
安吉毛竹林非生长季CO2通量与其光合响应关系探究
2017-10-13 20:46舒海燕江洪陈晓峰孙文文马锦丽张敏霞
生态科学 2017年2期
舒海燕, 江洪,,3*, 陈晓峰, 孙文文, 马锦丽, 张敏霞
安吉毛竹林非生长季CO2通量与其光合响应关系探究
舒海燕1, 江洪1,2,3*, 陈晓峰2, 孙文文2, 马锦丽2, 张敏霞2
1. 西南大学三峡库区生态环境教育部重点实验室, 重庆400715 2. 浙江农林大学林业与生物技术学院, 浙江临安311300 3. 南京大学国际地球系统科学研究所, 江苏南京210046
利用涡度相关法和生理生态法(光合)对安吉毛竹林生态系统非生长季(2014.10—2015.4, 无2月)的CO2通量进行测定, 结果表明, 两种方法具有良好的响应关系, 基于涡度的生态系统净交换量(和基于光合的生态系统净初级生产力(日变化均呈单峰型曲线, 受光合有效辐射()的直接影响, 在午间12:00前后有最大值, 且两者均表现为秋季最大(10月最高), 春季次之, 冬季最低。就各月而言, 基于光合测定的日最大值和月总量均低于涡度相关技术白天的测定值, 但均占到后者的40%和65%以上, 在毛竹光合能力比较强的10、11和4月, 所占比例更大。表明在白天, 安吉毛竹林生态系统的主要取决于毛竹的光合作用。此外, 观测期间, 毛竹林生态系统在的净初级生产力总量为1365.59 gCO2·m–2, 吸收的碳约372.44 gC·m–2, 整体表现为较强的碳汇。
毛竹林生态系统; 非生长季; 涡度相关技术; 光合作用; 净初级生产力
1 前言
随着社会的进步、经济水平的提升以及工业产业迅速发展, 越来越多的环境问题日益突出, 尤其是全球气候变暖吸引着全社会人类的目光, 不止对我国的经济、社会及生态环境造成了重大的影响, 也是世界社会经济可持续发展所面临的严峻挑战, 同时严重威胁着人类的生活及安全, 正如所指: “温室效应”是工业革命以来影响范围最广的生态环境问题之一[1]。大气中CO2等温室气体浓度不断升高是引起气候变暖的重要原因, 对全球变暖的贡献率达60%[2]。森林生态系统作为陆地生态系统的主体, 覆盖面积约1/3左右[3], 因其高大的植被特性和复杂的结构, 使之具有更高的碳密度贮量和最高的生产力以及生物量积累, 使得森林生态系统成为吸收和储存CO2的有效碳库, 具有很强的碳汇能力[4]。我国自20世纪70年代以来, 人工林生态系统面积逐渐增加且居世界首位, 已成为我国森林生态系统的重要组成部分[5–8], 所以量化人工林生态系统的固碳对研究陆地生态系统碳循环以及全球碳的源/汇具有相当重要的意义。毛竹林作为典型的人工林生态系统, 具有较高的固碳能力[9]。毛竹还具有生长快、产量高、用途广、效益大和持续更新等特点, 在竹类资源中占有最大比例, 现成为我国分布最多、面积最大、利用领域最广的经济竹种[10–11]。此外, 由于毛竹林根鞭发达, 更新频繁, 对区域防风固沙、固碳排氧和调节气候等方面发挥着巨大的生态保护作用, 具有较高的生态效益[12–13]。但是, 在非生长季, 主要由毛竹的光合作用转向植被、凋落物和土壤微生物等的呼吸作用, 使得森林碳吸收降低, 甚至表现为碳释放, 很大程度上影响了毛竹林生态系统全年的碳源/汇强度。因此, 研究毛竹林非生长季的CO2通量显得尤为重要。
关于生态系统CO2通量研究的方法较多, 主要的是以微气象学为基础的涡度协方差法和生理生态学方法。其中最为直接的可连续测定的方法是涡度协方差法, 1986年被首次用于森林生态系统CO2通量的研究中[14], 发展至今, 尽管还存在着一些不足之处, 该方法仍然作为现今碳通量研究的一个标准方法[15–17]; 而生理生态学方法主要是使用测定室对森林生态系统不同组分分别进行测定并通过数学方法进行总量估计的方法, 该法能够细化生态系统的碳平衡于不同组分之间[18]。关于涡度协方差法, 运用较广, 如草原[19]、荒漠[20]、稻田[21]、针叶林[22–25]、针阔混交林[26–29]、阔叶混交林[30–32]、热带季雨林[33]、雷竹林[34]、毛竹林[9][13]等等, 但是将生理生态法与微气象法同时结合运用的研究较少。本研究将以安吉毛竹林为研究对象, 同时结合两种研究方法研究其非生长季(2014.10—2015.04)的CO2通量, 探究毛竹林非生长季CO2通量与其光合的响应关系, 明确毛竹光合在生态系统净交换量(net ecosystem exchange,), 乃至净初级生产力(net ecosystem productivity,)中的贡献, 以便揭示毛竹林生态系统的生理生态学基础, 从而解释安吉毛竹林生态系统的碳源/汇能力。
2 研究地区与研究方法
2.1 研究地区概况
研究区位于浙江省湖州市安吉县山川乡(30°28′34.5″N, 119°40′25.7″E), 为典型的亚热带季风气候, 全年气候温和、雨水充沛、光照充足、四季分明。历史年均气温14.6 ℃, 年日照1980—2050 h, 年无霜期203—226 d, 年降雨日140—170 d, 降水量达1100—1900 mm。降水量由北至南递增, 6月和8月雨量最多, 12月雨量最少, 具明显的双峰型降雨的特点, 各年份梅雨量的实际差异大, 是造成旱涝的原因[13]。该地区森林覆盖率达到71%, 拥有山林13.2万hm2, 其中竹林面积6.33万hm2, 毛竹林面积为4.99万hm2, 占森林总面积37.8%, 竹林总面积的78.8%。观测站点海拔高度380 m, 站点下垫面坡度为2.5°—14.0°, 以观测塔为中心1000 m范围内主要森林类型为毛竹林, 土壤类型主要为黄壤。试验区内的毛竹林为粗放型人工纯林, 竹林密度为4500株·hm–2, 胸径12—18 cm, 高度16—20 m, 冠幅3.0 m × 2.5 m, 盖度90%, 郁闭度0.9, 毛竹节间短, 壁厚, 一般3—4月为发笋期, 5月毛竹开始抽叶, 6月份大量展开, 7—9月份为主要生长季, 生长能力较强。
2.2 研究方法
2.2.1 CO2通量的测定
本研究采用了开路式涡度相关系统对安吉毛竹林生态系统的CO2通量进行连续测定。观测林地微气象观测塔高40 m, 开路涡度相关系统(OPEC)探头的安装高度为距地面38 m, 由三维超声风速仪(CAST3, Campbell Inc., USA)和开路CO2/H2O分析仪(Li-7500, Li-Cor Inc., USA)组成, 前者用于测定风速和温度脉动, 后者可测定CO2/H2O密度脉动, 并在线计算并存储30 min CO2通量(), 其原始采样频率10 Hz, 通过数据采集器(CR1000, Campbell Inc., USA)存储数据。此外, 常规气象观测系统包括7层风速(010C, metone, USA)、7层大气温度和湿度(HMP45C, Vaisala, Helsinki, Finland), 安装高度分别为1 m、7 m、11 m、17 m 、23 m、30 m和38 m; 光合有效辐射和净辐射传感器安装高度38 m, 用于采集光合有效辐射、上行/下行的长波/短波辐射、净辐射数据。常规气象观测系统数据采样频率为5 Hz, 通过数据采集器(CR1000, Campbell Inc., USA)每30 min自动记录平均风速、湿度、温度、气压、净辐射等常规气象信息。本研究中空气温度、空气湿度与饱和水汽压差选用23 m高冠层气象数据。仪器设备详情可参考孙成[9]、刘玉莉[12]和杨爽[13]相关研究报道。
关于植被与大气间CO2通量的计算, 用植被与大气间净生态系统CO2交换量(, mg·m–2·s–1)表示, 定义为:
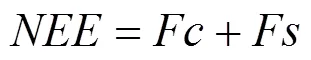
(2)
式中:c(mg·m–2·s–1)为CO2湍流通量, 通过利用三维超声风速仪测定三维风速, 以及红外线CO2/H2O气体分析仪测定CO2密度脉动, 再采用EdiRe ()软件进行协方差运算, 并进行二次坐标轴旋转、WPL()修正等, 最终将利用数据采集器记录并储存的10Hz的原始数据转化为30 min数据而得到。具体数据处理方法详见文献[13]。s(mg·m–2·s–1)为观测高度以下的碳储存通量;为观测高度(m);c为高度h处前后两次相邻时间测定的CO2浓度差值(mg·m–3);t为前后两次测定的时间间隔(s)。当CO2从生态系统排放到大气时, 定义符号为正, 当CO2从大气进入到生态系统时, 定义为负。
2.2.2 叶片光响应曲线的测定
利用便携式光合测定仪LI-6400XT(Li-cor, USA)对毛竹林冠层叶片的光响应曲线进行测定。于2014年10月—2015年4月间每月选定一个晴天, 就毛竹林冠层中上部的叶片进行光合测定, 测量时, 人在铁塔上, 选用LED 2 cm×3 cm叶室。每次至少测定5个重复, 最后求取平均值。得到的光响应曲线采用IBM SPSS Statistics 21.0软件通过非直角双曲线公式进行拟合:
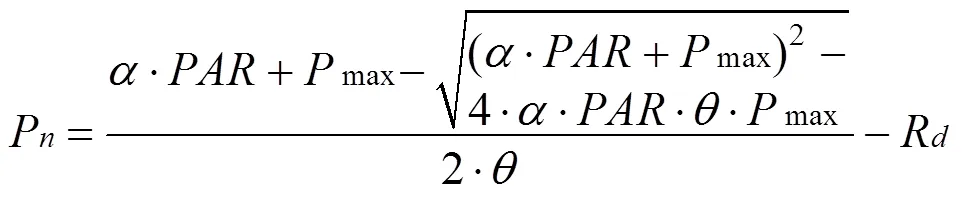
式中:为光合有效辐射(μmol·m–2·s–1),n为叶片在某一值下的净光合速率(μmol·m–2·s–1),max为光饱和时最大净光合速率(μmol·m–2·s–1),d为植物的暗呼吸速率(μmol·m–2·s–1)。为表观量子效率(μmol·μmol–1),为曲线曲率系数, 在0—1之间。用公式(3)拟合出这些参数之后, 再根据通量塔得到的值, 计算出各月白天(≥2 μmol·m–2·s–1)n的平均日变化。
根据公式(1)可以计算出林分的值, 在生理生态学方法中, 与之对应的是净生态系统生产力(, mg·m–2·s–1)[18]。符号的定义与植被/大气间净生态系统CO2交换量()正好相反, 当CO2从大气进入到生态系统时, 定义为正; 当CO2从生态系统排放到大气时, 定义符号为负。但它们绝对值相等, 即。在此,可以粗略地计算为毛竹叶片的净光合速率与当月叶面积指数(leaf index area,)的乘积。即:
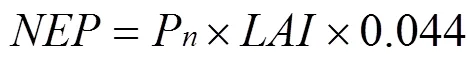
式中0.044为单位换算系数, 即1 μmol·m–2·s–1=0.044 mg·m–2·s–1。这样, 白天n的平均日变化就可以转化为白天的平均日变化, 从而与通量的日变化进行比较。然而, 因为温度和天气等原因, 2015年2月数据空缺。
2.3 数据处理
首先, 植被和大气间净生态系统CO2交换()在白天(≥2 μmol·m–2·s–1)仅仅来源于生态系统植被的光合作用[35]。同植物叶片的光响应曲线一样,对也存在一定的响应关系, 且符合直角双曲线方程, 其函数通常可以用Michaelis-Menten模型表达[36–37]:
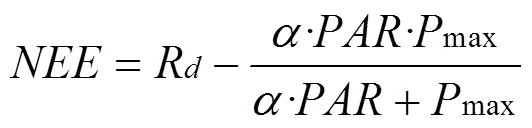
公式(5)中,为白天净生态系统交换量(mg·m–2·s–1), 为表观初始初始光能利用率(表观量子效率, mg·μmol–1),为光合有效辐射(μmol·m–2·s–1),max为光饱和时最大净光合速率(mg·m–2·s–1),d为光照强度趋近于0时植物的呼吸速率(暗呼吸, mg·m–2·s–1)。
其次, 利用IBM SPSS Statistics 21.0软件拟合毛竹叶片的光响应曲线, 进而拟合对的响应曲线, 并得到d、max和等参数。
最后, 采用Excel 2003对数据进行处理, 同时运用统计分析软件IBM SPSS Statistics 21.0对数据进行显著性分析。作图采用Origin 8.6软件。数据以平均数±标准差(Mean ± SD)形式呈现。
3 结果与分析
3.1 毛竹林生态系统气候因子变化特征
试验区属于亚热带季风气候, 全年气候温和、雨水充沛、光照充足、四季分明。就2014年10月—2015年4月毛竹林生态系统的光合有效辐射(photosynthetic active radiation,)、空气温度(air temperature,a, 23m)、空气相对湿度(relative humidity,, 23m)和饱和水汽压差(vapor pressure deficit,, 23m)的动态变化特征进行分析, 如图1所示。
由图1(a)可知, 2014年10月—2015年4月的光合有效辐射()先降低后升高, 但因为2014年11月阴雨天气较多, 光照被削弱,值偏低; 此外, 冬季值相对较低, 1月最低, 平均为181.74± 268.01 μmol·m–2·s–1, 10月和4月相对较高, 平均分别达到311.10±434.98 μmol·m–2·s–1和349.45± 440.77 μmol·m–2·s–1, 入春之后,值继续增大。空气温度()变化趋势呈“U”型曲线(图1(b)), 10月最高, 为17.53±1.18 ℃, 最小值出现在12月, 仅4.23± 1.78 ℃, 入春后,a迅速升高。相对湿度()和饱和水汽压差()变化趋势刚好相反(图1(c, d))。受值的影响,和在12月存在转折, 12月最低, 平均仅47.66%±5.99%, 表明12月份降雨最低, 符合冬季寒冷少雨特征; 而该月的稍微升高, 达到4.59±1.15 kPa。另外, 受和a共同影响,在2月最低, 为2.81±1.003 kPa, 2月以后逐渐上升, 表明2月之后, 毛竹林的水分利用能力(或者蒸腾作用)逐渐增强, 主要因为2015年为毛竹生理大年, 出笋量较大, 水分消耗相应较高。
3.2 毛竹林生态系统CO2通量的变化特征
统计得到2014年10月—2015年4月(不包括2015年2月, 下同)同时刻的数据, 并进行平均处理, 即得到该时间段毛竹林生态系统CO2净交换量()的月平均日变化, 如图2。同时, 图3以2014年10月和2015年1月为例, 比较了随光照强度()的改变而发生的变化。
由图2可知, 2014年10月—2015年4月各月的变化规律性较强, 均呈“U”型曲线变化, 各月CO2交换速率最大值出现在午间12:00前后, 夜间为正值(释放CO2), 白天为负(吸收CO2), 冬季数值较低。各月均值也为负, 表明毛竹林生态系统在非生长季各月均表现为碳吸收, 10月吸收速率最大, 为0.12±0.29 mg·m–2·s–1, 1月吸收速率最低, 仅0.04±0.17 mg·m–2·s–1, 但各月间没有显著差异(>0.05); 表明毛竹林生态系统非生长季总体表现为一定的碳汇作用, 10月的碳汇作用较强, 也证明了毛竹林具有较强的固碳能力[9], 甚至在冬季还能够表现为碳吸收。另外, 因为11月份的值偏低(图1(a)), 限制了植被的光合作用, 使得其数值略低于12月。
图3结合日出和日落显示了与的响应变化, 可知, 每天的值呈现倒“V”变化, 并且与有着良好的负相关关系。综合图2和图3, 可以看出早晨6:00(冬季6:30)之前和傍晚18:00(冬季17:30)以后,值为0, 此时, 毛竹林不能依靠光能进行光合作用吸收CO2, 而呼吸作用仍在进行, 导致继续向外界释放CO2, 使得毛竹林生态系统为正值; 然而, 受低温、黑暗等的影响, 各月该时间段值较低, 维持在0—0.15 mg·m–2·s–1之间变化, 尤其是12—1月更加明显, 其中1月最低, 平均仅0.08 mg·m–2·s–1, 而春秋两季相对较高。说明该生态系统在此时间段(夜间)表现为较弱的碳源作用, 且有明显的季节变化, 即春秋季高于冬季。早晨6:00(或6:30)以后, 伴随着日出,值逐渐增大, 毛竹林开始通过光合作用从外界吸收CO2, 与此同时, 毛竹林生态系统慢慢从正值向负值转变, 表明该生态系统由碳源逐渐变为碳汇; 午间12:00前后,达到峰值, 同时, CO2的吸收速率也达到一天的最大值, 10月最高, 达到0.73 mg·m–2·s–1, 而1月仅0.38 mg·m–2·s–1, 表明此时毛竹林的光合作用为当天最强, 吸收的CO2最多; 再之后, 随着时间的推移, 太阳西下,值逐渐减小, 毛竹光合作用逐渐减弱,也相应降低, 并慢慢在18:00(或17:30)后变为负值, 毛竹林又逐渐变成碳源。分析原因, 生态系统的变化主要是由植物的光合作用和生态系统呼吸引起的, 而白天最主要的是植物的光合作用, 它是植物从大气中吸收固定CO2的主要途径, 而值是植物光合作用的必须条件, 制约着光合作用的进行,值为0时, 植物没有光合作用, 此时为正值; 而增大时, 植物的光合作用增强,逐渐变为负值, 因此说毛竹林生态系统取决于毛竹林的光合作用, 也间接取决于值, 从另一角度表明的变化基本取决于林木光合作用的变化[35]。
3.3 毛竹叶片光响应曲线及的变化特征
毛竹叶片的光合作用强度影响着该生态系统的变化, 而光合作用强度又主要受控于环境光照强度, 光响应曲线(light response curve)可以直接反映其净光合速率与光照强度的关系, 从而使得光合作用和光照强度之间的关系变得易于理解和研究。图4分析比较了2014年10月—2015年4月(不包括2015年2月)毛竹叶片的光响应曲线。
可知, 10月光合作用最高, 平均净光合速率为1.41±0.82 μmol·m–2·s–1, 光补偿点较低(<10 μmol·m–2·s–1),光饱和点较高(>500 μmol·m–2·s–1), 呼吸较低 (0.17 μmol·m–2·s–1), 而1月光合作用最低, 平均仅0.59±0.52 μmol·m–2·s–1。毛竹叶片12—4月暗呼吸偏高, 约0.55 μmol·m–2·s–1; 此外, 各月最大净光合速率(max)由大到小依次为: 10月>11月>4月>3月>12月>1月, 其中最大值为2.15 μmol·m–2·s–1(10月), 约是最小值(1月, 约0.96 μmol·m–2·s–1)的2.2倍。表明冬季和早春时节, 毛竹的光合作用较弱, 尤其冬季最低, 主要是因为此时间段环境温度低, 影响着一些酶的生物活性, 从而限制着植物的生理活动; 此外, 入冬后, 光照时间缩短, 光照强度减弱, 也削弱了植物的光合作用。总之, 对比图2可知毛竹叶片的光合作用强度与毛竹林生态系统有着一致的变化, 净光合速率越大,数值也越高(负向越大), 反之, 也越低。
同时, 运用公式(3)和(4)计算出该非生长季各月白天(≥2 μmol·m–2·s–1)的净初级生产力(), 设定夜间(=0)的为0, 如图5。可见各月白天的均为正值, 并呈现倒“U”型变化, 直接受控于值的改变。因为受到饱和光照的影响, 尽管中午时分光照强烈, 但维持在最大值而没有剧烈起伏变化, 10月最高, 1月最低, 各月依次为0.43、0.36、0.23、0.19、0.25和0.31 mg·m–2·s–1, 均低于涡度相关系统观测结果。主要是因为涡度相关系统观测范围较广, 观测结果较为全面, 而基于光合的测定组分较为狭窄, 不包括除了毛竹叶片以外的树干、枝条等进行光合作用吸收CO2的部分。另外, 受光照、温度、湿度等环境因子的影响, 各月总体表现为秋季最高, 春季居中, 冬季最低, 与的季节变化一致, 表明两者响应关系较好。
3.4 毛竹林生态系统和对的响应
由2.2可知,与有着显著的负相关关系, 而这种关系可以通过Michaelis-Menten方程来描述和体现。就2014年10月—2015年4月(无2月)白天(≥2 μmol·m–2·s–1)的对的响应曲线进行拟合, 结果如图6所示。同时利用SPSS软件对白天和进行拟合, 所得拟合参数结果见表1。
据图6可知, 12月和1月份曲线比较接近,最大数值分别是0.44和0.38 mg·m–2·s–1, 相对应光合有效辐射分别是944.90和734.77 μmol·m–2·s–1; 秋季和春季的数值相对较高, 10、11和4月近似线性变化, 10月有最大值, 达到0.73 mg·m–2·s–1, 对应的光照强度为1244.45 μmol·m–2·s–1, 4月达到0.60 mg·m–2·s–1, 对应值为1203.83 μmol·m–2·s–1。可见冬季的光照强度弱, 日照时间短, 环境温度低, 毛竹林吸收CO2的能力也相应减弱, 随着春季的到来, 光照强度又逐渐增加, 毛竹林吸收CO2的能力也逐渐增强。
采用Michaelis-Menten方程, 非线性回归拟合毛竹林生态系统非生长季不同月份白天对值的响应曲线, 拟合系数2均在0.93以上, 且均达到极显著水平(图6)。2014年10月—2015年4月生态系统暗呼吸速率d较低, 平均值变化为0.004—0.015 mg·m–2·s–1, 11月份偏高(表1), 主要是该月阴雨天较多, 云层较厚, 白天云层吸收热量, 在夜间释放出来, 使得生态系统内部温度稍高, 利于植物的呼吸[38]; 12月之后偏低, 因为环境温度较低, 直接影响着植物的生理活动, 导致其呼吸速率也相应较低, 而随着温度的升高, 毛竹的呼吸速率逐渐增大; 最大净光合作用速率先降低后升高, 而初始光能利用率则先升高后下降, 4月最小, 为0.0005mg·μmol–1Photons, 表明光照强度较弱时, 毛竹的光合作用也较弱, 但是毛竹的初始光能利用率反而较高, 能捕获更多的光能进行光合作用, 从而维持自身的生长。相关研究表明[18], 阴天植物的光能利用率更高, 主要因为阴天有较多的散射光及植物对散射光较高的光合利用效率; 相应地, 外界光照强度较大时, 毛竹的光合作用较强, 但是初始光能利用率却相对降低, 因为此时有多余的光能供利用。
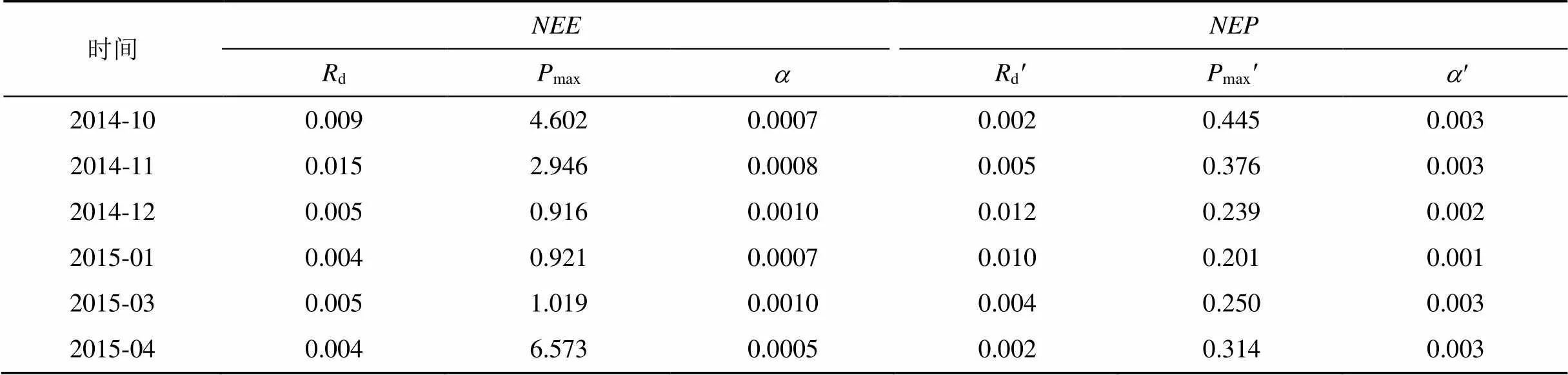
表1 白天NEE/NEP—PAR拟合参数
用非直角双曲线拟合基于生理生态学方法测得的安吉毛竹林生态系统白天净初级生产力与的响应曲线而得到d、max和(表1), 可知, 各月d先升高后降低, 变化范围为0.002—0.012 mg·m–2·s–1, 低于Michaelis-Menten方程拟合的结果; 与d变化相反,max和先降低再升高, 明显低于max, 但高于。主要是因为利用封闭叶室在可控条件下进行测定, 排除了其他因子的干扰, 就只有毛竹单一物种的光合, 对象是毛竹叶片, 且经过光诱导, 在给予其不同的光照强度时, 其光能利用效率较高; 此外, 冬季寒冷干燥, 温度和水分同时控制的植物的呼吸, 使得毛竹叶片的呼吸相对较低。而的测定是在自然条件下测定, 存在着诸多不可控因素, 就算是白天, 除了毛竹自身的CO2释放外, 还有林下其他植物的光合和呼吸作用以及土壤呼吸等, 因此d和max较高。总之,的测定的成分复杂, 要想确定各个组分, 须结合更详细的生理生态方法, 具体有待进一步研究。
4 讨论
利用涡度相关法和生理生态法对安吉毛竹林非生长季(2014年10月—2015年4月, 无2月)的CO2通量进行测定, 总体上, 涡度相关法与生理生态法(光合)响应关系较好。
4.1 日最大值比较
设定用涡度相关技术观测得的净初级生产力为1, 基于光合测定的为2。通过2.2和2.3知道两种方法测定毛竹林非生长季各月白天的日变化均呈现倒“U”型变化, 中午有最大值, 但由于有饱和光强,2达到饱和光强后, 会稳定在最大值附近波动, 直到光强低于饱和值。表2比较了两种方法测定的各月日变化的最大值。可知, 各月日2最大值均低于1, 即生理生态法低估了生态系统日变化的最大值。分析原因,1是利用通量塔进行测定, 能够在一个较大的时间和空间尺度上估计系统的总生产力, 除了受到系统边缘农田作物的CO2吸收的影响外, 在系统内部测定的组分较多, 也比较全面, 克服了通过测定室测定后累加所导致的不确定性[18]; 而2是基于毛竹叶片的光合测定的, 测定组分较为狭窄, 不包括除了毛竹叶片以外的树干、枝条等进行光合作用吸收CO2的部分; 而且虽然此生态系统中近似毛竹纯林, 但还是避免不了存在着其他的一些物种, 如林下草本、苔藓类植物、蕨类等, 能够吸收CO2进行自身的生理活动, 而这些都是2所不包括的; 此外, 仪器配置的光源为人工冷光源, 虽尽可能模仿自然光, 但是散射光对叶片光合的影响很难模拟[18]; 加上毛竹的叶面积指数利用鱼眼镜头拍摄分析, 受光照影响大, 也很大程度地影响着2, 最终导致低估该生态系统2日变化的最大值。另一方面,2虽较低, 但占到1的40%以上, 在毛竹光合作用较旺盛的10月、11月和4月, 所占比例更大。表明毛竹林通过光合作用固定的CO2对生态系统有着较大的贡献, 即在白天, 安吉毛竹林生态系统的还是主要取决于毛竹的光合作用。
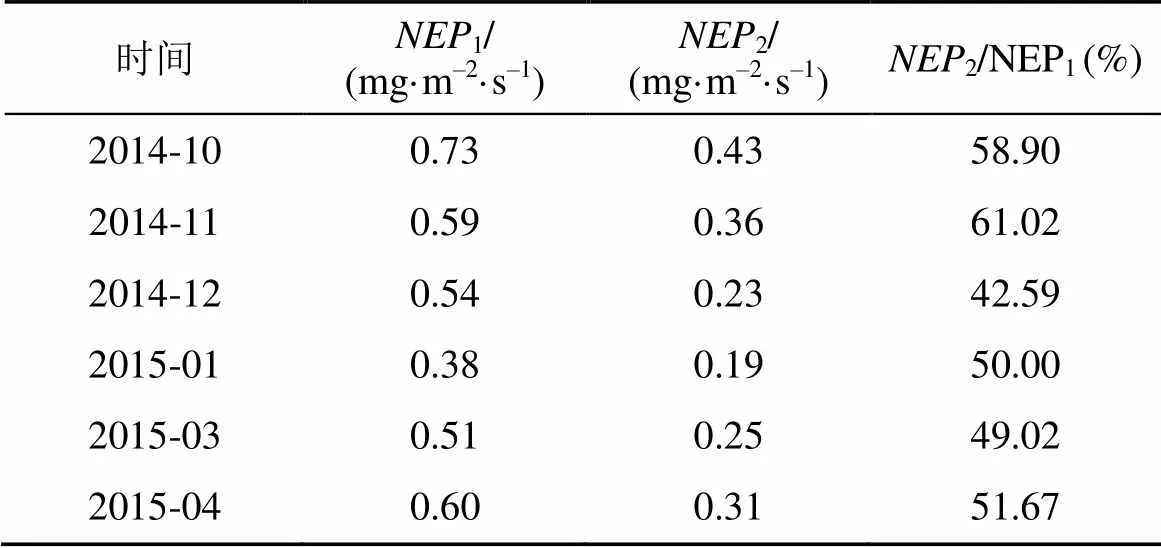
表2 基于涡度相关技术和光合的日NEP最大值比较
注:1是涡度相关技术观测得的净初级生产力,2是基于光合测定的净初级生产力。
4.2 月总量比较
设定用涡度相关技术观测得的各月总净初级生产力为1T, 白天的净初级生产力为1D, 基于光合测定的净初级生产力为2。表3比较了基于两种方法测定的毛竹林非生长季(2014年10月—2015年4月, 无2月)各月的总量。
可知, 毛竹林生态系统非生长季的净初级生产力总量(1T为1365.59 g·m–2, 吸收的碳约372.44 gC·m–2,整体表现较强的碳汇, 高于相似环境中的次生栎林生态系统(为碳源)[39], 也高于环境迥异的长白山红松针阔叶混交林(–127 gC·m–2, 为碳源)[28]。其中10月最高, 达到330.35g·m–2, 占总量的24.19%, 而1月最低, 仅94.38g·m–2, 占总量的6.91%, 3、4月又高于1月, 平均为269.88 g·m–2, 从而表现出明显的季节变化, 即秋末最大, 早春次之, 冬季最低。
就白天而言,1D总量为2237.40 g·m–2,2为1900.33 g·m–2, 并占到1D的84.93%, 比例较高。各月2均低于1D, 主要是因为2测定组分单一, 不包括除毛竹叶片以外的其他能够吸收消耗CO2的部分, 王文杰等[18]的研究也表明, 需要考虑到林下植物的光合能力, 才能够是涡度协方差法计算的结果与生理生态学方法测定结果吻合。而且测量时不能模拟散射光, 以及叶面积指数偏低也会导致2被低估; 但是, 各月2所占比例偏高, 尤其是毛竹光合能力比较强的10、11和4月, 占到96%以上, 甚至4月高达98.06%, 分析原因, 首先, 利用涡度相关技术观测CO2通量, 虽然比较全面, 受环境干扰小, 时间分辨率也较高, 但是地势不平坦、平流干扰都会影响其观测精度[30]。其次, 通量数据的处理, 包括一些极端天气数据的收集与整理, 会影响最终得到的1D。第三, 叶片光响应曲线的测定均是在毛竹生长良好的晴天进行的, 且采用的是封闭叶室, 受外界环境影响较小, 没有阴雨天的干扰。最后, 拟合光响应曲线的时候, 常常预测值略高于实测值, 甚至实测值会有“光抑制”, 从而使得2被高估。
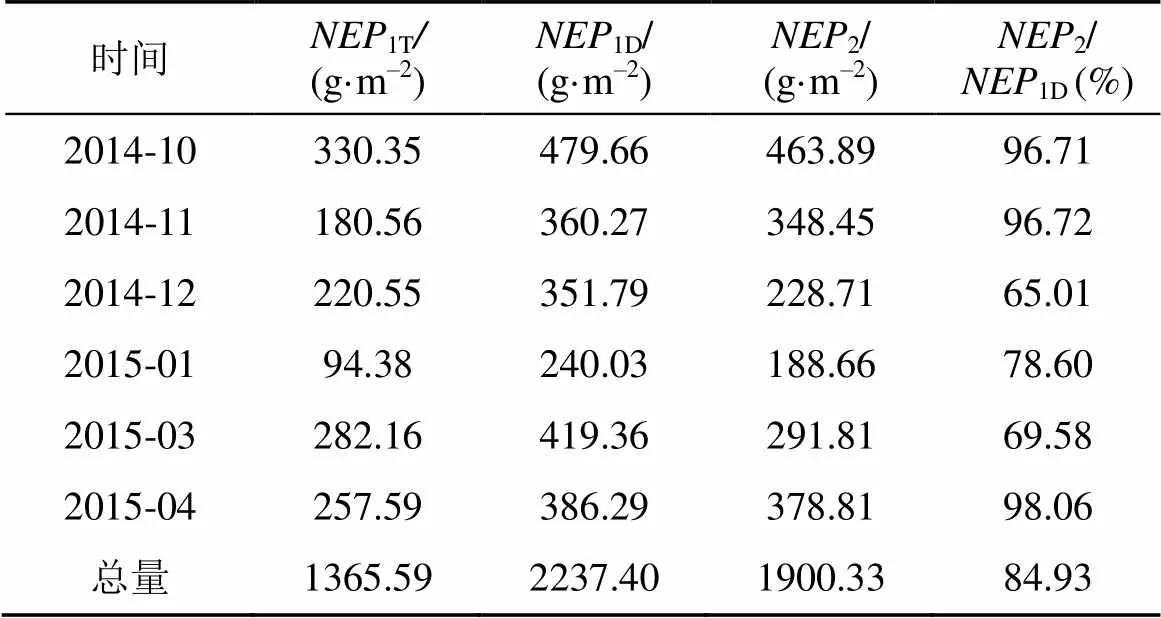
表3 基于涡度相关技术和光合的NEP总量的比较
注:1T是涡度相关技术观测得的净初级生产力总量,1D是基于涡度相关技术测定的白天的净初级生产力,2是基于光合测定的净初级生产力。
另外, 从表3还可以看出, 非生长季各月的1T均低于其1D和2, 主要是因为在非生长季, 植被、凋落物及土壤微生物的呼吸作用较为旺盛, 甚至森林主要的CO2交换由光合吸收转变为呼吸释放, 冬季最为明显, 从而对毛竹林生态系统净CO2的吸收具有抑制作用。11月份受阴雨天气影响, 土壤湿度大, 其土壤呼吸更强, 这在Michael等[40]以及王春林等[27]的研究中也得到证实。总之, 毛竹林生态系统的CO2通量变化受系统内多个组分和过程的影响, 而不仅仅是毛竹的光合作用, 要明确毛竹林生态系统净交换量的生理生态学基础, 还需要对系统内多个组分进行分别观测。
5 结论
利用涡度相关法和生理生态法对安吉毛竹林非生长季(2014年10月—2015年4月, 无2月)的CO2通量进行测定, 可知, 涡度相关法与生理生态法(光合)响应关系较好, 均具有明显的季节变化: 秋季最高, 春季次之, 冬季最低。两种方法测定的毛竹林生态系统净初级生产力()日变化均呈倒“U”型曲线变化, 受值的直接影响, 在午间12:00前后有最大值, 10月最高。但由于光合作用有饱和光强, 故基于光合的2达到饱和光强后, 会稳定在最大值附近波动, 直到光强低于饱和值。
就各月而言, 生理法测定的2日变化最大值均低于涡度相关技术测定的1, 但均占到1的40%以上;2月总量也均低于涡度相关技术白天测定的1D, 但2所占比例均在65%以上, 尤其是毛竹光合能力比较强的10、11和4月, 所占比例更大。表明毛竹林通过光合作用固定的CO2对生态系统有着较大的贡献, 即在白天, 安吉毛竹林生态系统的还是主要取决于毛竹的光合作用。另外, 观测期间, 毛竹林仍有一定的碳吸收能力, 该生态系统的净初级生产力总量(1T)为1365.59 gCO2·m–2, 吸收的碳约372.44 gC·m–2, 整体表现为较强的碳汇(碳吸收)。
[1] 方精云. 全球生态学: 气候变化的生态响应[M]. 北京: 高等教育出版社, 2000: 56.
[2] GIFFORD D M. The global carbon cycle: a viewpoint on the missing sink[J]. Australian Journal of Plant Physiology, 21(1): 1–15.
[3] 王效科, 冯宗炜. 森林生态系统生物碳储存量的研究历史[M]//王如松. 现代生态学的热点问题研究. 北京: 中国科学技术出版社, 1996: 335–347.
[4] 崔崧. 基于微气象学方法的落叶松人工林CO2通量研究[D]. 哈尔滨: 东北林业大学, 2007.
[5] BONAN G B. Forests and climate change: forcings, feedbacks, and the climate benefits of forests[J]. Science, 2008, 320(5882): 1444–1449.
[6] O'GRANDY A P, TISSUE D T, BEADLE C L. Canopy processes in a changing climate[J]. Tree Physiology, 2011, 31(9): 887–892.
[7] FANG Jingyun, CHEN Anping, PENG Changhui, et al. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 2001, 292(5525): 2320–2322.
[8] PIAO Shilong, FANG Jingyun, CIAIS P, et al. The carbon balance of terrestrial ecosystems in China[J]. Nature, 2009, 458 (7241): 1009–1013.
[9] 孙成, 江洪, 周国模, 等. 我国亚热带毛竹林CO2通量的变异特征[J]. 应用生态学报, 2013, 24(10): 2717–2724.
[10] 李睿, 维尔格, 钟章成. 施肥对毛竹竹笋生长的影响[J]. 植物生态学报, 1997, 21(1): 19–26.
[11] 马少杰, 李正才, 王刚, 等. 集约和粗放经营下毛竹林土壤活性有机碳的变化[J]. 植物生态学报, 2011, 35(5): 551–557.
[12] 刘玉莉, 江洪, 陈健, 等. 安吉毛竹林碳水通量及水分利用效率的日动态研究[J]. 生态科学, 2015, 34(4): 43–51.
[13] 杨爽. 浙江安吉毛竹林生态系统CO2通量观测研究[D]. 临安: 浙江农林大学, 2012.
[14] VERMA S B, BALDOCCHI D D, ANDERSON D E, et al. Eddy fluxes of CO2, water vapor, and sensible heat over a deciduous forest[J]. Boundary Layer Meteorology, 1986, 36: 71–91.
[15] BALDOCCARDO D, VALENTINI R, RUNNING S, et al. Strategies for measuring and modeling carbon dioxide and water vapor fluxes over terrestrial ecosystem [J]. Global Change Biology, 1996, 2(3): 159–168.
[16] LEE X H. On micrometeorological observation of surface- air exchange over tall vegetation [J]. Agricultural and Forest Meteorology, 1998, 91: 39–49.
[17] FALGE E, BALDOCCHI D, OLSON R, et al. Gap filling strategies for defensible annual sums of net ecosystem exchange[J]. Agricultural and Forest Meteorology, 2001, 107(1): 43–69.
[18] 王文杰, 祖元刚, 王辉民, 等. 基于涡度协方差法和生理生态法对落叶松CO2通量的初步研究[J]. 植物生态学报, 2007, 31(1): 118–128.
[19] 张宪洲, 石培礼, 刘允芬, 等. 青藏高原高寒草原生态系统土壤CO2排放及其碳平衡[J]. 中国科学D辑地球科学, 2004, 34(增刊II): 193–199.
[20] 王玉辉, 井长青, 白洁, 等. 亚洲东部干旱区3个典型生态系统生长季水碳通量特征[J]. 植物生态学报, 2014, 38(8): 795–808.
[21] 朱咏莉, 童成立, 吴金水, 等. 亚热带稻田生态系统CO2通量的季节变化特征[J]. 环境科学, 2007, 28(2): 283–288.
[22] 周丽艳, 贾丙瑞, 周广胜, 等. 中国北方针叶林生长季碳交换及其调控机制[J]. 应用生态学报, 2010, 21(10): 2449–2456.
[23] 刘允芬, 宋霞, 孙晓敏, 等. 千烟洲人工针叶林CO2通量季节变化及其环境因子的影响[J]. 中国科学D辑地球科学, 2004, 34(增刊II): 109–117.
[24] 邱岭, 祖元刚, 王文杰, 等. 帽儿山地区落叶松人工林CO2通量特征及对林分碳收支的影响[J]. 应用生态学学报, 2011, 22(1): 1–8.
[25] 周丽艳, 曹丙瑞, 曾伟, 等. 原始兴安落叶松林生长季净生态系统CO2交换及其光响应特征[J]. 生态学报, 2010, 30(24): 6919–6926.
[26] 王春林, 周国逸, 王旭, 等. 鼎湖山针阔叶混交林冠层下方CO2通量及其环境响应[J]. 生态学报, 2007, 27(3): 846–854.
[27] 王春林, 于贵瑞, 周国逸, 等. 鼎湖山常绿针阔叶混交林CO2通量估算[J]. 中国科学D辑地球科学, 2006, 36(增刊I): 119–129.
[28] 吴家兵, 关德新, 施婷婷, 等. 非生长季长白山红松针阔叶混交林CO2通量特征[J]. 林业科学, 2006, 4(9): 1–6.
[29] 沈艳, 缪启龙, 刘允芬. 亚热带红壤丘陵人工混交林区CO2源汇及变化[J]. 生态学报, 2005, 25(6): 1371–1375.
[30] 于成龙, 刘丹. 小兴安岭天然阔叶混交林生长季CO2通量特征分析[J]. 中国农业气象, 2011, 32(4): 525–529.
[31] 焦振, 王传宽, 王兴昌. 温带落叶阔叶林冠层CO2浓度的时空变异[J]. 植物生态学报, 2011, 35(5): 512–522.
[32] 赵晓松, 关德新, 吴家兵, 等. 长白山阔叶红松林CO2通量与温度的关系[J]. 生态学报, 2006, 26(4): 1088–1095.
[33] 赵双菊, 张一平, 于贵瑞, 等. 西双版纳热带季雨林晴天CO2交换的日变化和季节变化特征[J]. 植物生态学报, 2006, 30(2): 295–301.
[34] 陈云飞, 江洪, 周国模, 等. 人工高效经营雷竹林CO2通量估算及季节变化特征[J]. 生态学报, 2013, 33(11): 3434–3444.
[35] 刘允芬, 于贵瑞, 温学发, 等. 千烟洲中亚热带人工林生态系统CO2通量的季节变异特征[J]. 中国科学D辑地球科学, 2006, 36(增刊I): 91–102.
[36] HOLLINGER D Y, KELLIHER F M, BYERS J N, et al. Carbon dioxide exchange between an undisturbed old- growth temperate forest and the atmosphere[J]. Ecology, 1994, 75: 134–150.
[37] LOESCHER H W, OBERBAUER S F, GHOLZ H L, et al. Environmental controls on net ecosystem-level carbon exchange and productivity in a Central American tropical wet forest[J]. Global Change Biology, 2003, 9: 396–412.
[38] 蔺恩杰, 江洪, 陈云飞. 太湖源雷竹林水汽通量变化及其对净辐射的响应[J]. 浙江农林大学学报, 2013, 30(3): 313–318
[39] 刘乙, 胡海波, 刘准桥. 北亚热带次生栎林生态系统非生长季CO2通量特征[J]. 东北林业大学学报, 2013, 41(7): 22–27.
[40] MICHAEL L G, BRUCE C D, FAN S M, et al. Physiological responses of a black spruce forest to weather[J]. Journal of geophysical research, 1997, 102: 28987–28996.
Study on the response relationship between CO2flux and photosynthesis offorest during non-growing season in Anji, Zhejiang
SHU Haiyan1, JIANG Hong1,2,3*, CHEN Xiaofeng2, SUN Wenwen2, MA Jinli2, ZHANG Minxia2
1. Key Laboratory of Eco-environments of Three Gorges Reservoir Region, Ministry of Education, Southwest University, Chongqing 400715, China 2. College of Forestry & Bio-technology, Zhejiang A & F University, Lin’an, Zhejiang 311300, China 3. International Institute for Earth System Science, Nanjing University, Nanjing 210046, China
CO2fluxes offorest ecosystem during non-growing season (from October in 2014 to April in 2015, but no February) in Anji, Zhejiang Province were measured by eddy covariance micrometeorological and eco-physiological method (photosynthesis of leaves). Both the results showed a good relationship. Diurnal variations of the net ecosystem exchange () based on eddy covariance micrometeorological method and the net ecosystem productivity () based on the eco-physiological method were single peak curves, and the maximum values were at around 12:00 noon, which were influenced bydirectly. Moreover,andwere the highest in autumn (October was the highest) and the lowest in winter. For months, the daily maximums and monthly totals ofbased on photosynthesis method were lower than those measured by the eddy covariance technique during the day, but each accounted to over 40% and 65% out of the latter respectively. Especially in October, November and April, the proportions were greater because of the high photosynthesis of. What is more, during the non-growing season, the totalofforest ecosystem was 1365.59 gCO2·m–2, and the absorption of carbon was about 372.44 gC·m–2. That was, the ecosystem was a moderate carbon sink outside the growing season.
forest ecosystem; non-growing season; eddy covariance technique; photosynthesis; net ecosystem productivity
10.14108/j.cnki.1008-8873.2017.02.003
Q945.79 , S718.5
A
1008-8873(2017)02-015-11
2015-09-02;
2015-12-03
国家自然科学基金(61190114和41171324); 上海市科学委员会项目(12231205101); 科技部国家科技基础条件平台项目“地球系统科学数据共享网”(2005DKA32300)
舒海燕(1990—), 女, 四川广元人, 硕士生, 主要从事植物生理生态学研究, E-mail: shu001297@163.com
江洪, 男, 博士, 教授, 主要从事全球变化等方向研究, E-mail: jianghong_china@hotmail.com
舒海燕,江洪,陈晓峰, 等. 安吉毛竹林非生长季CO2通量与其光合响应关系探究[J]. 生态科学, 2017, 36(2): 15-25.
SHU Haiyan, JIANG Hong, CHEN Xiaofeng, et al. Study on the response relationship between CO2flux and photosynthesis offorest during non-growing season in Anji, Zhejiang[J]. Ecological Science, 2017, 36(2): 15-25.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
中国海洋大学学报(自然科学版)(2019年11期)2019-10-12
热带气象学报(2019年6期)2019-03-02
乡村科技(2018年8期)2018-06-27
科技资讯(2018年30期)2018-03-11
湖南林业科技(2017年1期)2017-02-06
大气科学学报(2014年5期)2014-09-11
