囊(图2H)。随后每个核又进行1次分裂,分别产生2个子核,形成四核胚囊(图2J、K)。但在此发育时期也有异常情况发生,由于核分裂不同步,珠孔端的核先分裂,可能会看到一端一核、一端二核的三核胚囊(图2I)。继而四核胚囊的每个核又各自进行1次有丝分裂,形成了八核胚囊(图2L),8个核处于共同的细胞质中。八核胚囊停留时间较短,随即进入细胞化阶段,各核间生成细胞壁构成细胞。卵细胞是胚囊的核心,最初形成的卵细胞,细胞核明显,液泡小(图2M)。成熟的卵细胞具有明显的极性,通常位于近合点端(图2N),但有时也会处于近珠孔端(图2O)。2个助细胞并列分布于近珠孔端,且与卵细胞组成近“品”字形的卵器。助细胞极性明显且与卵细胞相反(图2P、Q)。助细胞的存活时间极短,其中一个在花粉管进入胚囊时便已被破坏并退化,另一个在双受精过程完成之后不久退化。两个极核在受精之前没有发生融合(图2Q)其排列方式为并列分布(图2R)或上下分布(图2S)。3个反足细胞成"品"字形紧密的排列于胚囊合点端(图2T)。
fu.珠柄;ii.内珠被;mec.孢原细胞;nu.珠心;oi.外珠被;vb.维管束;ant.反足细胞;ec.卵细胞;pn.极核;sy.助细胞;A.胚珠原基;B.胚珠纵切面;C.孢原细胞有丝分裂;D.大孢子母细胞;E.大孢子母细胞减数分裂;F.二分体;G.单核胚囊;H.二核胚囊;I.珠孔端细胞先分裂;J.四核胚囊中位于合点端的2个核;K.四核胚囊中位于珠孔端的两个核;L.八核胚囊(示七核);M.刚分化的卵细胞;N.核位于合点端的卵细胞;O.核位于珠孔端的两个细胞;P.卵器;Q.上下排列的极核和2个助细胞;R.左右排列的极核和卵细胞;S.上下排列的极核和卵细胞;T.3个反足细胞(A、B.Bar=20 μm;C-T.Bar=10 μm)。
图2 胚珠发育和大孢子发生与雌配子发育
3 结论与讨论
本试验观察到侧金盏的花药为4室,花药壁由外向内分为表皮、药室内壁、2层中层和绒毡层5层,绒毡层类型为腺质绒毡层。宋萍[10]在毛茛科川陕金莲花(Trolliusbuddae)和驴蹄草(Calthapalustris)中观察到的绒毡层也为腺质绒毡层。有相关研究认为,绒毡层细胞通过自溶为迅速发育的花粉提供营养物质[11],在侧金盏的花粉粒发育过程中,小孢子母细胞减数分裂期起,绒毡层开始解体,至花药成熟时完全退化。由此可见,侧金盏绒毡层活动状态与小孢子、雄配子发育对营养物质的要求相一致。根据Davis[12]对花药壁发育方式的划分,侧金盏花药壁的发育方式为典型的双子叶型。
毛茛科胞质分裂在小孢子母细胞减数分裂中为同时型,四分体为四面体形、左右对称形或交叉形[13]。侧金盏小孢子母细胞减数分裂为同时型,四分体排列方式有左右对称形或正四面体形两种类型。而刘萍[14]对铁筷子(Helleborusthibetanus)和宋萍[10]对驴蹄草(C.palustris)和川陕金莲花(T.buddae)的研究结果均显示四分体排列方式只有四面体形一种。
侧金盏成熟花粉多数为二细胞型,少数为三细胞型。二细胞型花粉在毛茛科是很普遍的,Tamura[13]报道在毛茛科21属中两细胞花粉较常见,极少观察到三细胞花粉。
对侧金盏观察发现,在同一朵花内,不同花药之间和同一花药内不同药室中的小孢子母细胞的减数分裂表现为明显的不同步,一般相差2~4个时期,但在同一个药室内,小孢子母细胞的减数分裂基本保持一致。Heslop-Harrison[15]认为,胞间连丝的存在使同一药室内的小孢子母细胞减数分裂同步化,但在试验过程中并未发现侧金盏存在胞间连丝结构。
侧金盏子房为单心皮,单室,横生型胚珠,厚珠心,双珠被。侧金盏的由珠心表皮下的薄壁细胞特化而来的1个孢原细胞并不直接分化出大孢子母细胞,而是先经过平周分裂形成1个周缘细胞和1个造孢细胞,造孢细胞进一步发育行使大孢子母细胞的功能,这与Tamura[13]对毛茛科植物大孢子母细胞起源的描述一致,刘萍[14]在对铁筷子(H.thibetanus)的研究中发现了同样的现象。但现有的文献资料中也记载了特殊情况,毛茛科驴蹄草属的驴蹄草(C.palustris)的孢原细胞不发生分裂,直接起大孢子母细胞的作用[10]。侧金盏的厚珠心来源于珠心表皮细胞的分裂和周缘细胞的共同分裂,而王自芬[16]在对毛茛科驴蹄草(C.palustris)胚珠的厚珠心来源于珠心表皮的分裂。
侧金盏的功能大孢子经单核胚囊、二核胚囊、四核胚囊发育成为8核8细胞胚囊;但在四核胚囊形成期,由于核分裂不同步,偶见一端有一核,一端有二核的三核胚囊。观察还发现侧金盏的助细胞存活时间极为短暂,其中一个在花粉管进入胚囊时便已退化,另一个在受精完成后不久退化,这种现象在棉花(Gossypiumhirsutum)、玉米(Zeamays)、大麦(Hordeumvulgare)、针茅(Stipacapillata)中广泛存在[11]。
侧金盏的果实为聚合瘦果,每个果实上有50~60枚种子。有研究发现,无论在野生还是引种状态下,侧金盏种子都存在一定的“空籽”,即种胚败育[4]。通过对大量不同时期的花芽材料进行切片观察,并未发现侧金盏大小孢子发生和雌雄配子体发育过程中存在发育不良的情况,可见侧金盏的“空籽”形成原因与后期的传粉受精过程有密切的关系,需要进一步进行试验研究。
[1] 王文采.中国植物志[M].北京:科学出版社,1980:246-255.
[2] 王文采.侧金盏花属修订(一)[J].植物研究,1994,14(1):1-31.
[3] 尹蕾,张冤,田海妍,等.福寿草的化学成分研究[J].中草药,2014,45(23):3361-3366.
[4] 陈士惠.侧金盏花繁殖生物学研究[D].哈尔滨:东北林业大学,2013.
[5] 曲艳婷,唐焕伟,陈菲.低温胁迫对侧金盏部分生理生化指标的影响[J].国土与自然资源研究,2009(3):82-83.
[6] 李欢,肖筝,王本全,等.侧金盏花组织培养及无性系建立的研究[J].特产研究,2010,32(1):22-24.
[7] TAMURA M. A new classification of the family Ranuneulaeeae 2 (in Japanese)[J]. Acta Phytotaxonomica et Geobotanica,1991,42(2):177-187.
[8] 王文采.侧金盏花属修订(二)[J].植物研究,1994,14(2):105-138.
[9] HEYN C C, PAZY B. The annual species ofAdonis(Ranunculaceac)-a polyploid complex[J]. Plant Systematics and Evolution,1989,168(3):181-193.
[10] 宋萍.金莲花属和驴蹄草属(毛茛科)的比较形态学研究[D].西安:陕西师范大学,2007.
[11] 胡适宜.被子植物生殖生物学[M].北京:高等教育出版社,2005:33-43.
[12] DAVIS G L. Systematic embryology of the angiosperms[M]. New York: John Wiley & Sons, Lnc,1996.
[13] TAMURA M. Ranunculaceae[M]//KUBITZKI K, ROHWER J G, BITTRICH V. The families and genera of vascular plants: vol.2. flowering plants dicotyledons: magnoliid, hamameliid and caryophyllid families. Berlin: Springer-Verlag Berlin Heidelberg,1993:567-583.
[14] 刘萍.铁筷子毛茛科的形态、结构、发育及系统位置的研究[D].西安:陕西师范大学,2006.
[15] HESLOP-HARRISON J. Cytoplasmic connexions between angiosperm meiocytes[J]. Annals of Botany,1966,30(2):221-222.
[16] 王自芬.毛茛科植物胚珠形态、结构、发育及其系统学意义[D].西安:陕西师范大学,2008.
孙颖,女,1979年10月生,东北林业大学园林学院,副教授。E-mail:littlesuning@126.com。
何淼,东北林业大学园林学院,副教授。E-mail:hemiao_xu@126.com。
2017年2月8日。
S681.7;Q944.4
Megasporogenesis, Microsporogenesis and Development of Male and Female Gametophytes ofAdonisamurensisRegel et Radde//Sun Ying, Wang Lei, Yang Xue, He Miao(Northeast Forestry University, P. R. China)//Journal of Northeast Forestry University,2017,45(7):61-65.
1)中央高校基本科研业务费专项资金项目(2572017CA13)。
责任编辑:任 俐。
We studied the megasporogenesis and development of female and male megasporogensis ofAdonisamurensisRegel et Radde, which blooms in early spring, by paraffin wax. The development of the anther wall belongs to a dicotyledonous type. The wall of anther consists of epidermis, endothecium, two middle layers and tapetum from out to inside, and tapetum cell is of glandular tepetum. Successive cytokinesis in microspore mother cell accompanies meiosis to produce tetrahedral tetrad as well as medianly zygomorphic tetrad. Mature pollen grains, with three germ pores, most are 2-cell type, while a few are 3-cell type. The ovary ofA.amurensishas 1 carpel, 1 loculuses with amphitropous ovule which is crassinucellate and bitegmic. Megasporocyte developes from sporogenous cell. In most cases, the first chalazal megaspore differentiates into functional megaspore. The mature embryo sacs has 8 cells with 8 nucleis, and belongs to polygonum type embryo-sac.
猜你喜欢
食品研究与开发(2021年19期)2021-11-17
雪豆月读·低年级(2021年1期)2021-09-10
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
河北画报(2020年19期)2020-06-16
大理文化(2020年3期)2020-06-11
广西植物(2016年10期)2016-11-11
中国医药指南(2014年21期)2014-05-18
浙江农业科学(2013年6期)2013-11-08
中国蔬菜(2013年8期)2013-01-28
中国实用医药(2012年7期)2012-11-21
