
河川沙塘鳢的形态指标体系及雌雄差异分析
2018-04-13 00:35张宏叶陈树桥周国勤尹绍武
江苏农业科学 2018年6期
张宏叶, 陈树桥, 王 涛, 周国勤, 尹绍武
(1.南京师范大学生命科学学院/江苏省生物多样性与生物技术重点实验室,江苏南京210023;2.南京市水产科学研究所,江苏南京210036)
河川沙塘鳢(Odontobutispotamophila)属鲈形目(Perciformes)沙塘鳢科(Odontobutidae)沙塘鳢属(Odontobutis),俗称虎头鲨、土布鱼、虎头呆子等。河川沙塘鳢是个体较小的一种淡水底栖型鱼类,为中国特有,广泛分布于长江中下游、钱塘江、闽江等水系。近年来,由于自然水域污染加剧,河川沙塘鳢的生存环境不断恶化,产卵环境遭到破坏,其野生资源急剧衰退,部分地区已难觅踪迹。目前,该鱼主要靠捕捞供应市场,但远不能满足市场需求,价格不断攀升。因此,河川沙塘鳢已发展成为一个非常具有潜力的水产养殖新品种[1-3]。
目前,我国研究学者对河川沙塘鳢进行了大量的研究,取得了一系列成果,如通过苗种培育选育出了建德×当涂(正反交)最佳杂交组合,具有生长快、产量高、抗逆性强的优良性状[4-5]。种群结构与遗传多样性研究显示,河川沙塘鳢具有高的单倍型多样性、低的核苷酸多样性和低的遗传多样性,这将为河川沙塘鳢的遗传多样性监测、物种保护及其人工繁殖等工作提供科学依据[6]。河川沙塘鳢高密度遗传连锁图谱显示,总共6 311标记作图在包含SLAF标记物理位置的22个连锁群,并检测出共9 468 SNP位点,这张高密度遗传连锁图谱可为将来获得该鱼生长QTL相关SNP标记和分子标记辅助选择提供有用的平台[7]。运用微卫星分子标记技术对河川沙塘鳢家系的遗传多样性研究表明,9个微卫星位点在5个家系(F1、F16、F29、F35和F36)中总共检测到128个等位基因,且各家系间存在一定的遗传分化,这为河川沙塘鳢家系选育提供相关数据,为下一步良种选育奠定基础[8]。然而到目前为止,关于河川沙塘鳢形态特征的研究报道较少[9],对应到河川沙塘鳢雌雄形态差异的研究尚未见报道。

1 材料与方法
1.1 试验材料
试验用鱼分2批,均为池塘养殖鱼类。分别于2015年9月和2016年12月,取自江苏南京市水产科学研究所周岗基地。第1批计115尾,体长为3.17~7.11 cm,体质量为0.78~9.89 g;第2批计220尾,体长为7.90~13.20 cm,体质量为13.20~59.60 g。
1.2 数据测量
采用游标卡尺(573-642,昆山铭尚精密机电有限公司)测量全长(overall length)、体长(body length)、体高(body height)、体厚(body thickness)、尾柄长(caudal peduncle length)、尾柄高(caudal peduncle height)、头长(head length)等形态性状指标,精确至0.01 cm;使用电子天平(JSC-T29-6,惠而邦电子衡器有限公司)称量体质量,精确至0.01 g。丰满度(fatness)通过公式:体质量/体长3×100%计算得到,测定形态指标性状指标后,再逐一解剖观察其性腺特征判断性别。
1.3 数据处理
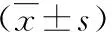
2 结果
2.1 河川沙塘鳢雌雄形态性状分析
由表1可知,河川沙塘鳢雌雄各计量性状均值及其差异系数。雌雄鱼计量性状变异最大的均为体质量,变异系数分别为0.67和0.57。从7项计量性状来看,在个体间大小差异方面,雄鱼个体间产生的变异较雌鱼大,雌鱼相对均匀。
为了分析雌雄间形态特征的差异,将雌雄计量性状的均值差值和总体均值作比以进行标准化(MD/Mean),可得出在所有计量性状中雌雄鱼的体质量差异最大,其均值差/均值达到0.47,其次是尾柄高和体高的差异,该值分别为0.33和0.29,全长和体长的均值差/均值相对较低,均为0.22。
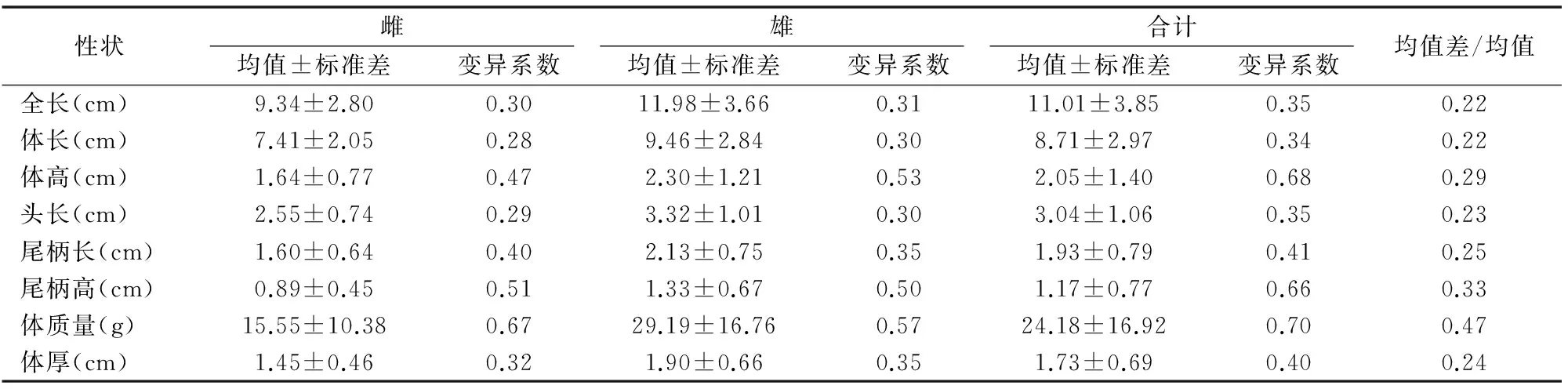
表1 河川沙塘鳢雌雄计量性状比较
2.2 河川沙塘鳢主成分分析
为简化河川沙塘鳢外形特征,明确河川沙塘鳢的形态指标体系,对14项性状进行主成分分析,筛选出前4个主成分贡献率,以及各指标在这4个主成分上的负荷量,它们的累积贡献率达到86.46%,4个主成分及其包含的性状和负荷量见表2。其中,主成分Ⅰ贡献率达到58.26%,负荷量较大的主要是全长、体长、体质量、体高、尾柄高、体厚、头长、尾柄长、体长/体高、体长/体厚10项性状,反映了河川沙塘鳢的整体框架轮廓指标;主成分Ⅱ贡献率为10.91%,包括肥满度、体高、体高/体厚3项性状,反映了河川沙塘鳢体型肥瘦程度的指标;主成分Ⅲ贡献率为9.65%,包括体长/尾柄长、尾柄长/尾柄高2项性状,反映了河川沙塘鳢躯干及尾部指标;主成分Ⅳ贡献率为7.82%,包括头长、体长/头长2项性状,反映了河川沙塘鳢头部特征。
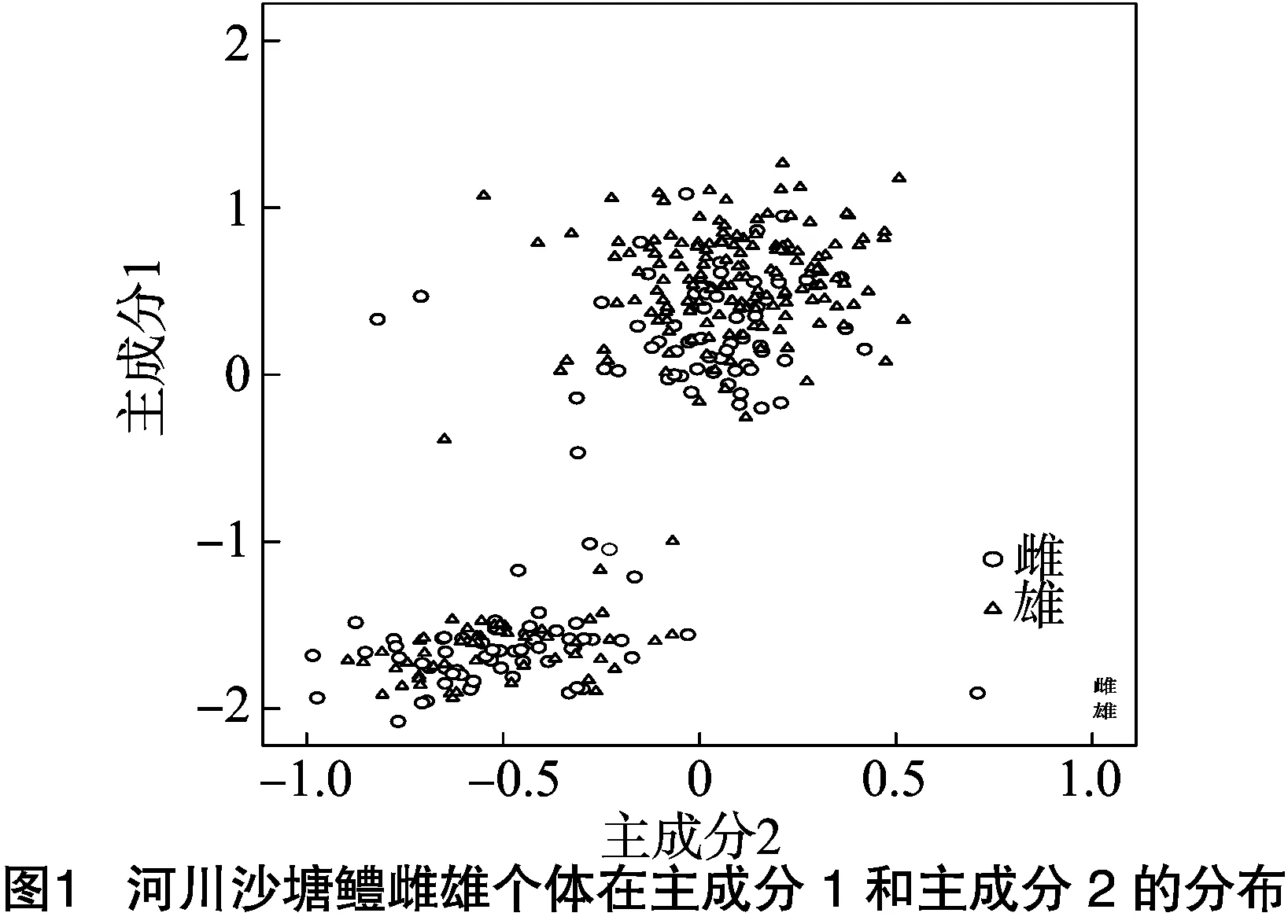
根据主成分分析的得分,分别对主成分Ⅰ与主成分Ⅱ及主成分Ⅰ与主成分Ⅲ之间的雌雄个体绘制散布图,见图1、图2。图1中雌雄个体分布在主成分Ⅰ、Ⅱ轴上均表现出明显的分离态势,通过得分可知差异主要由体厚造成。图2中雌雄个体在主成分Ⅰ、Ⅲ轴上均表现稍有分离态势,而在主成分Ⅰ上更明显,比较各个性状的得分,可知其差异主要由体厚、尾柄长和尾柄高导致,散布图显示雌雄个体可能在体厚和尾部形态上稍有差异。
2.3 河川沙塘鳢相对性状的R-聚类分析
由图3可知,对335尾河川沙塘鳢的7项标准化形态性状进行R-聚类分析的结果。此标准化后的相对性状可分为两大类。第Ⅰ类为体长/头长、体长/尾柄长、尾柄长/尾柄高,为体形的特征参数;第Ⅱ类为体高/体厚、体长/体厚、体长/体高及肥满度,主要反映鱼体丰满度。
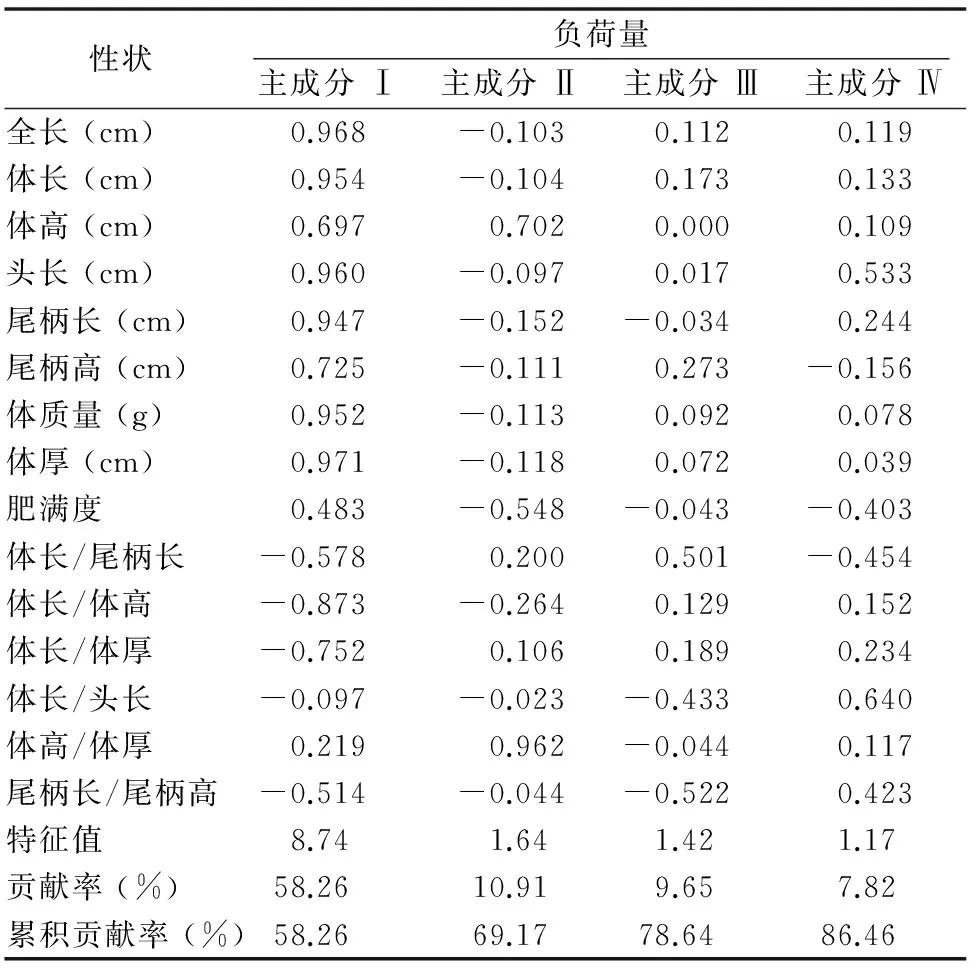
表2 4个主成分贡献率及各指标负荷量
R-聚类划分的第Ⅰ类标准化性状与主成分分析所得的第Ⅲ、Ⅳ主成分的组分相对应,均反映河川沙塘鳢头部及尾部指标; 第Ⅱ类标准化性状也可与主成分分析所得的第Ⅱ组组分相对应,反映了河川沙塘鳢体型肥瘦程度的指标。两者所得的结果大体一致。


2.4 河川沙塘鳢雌雄判别分析
用SPSS 19.0软件对所收集的335尾河川沙塘鳢个体的体长、体质量连同7项标准化性状(表2)进行逐步判别分析,根据各变量对模型的贡献大小,逐步剔除不相关变量,最终筛选出4个变量,即体长(X1)、体质量(X2)、体长/体厚(X3)、体长/头长(X4)(变量的单位均为%)。这几个指标反映了雌雄个体在头部形态(X4)、身体形态(X1、X2、X3)方面的差异。所建立的判别模型方程为
雌:F1=6.543X1-0.546X2+20.634X3+40.504X4-153.512;
雄:F2=6.053X1-0.388X2+21.296X3+39.396X4-134.657。
判别函数的显著性检验显示,P<0.01,表明判别函数达到极显著水平。计算出雌雄个体的判别分数值,分别得到雌雄个体的频布图。由图4、图5可见,该模型可以用于区分河川沙塘鳢的性别。
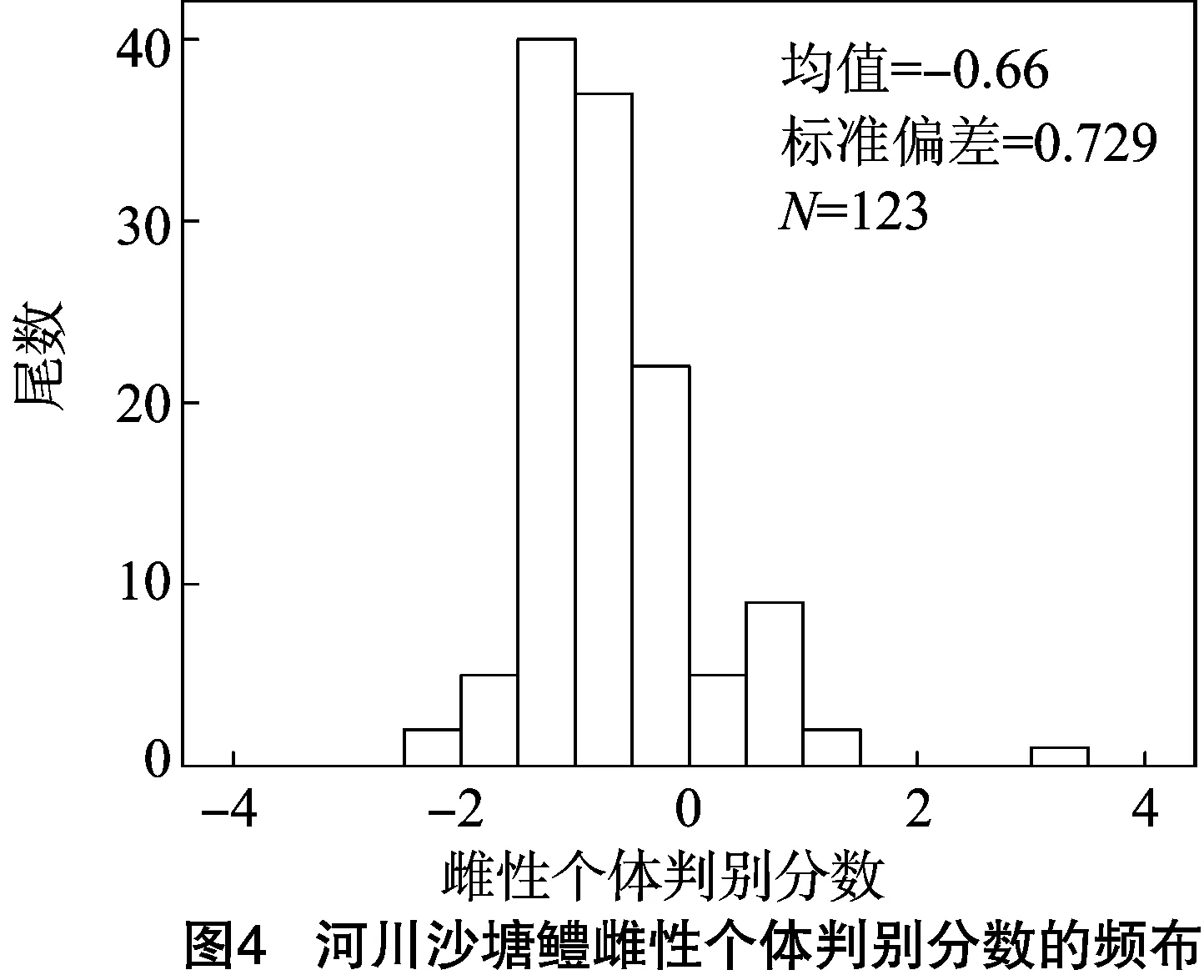

利用所建立的判别方程,对所建立方程的335尾个体进行性别回判。将每尾个体的4个特征参数值代入方程,分别计算F1和F2。如F1>F2,则为雌性,反之为雄性。经过验证,雌性判别准确率为74.80%,雄性判别准确率为68.90%,综合判别率为71.00%(表3)。

表3 河川沙塘鳢雌雄判别结果
对筛选出的用于建立判别函数的4个性状进行t检验,结果显示体长、体质量和体长/体厚的差异极显著(P<0.01);体长/头长的差异显著(P<0.05)。以上数据表明,雄性个体的体型相对于雌性较宽较丰满,躯干和头较长(表4)。

表4 河川沙塘鳢性状的t检验及雌雄形态差异
3 讨论
形态学研究是河川沙塘鳢养殖及育种研究的重要内容,也是开展性别差异分析,进行河川沙塘鳢性别鉴定和控制的基础[16]。近年来,许多学者将主成分分析、R-聚类分析和判别分析等多元分析的方法应用于水产动物雌雄个体形态差异的分析上。丁文超等采用聚类分析、主成分分析和判别分析的方法对大黄鱼(Larimichthyscrocea)4个家系(反交家系、岱衢洋家系、官井洋家系和正交家系)的形态差异进行了分析,结果表明岱衢洋家系、官井洋家系和正交家系之间形态差异较小,而它们同反交家系差异显著[17]。郭弘艺等对153尾银色鳗(Anguillajaponica)的形态指标进行主成分分析,标准化比例性状进行R-聚类分析,并建立了雌雄个体的判别模型,结果表明长江口银色鳗的雌性个体比雄性更为丰满,体型较高但眼较小[18]。吴波等对178尾金钱鱼(Scatophagusargus)形态性状进行主成分分析和R-聚类分析,建立了金钱鱼雌雄判别方程,结果表明雌性个体相比于雄性具有体形较高、头较大且吻较长的特征[19]。岳亮等采用主成分分析、逐步判别分析和t检验的方法对红鳍东方鲀(Takifugurubripes)雌雄个体的形态差异进行了分析研究,结果表明红鳍东方鲀雄性与雌性相比,体型较宽,体周长较长[20]。安丽等采用主成分分析、R-聚类分析和判别分析对67尾翘嘴红鲌(Eryghroculterilishaeformis)主要形态指标和雌雄个体的形态差异进行了分析研究,结果表明雌性相较于雄性体型较宽、较丰满,但尾柄较矮[21]。但到目前为止,通过主成分分析、聚类分析和判别分析的方法对河川沙塘鳢雌雄个体间的差异及其主要形态指标却未见报道。因此,为明确河川沙塘鳢的主要形态指标体系,找出雌雄个体形态上存在的差异,本研究对335尾河川沙塘鳢的14项性状(7项计量性状及7项标准化性状)进行了主成分分析、R-聚类分析和判别分析。
主成分分析结果显示,共得到4个主成分,累计贡献率为86.46%,这4个主成分分别代表河川沙塘鳢的整体框架结构指标、肥瘦程度指标、躯干及尾部指标和头部指标。通过对这些指标的测定和分析可以基本反映河川沙塘鳢的外形特征。聚类分析与主成分分析所得结果基本一致,聚类分析中的第Ⅰ、Ⅱ类标准化性状分别与主成分分析所得的第Ⅲ、Ⅳ和Ⅱ类主成分的组分相对应,反映河川沙塘鳢头部、尾部及体型肥瘦程度的指标。表明河川沙塘鳢个体差异主要集中在头部形态和鱼体形态等方面。通过主成分分析的散布图可知雌雄个体主要在主成分Ⅰ轴上表现出明显的分离态势,说明雌雄个体在主成分Ⅰ的指标有差异。根据得分情况可知,体厚对此分离的作用较大,分析其原因可能是在实际操作中对其测量时产生的误差所导致,因此造成雌雄分析在体厚上有差异。
对河川沙塘鳢雌雄形态性状的分析发现,雌雄鱼计量性状变异最大的均为体质量,变异系数分别为0.67及0.57,雌雄间差异较大。从7项计量性状来看,在个体间大小差异方面,雄鱼个体间产生的变异较雌鱼大,雄鱼更为丰满,体型为厚胖型,而雌鱼体型为扁瘦型。此结果在下面的判别分析中也可以得到验证。运用逐步判别分析的t-检验结果表明,河川沙塘鳢雌雄差异主要集中在体型肥瘦程度和头部形态上,即雄性相较于雌性体型显著较宽、较丰满、头较长,这与谌微等对大黄鱼(Larimichthyscrocea)[22]、安丽等对翘嘴红鲌(Eryghroculterilishaeformis)的研究结果[21]相似。同时,本研究建立了雌雄判别方程式,最终保留了体长、体质量、体长/体厚和体长/头长4个变量,综合判别成功率达71.00%。郭弘艺等对长江口银色鳗(Anguillajaponica)雌雄个体建立的判别方程分析显示群体的识别正确率达88.50%[18];吴波等对金钱鱼(Scatophagusargus)雌雄个体建立的性别判别方程分析显示,其综合判别率为85.96%[19];岳亮等对72尾雌、雄红鳍东方鲀(Takifugurubripes)的判别研究显示,其准确率为81.90%[20]。与之相比,本研究结果相对处于较低水平,推测可能是由于试验鱼个体间差异较大造成的,雌雄体质量变异系数分别达到0.76及0.57,个体间体质量相差较大,使标准化很难进行,这样不利于差异比较和雌雄判别。其次,试验所选择的性状不够全面也是可能原因之一,在进行判别分析时得到的标准化性状有限,使建立的雌雄判别函数方程式有所差异,故对总体的判别准确率造成一定的影响。后续研究应增加眼径、吻长、眼间距等表型数据来进行相关的分析。
虽然判别分析和t-检验均不能有效地对河川沙塘鳢雌雄进行判别,这并不意味着多元分析的方法不适用于河川沙塘鳢雌雄判别,本研究由于受限于试验个体间差异大、形态指标选取不完善,导致判别结果不理想。事实上,通过本研究已经确定河川沙塘鳢雌雄形态存在明显差异,多个性状差异均达到极显著水平,这也证明河川沙塘鳢雄鱼生长明显快于雌鱼。但是要找出河川沙塘鳢雌雄生长出现差异的阶段,并从外形上对雌雄鱼进行准确判别,还需更加深入地研究。在以后的性别形态差异研究中,需充分考虑个体间差异大小、鱼体规格以及形态指标选取完善与否等这些因素。
参考文献:
[1]Hou X,Zhu F,Yin S,et al. Genetic diversity ofOdontobutispotamophilafrom different geographic populations inferred from mtDNA control region[J]. Mitochondrial DNA,2013,25(5):400.
[2]朱永安,王南明,杨德先.沙塘鳢耗氧率与窒息点的测定[J]. 内陆水产,2005,30(9):35-36.
[3]乔德亮,洪磊. 淮河水系沙塘鳢形态特征和分类地位初步研究[J]. 淡水渔业,2007,37(2):20-23.
[4]王佩佩,丁严冬,于兴达,等. 河川沙塘鳢3个不同地理群体杂交 F1代生长性能及遗传多样性[J]. 水产科学,2016,35(5):528-534.
[5]陈树桥,周国勤,张厚冰,等. 河川沙塘鳢大规格苗种培育试验[J]. 科学养鱼,2014,30(7):8-9.
[6]侯新远. 河川沙塘鳢群体遗传多样性及其系统进化研究[D]. 南京:南京师范大学,2014:34-49.
[7]Zhang H,Huang L,Yin S,et al. High-density SNP-based genetic map forOdontobutispotamophila[J]. Animal Genetics,2016,48(1):126.
[8]丁严冬,王佩佩,贾秀琪,等. 河川沙塘鳢五个家系遗传多样性的微卫星分析[J]. 水产科技情报,2016,43(3):113-118.
[9]徐宇,史杨白,郑啸宇,等. 河川沙塘鳢表型性状对体重影响的通径分析[J]. 江苏农业科学,2016,44(3):262-264.

[11]林植华,雷焕宗.雌雄两性黄颡鱼头部形态特征的增长[J]. 丽水师范专科学校学报,2004,26(2):39-41.
[12]梅洁,桂建芳.鱼类性别异形和性别决定的遗传基础及其生物技术操控[J]. 中国科学(生命科学),2014,44(12):1198-1212.
[13]Afeworki Y,Videler J J,Berhane Y H,et al. Seasonal and life-phaserelateddifferencesingrowthinScarusferrugineus on a
southern Red Sea fringing reef[J]. Journal of Fish Biology,2014,84(5):1422-1438.
[14]贾一何,黄鹤忠,李倩倩,等. 池养乌苏里拟鲿雌雄鱼生长及周年性激素与性腺发育研究[J]. 海洋科学,2012,36(3):61-66.
[15]朱邦科,谢从新,王明学,等. 保安湖沙塘鳢的食性、繁殖、年龄及生长的研究[J]. 水生生物学报,1999,23(4):316-323.
[16]丁严冬,藏雪,张国松,等. 河川沙塘鳢4个不同地理群体的形态差异分析[J]. 海洋渔业,2015,37(1):24-30.
[17]丁文超,李明云,管丹冬,等. 大黄鱼4个家系的形态差异分析[J]. 宁波大学学报(理工版),2009,22(2):185-190.
[18]郭弘艺,魏凯,谢正丽,等. 长江口银色鳗的形态指标体系及其雌雄鉴别[J]. 水产学报,2011,35(1):1-9.
[19]吴波,张敏智,邓思平,等. 金钱鱼雌雄个体的形态差异分析[J]. 上海海洋大学学报,2014,23(1):64-69.
[20]岳亮,王新安,马爱军,等. 红鳍东方鲀(Takifugurubripes)雌、雄个体的形态特征比较[J]. 渔业科学进展,2016,37(1):30-35.
[21]安丽,孟庆磊,张龙岗,等. 翘嘴红鲌雌雄个体的形态差异分析[J]. 淡水渔业,2017,47(2):36-41.
[22]谌微,王盼盼,肖世俊,等. 大黄鱼形态指标体系及雌雄差异分析[J]. 集美大学学报(自然科学版),2014,19(6):401-408.
猜你喜欢
流程工业(2022年12期)2022-12-30
今日畜牧兽医(2022年10期)2022-12-23
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
东坡赤壁诗词(2018年3期)2018-07-16
黄河之声(2016年8期)2016-10-21
Coco薇(2015年12期)2015-12-10
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
食品科学(2013年17期)2013-03-11
大学(2008年4期)2008-07-10
