
苏丹草转录组SSR分子标记开发及遗传多样性评价
2018-05-22 03:14朱永群彭丹丹林超文聂刚许文志黄琳凯罗付香彭建华张新全
草业学报 2018年5期
朱永群,彭丹丹,林超文,聂刚,许文志,黄琳凯,罗付香,彭建华,张新全*
(1.四川省农业科学院土壤肥料研究所,四川 成都 610066;2.四川农业大学动物科技学院,四川 成都 611130;3.四川省农业科学院,四川 成都 610066)
苏丹草(Sorghumsudanense)系高梁属一年生草本植物,染色体数为20(2n=20),其开花习性与一般禾本科植物不同,属于异花授粉植物,因原产于非洲北部苏丹地区而得名。苏丹草具有耐旱、高产、营养价值丰富等特点,作为一种优质的禾本科牧草在世界各地广泛栽培和使用[1-2]。自20世纪30年代由美国引入后,苏丹草在我国的种植区域不断扩大,主要分布在内蒙古、新疆、甘肃和宁夏等北部省区以及长江中下游地区,不仅可用于放牧、刈割青饲、调制青贮等为家畜所喜食,还具有“渔业青饲料之王”的美称[3-4]。苏丹草与高粱的杂交种高丹草(Sorghumbicolor×Sorghumsudanenes)因具有明显的杂种优势,亦是草业生产中一种重要的牧草,具有显著的经济效益[5]。我国开展苏丹草品种选育和整理工作始于20世纪70年代,但由于缺乏品种在不同条件下生长表现的系统性评价和研究,其选育多凭个人经验,缺乏一定的科学预见性,导致目前为止通过我国草品种审定的苏、高丹草品种数还屈指可数,加之市面上销售的草种子多存在鱼龙混杂的现象,使得这一优良牧草的利用、推广和品种的更新受到严重的阻碍,与目前全国畜牧业和养殖业的大力发展对苏、高丹草生产规模的需求极不协调[6-7]。
简单重复序列(simple sequence repeats,SSRs),是一类由1~6个核苷酸碱基为基本单元,组成的较长串联重复的DNA序列[8]。SSR分子标记具有多态性高、重复性好、覆盖面广、共显性和易于操作等特点,广泛应用于牧草和作物的基因定位、遗传图谱构建、遗传多样性分析及分子标记辅助育种等方面[9-10]。然而,传统SSR分子标记开发过程具有盲目性和随机性,且开发成本高,开发效率低。虽然Zhan等[11]根据Gramene在线数据库设计引物,利用SSR标记对多份苏丹草及高粱材料间的多样性及系统发育关系进行了研究,卢杰等[12]和李杰勤等[13]利用了高粱(Sorghumbicolor)表达序列标签(expressed sequence tags, EST)序列信息建立EST-SSR标记应用于苏丹草证明了标记的可行性,但迄今为止关于专门针对苏丹草开发的SSR分子标记还鲜有报道,且缺乏与性状紧密相关的特异性标记,严重制约了苏丹草分子育种工作的进程。基于高通量测序技术(next generation sequencing, NGS)获得的基因组转录信息用于分子标记的开发,具有简单快捷、高效准确、引物通用性好、成本低等显著优点,此外,还被证明标记可以有效与植物非生物逆境胁迫下抗性和重要的农艺性状等紧密关联[14-16]。因此,利用转录组信息开发SSR分子标记被认为是新时代提高作物遗传多样性和分子标记辅助育种研究准确性的有效途径[17],目前已成功运用于苜蓿(Medicagosativa)[18]、鸭茅(Dactylisglomerata)[19]、芒草(Miscanthus)[20]等牧草种质资源的研究中。在苏丹草测序工作研究方面,虽然Li等[21]先前根据S772苏丹草转录组测序信息开发了SNP和Indel标记,但关于苏丹草转录组SSR标记的开发及应用还有待进一步的研究,这为本工作的开展提供了契机。
本研究基于苏丹草转录组测序结果拟分析SSR位点分布特征,设计开发EST-SSR分子标记,对苏、高丹草共36份材料进行EST-SSR标记的遗传多样性分析,为丰富苏丹草分子标记类型、开发与利用转录组EST资源提供基础,也为进一步开展苏丹草种质资源评价、遗传图谱构建和分子标记辅助育种提供参考。
1 材料与方法
1.1 供试材料与DNA提取
本试验材料为来源于农业部全国畜牧总站国家牧草种质基因库的苏丹草及高丹草种质资源共36份,材料编号及来源地见表1。试验从2017年1月起,对苏丹草种子采用沙培法进行培育。种子经1% 次氯酸钠消毒5 min并用蒸馏水冲洗几遍后播种于盛有石英砂的培育盆中,放入光照培养箱中发芽,待发芽4 d后更换为Hoagland全营养液培养,温度设置为白天25 ℃,夜晚22 ℃,时长各为12 h。待幼苗长至18 d左右(2片成熟叶片),每份种质资源随机选取20个单株的幼嫩叶片等量混合并用植物DNA提取试剂盒(Qiagen)提取DNA(每份材料3个重复)。所提DNA经1.0%的琼脂糖凝胶电泳和紫外分光光度计检测纯度和浓度,将检测合格的DNA样品置于-20 ℃冰箱保存。试验开始前将各DNA样品取出一部分用TE(Tris-EDTA)缓冲液稀释至10 ng·μL-1,置于4 ℃冰箱备用。
1.2 SSR引物设计
在土壤肥料研究所土壤与耕作饲草研究室对乌拉特1号(Wulate No.1)苏丹草转录组测序结果(SRR登录号5183088)得到的13574条包含SSR位点的EST序列的基础上,利用MISA 1.0软件根据设定的单位大小和最小重复参数如1-10、2-6、3-5、4-5、5-5和6-5对位于非重复序列基因(unigenes)上的EST序列设计引物。用Primer 5.0引物设计软件设定参数:鸟嘌呤和胞嘧啶(guanine and cytosine, GC)含量40%~60%,退火温度50~60 ℃,且上下游引物熔解温度差值不超过5 ℃,引物长度18~23 bp,预期扩增产物长度范围为100~300 bp获得300对EST-SSR引物。所有引物由金斯瑞生物科技有限公司合成。
1.3 SSR扩增及多态性检测
从36份DNA样品中随机选取6份对300对SSR引物进行筛选,经PCR扩增和电泳分离鉴定后最终获得54对条带清晰、多态性良好的有效性引物用于进一步遗传多样性研究(表2)。25 μL PCR扩增体系包括2 μL模板DNA,0.5 μL Taq DNA 聚合酶 (2.5 U·μL-1),12 μL 2×Taq PCR Master Mix,上下游引物各1 μL,ddH2O 8.5 μL。扩增程序为94 ℃预变性4 min,接下来94 ℃变性 45 s,58 ℃退火45 s,72 ℃ 延伸1 min,共30个循环,最后72 ℃延伸10 min。PCR产物在200 V下经6%聚丙烯酰氨凝胶电泳预分离30 min后再转入350 V下电泳1.5 h。并用0.1% AgNO3进行银染和50 bp marker 鉴定PCR产物大小,显影完后用凝胶成像系统拍照保存。
1.4 数据统计与分析
根据PCR产物在凝胶电泳上的情况,选取清晰可辨的条带进行统计,对相同迁移位置或长度一致的条带有带记为“1”,无带记为“0”,构成标准的0/1矩阵,统计PCR产物扩增总带数(total number of bands,TNB)和多态性条带数(number of polymorphic bands,NPB),计算多态位点百分率(percentage of polymorphic bands,PPB)和引物多态信息含量(polymorphic information content,PIC)。其中,PPB=K/N×100%,式中:K为多态位点数目;N为所测位点总数;PICi=1-∑Pij2,式中:PICi表示引物i的多态性信息含量,Pij表示引物i的第j个带型出现的频率,引物i的总带型从 1 到n。利用NTSYS-pc2.10e软件对原始矩阵进行分析,基于similarity程序计算遗传相似系数(GS),用clustering程序中的SHAN进行聚类分析,再根据Graphics程序中的tree plot绘制亲缘关系树形图。
2 结果与分析
2.1 苏丹草转录组SSR分布频率
从苏丹草转录组的80686条Unigene序列中共鉴定出17548个SSR位点,分布在13574条Unigene序列中,发生频率为16.82%(含有SSR的Unigene 数量占总Unigene 数量的百分比),其中2965条Unigene序列含一个以上SSR位点,SSR分布频率为21.75%(SSR 的个数与总Unigene 的数量比),平均每43.14 kb出现一个SSR位点(SSR 的个数占序列总长的百分比)。

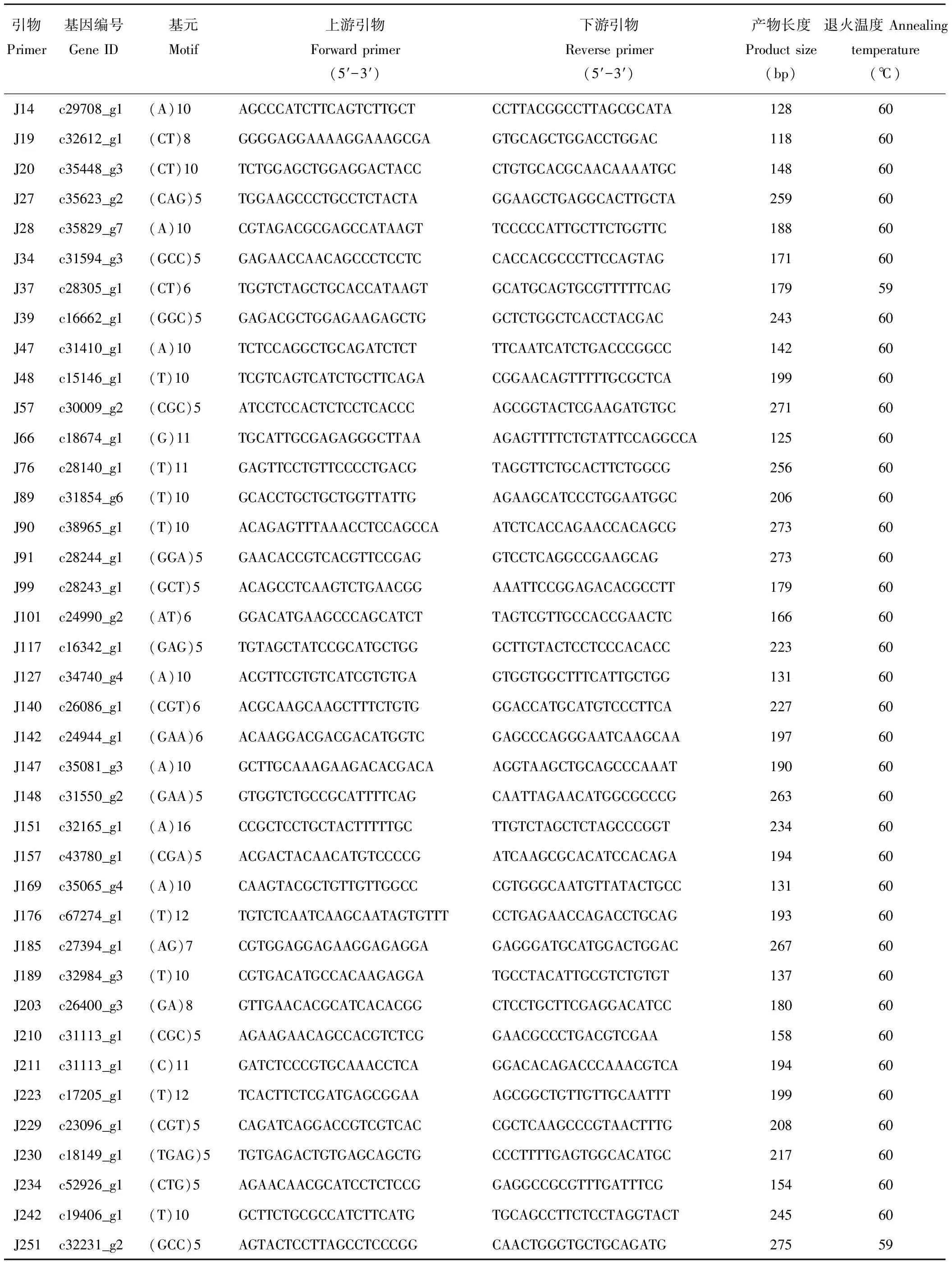
表2 引物信息及序列Table 2 Primer and sequences information applied in this study
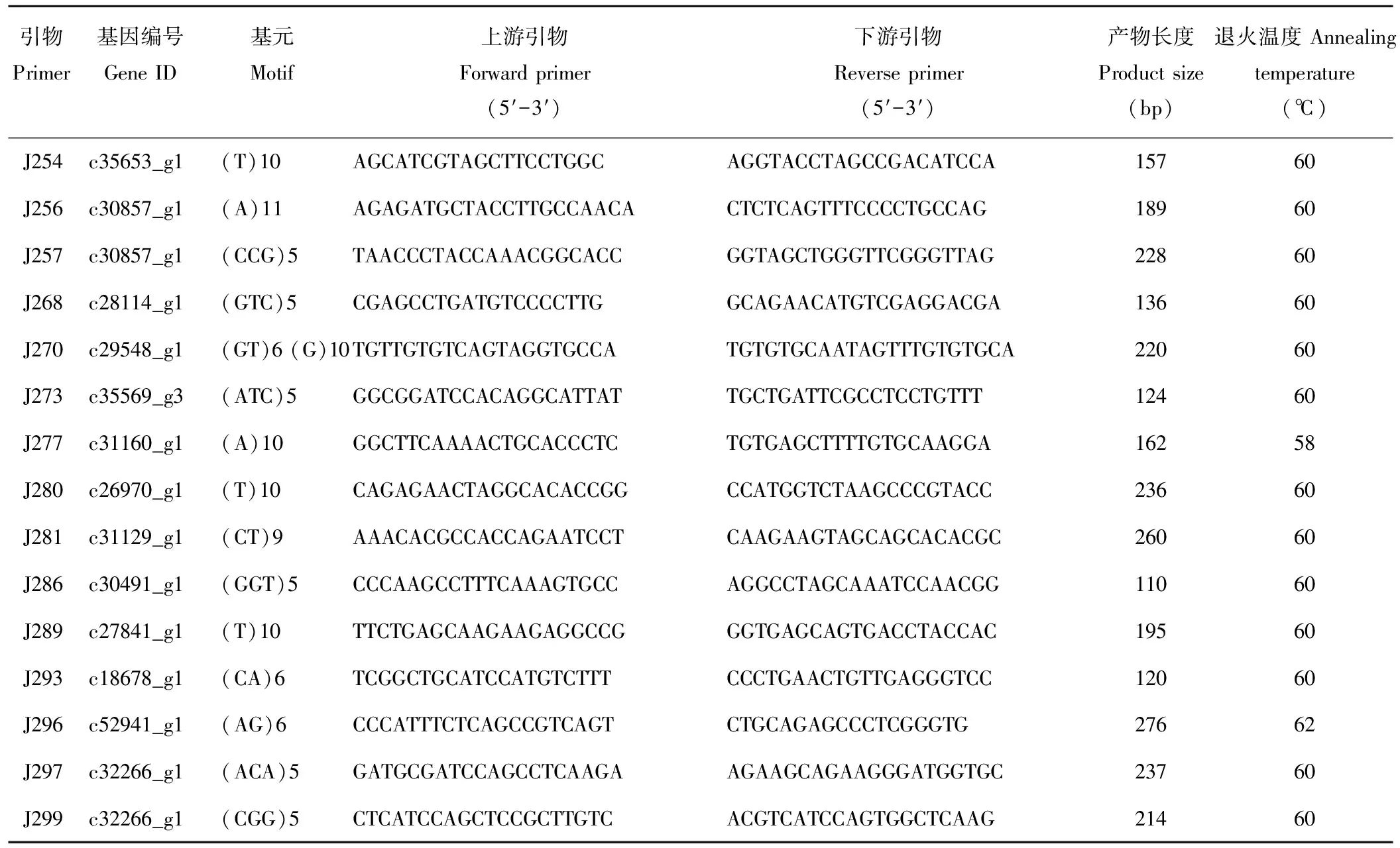
续表2 Continued Table 2
苏丹草转录组SSR种类包括一至六核苷酸6种重复类型,以单/一核苷酸重复和三核苷酸重复类型最为丰富,分别占38.59%(6771)和39.09%(6860),其次为二核苷酸重复,有3366个,占19.18%,四、五和六核苷酸重复远低于前3种重复类型,总计占3.14%(551)(表3)。
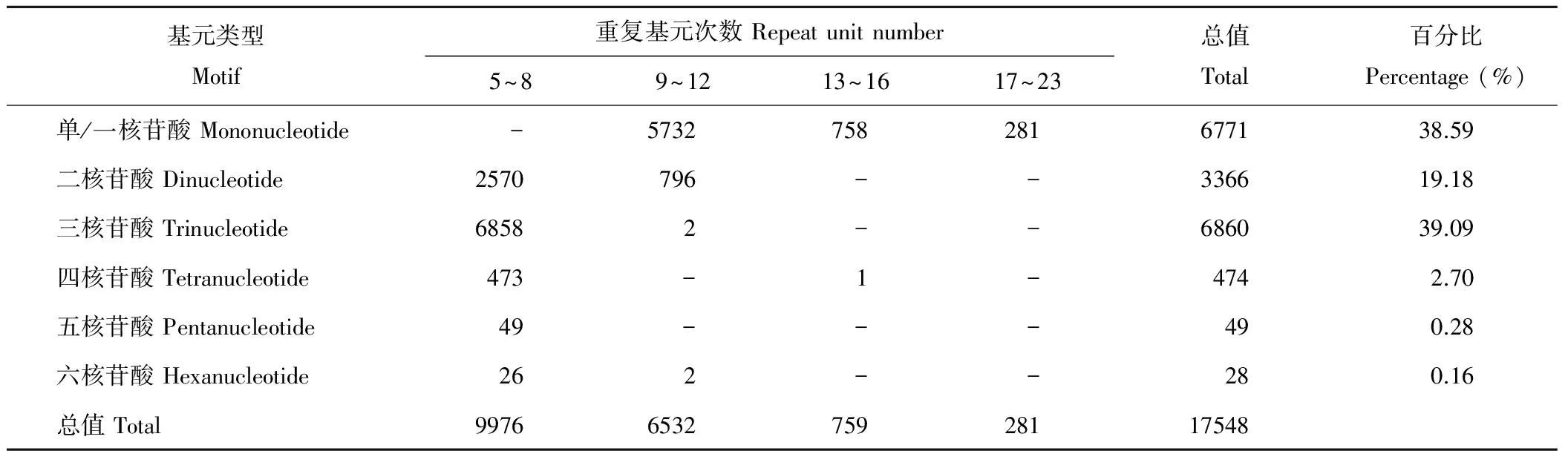
表3 基于重复基元的重复次数与重复数量分布Table 3 Frequencies of different SSR repeat motif types related to variation of repeat unit numbers
2.2 苏丹草转录组SSR位点的分布特点
苏丹草SSR重复单元的重复次数分布在5~23次,其中5~8次重复的有9976个SSR,其次为9~12次重复,有6532个SSR,往后逐渐减少。单核苷酸重复基元的重复次数跨度最大,在9~23次均有分布,其余核苷酸重复基元的重复次数主要集中于5~8次,在13~23次几乎没有分布(表3)。
17548个SSR位点中,占优势重复的一、二和三核苷酸重复基元分别有2、4和10种,其中以A/T、AG/CT和CCG/CGG出现的频率最高,分别占该重复基元类型的91.06%、46.67%和39.21%(表4)。
2.3 苏丹草转录组SSR引物筛选与多态性检测
随机选择的300对引物经初步筛选有77对引物未能扩增出清晰的条带,扩增成功率为73.67%。对扩增成功的223对引物做进一步的筛选验证,得到54对(24.22%)在供试材料间表现出扩增效果好、条带清晰且在目标条带范围内的多态性引物,如图1所示。54对有效引物在36份供试材料中共扩增出275条清晰带,平均每对引物扩增5.09条带,单个引物对最多扩增条带数为12,最少扩增条带数仅为2,扩增片段大小介于110~273 bp,其中多态性条带数有205条,多态位点百分率(PPB)平均值为73.02%,不同引物所揭示的供试材料的多态信息含量(PIC)介于0.10~0.94,平均值为0.41(表5)。
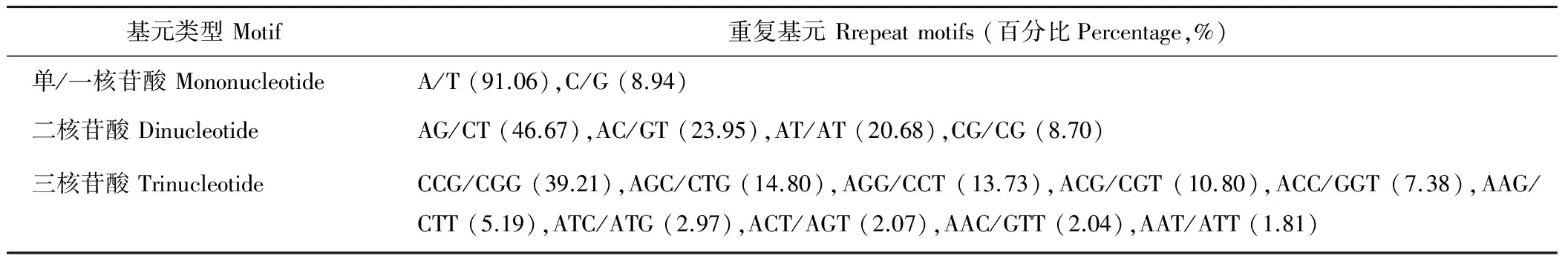
表4 重复基元类型的重复基元分布Table 4 Distribution of repeat motifs according to classified repeat types
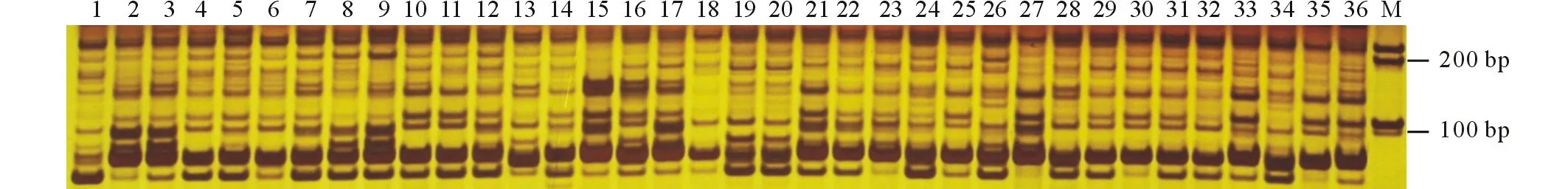
图1 引物J268对36份材料的扩增结果Fig.1 The amplification results by primer J268 on 36 accessions
2.4 基于苏丹草转录组SSR 的聚类分析
根据Nei[22]的方法用NTSYS-pc2.10e软件统计分析36份供试苏、高丹草材料间的遗传相似系数(GS),并根据遗传相似系数进行聚类分析。36份供试材料的GS介于0.3922~0.9020,平均为0.6471。其中,遗传相似系数最大的是29和30与29和31,达到0.9020,其次为30和31,29和32,31和32及27和35,达到0.8922,表明29、30、31和32这4份材料间及27和35这两份材料间的亲缘关系很接近。
由聚类结果(图2)可知,在相似系数为0.61,将36份供试材料分为3大类群:1、19和20号归为一类,1号为来源于甘肃的引进品种,19和20号均为两份栽培品种,分别来源于江苏和辽宁;27和35号两份高丹草材料以及来源于内蒙古的乌拉特2号共3份材料归为第2类群;剩余30份材料归为第3类,这一类群在相似系数为0.69处又可归为两个亚类,其来源地为川渝地区的24、26和34号以及内蒙古的28号均为引进品种的4份材料归为第一亚类,其余26份材料归为另一亚类,根据种子库提供的已有信息,该类材料大部分皆来源于甘肃、宁夏、内蒙古和四川地区。由于先前部分种子入库时的登记信息不够完善,且个别材料如登记为乌拉特2号的13号材料登记信息有待进一步的核实。针对这一情况,可通过田间试验结合表型调查、抗性鉴定等来进一步佐证。
3 讨论
转录组测序技术具有不依赖于物种全基因组信息就能快速、准确的获得全转录本序列信息的特点,近年来已成为各研究领域一种较为基础的研究手段。本研究基于苏丹草转录组测序结果分析可知,SSR位点广泛分布于苏丹草RNA序列中,占检测序列的16.82%,与鸭茅(16.43%)[23]、党参(Codonopsispilosula)(16.1%)[24]、芒草(14.44%)[20]等的测序结果基本一致。苏丹草转录组SSR种类较为丰富,一至六核甘酸重复类型均有出现,单核苷酸重复仅次于三核甘酸,其次为二核苷酸重复,其他重复类型所占比例很小,这一分布规律在高粱EST-SSR中也有体现[13]。核苷酸的重复类型因植物种类的不同而存在一定的差异,主要体现在以二核苷酸和三核苷酸重复为主导所占比例的差别上。本研究苏丹草中三核苷酸重复所占比例最高,与Yates等[25]、Huang等[26]和Cardoso-Silva等[27]分别在红三叶(Trifoliumpratense)、牛鞭草(Hemarthria)和甘蔗(Saccharumofficinarum)中的研究结果相一致。 由于密码子是以三核苷酸为一个功能单位, 在一个DNA的阅读框内三核苷酸发生迁移突变对氨基酸聚合物形成的蛋白质不会造成太大的影响[28],这可能合理的解释了为何大多数植物中三核苷酸为最为丰富的重复类型的现象。在大多数的单子叶植物中,AG/CT和CCG/CGG分别为二核苷酸和三核苷酸的优势重复基元[29],本研究结果也刚好印证了这一点。苏丹草转录组SSR标记中单核苷酸和三核苷酸SSR重复基元的重复次数多,尤其以单核苷酸重复基元的跨度最大,在9~23次均有分布。研究表明,核苷酸的重复次数与SSR标记的多态性呈正相关[30],所以理论上这两种核苷酸SSR具备更为丰富的多态性,可作为潜在的SSR重复基元进行引物设计和分子标记开发。
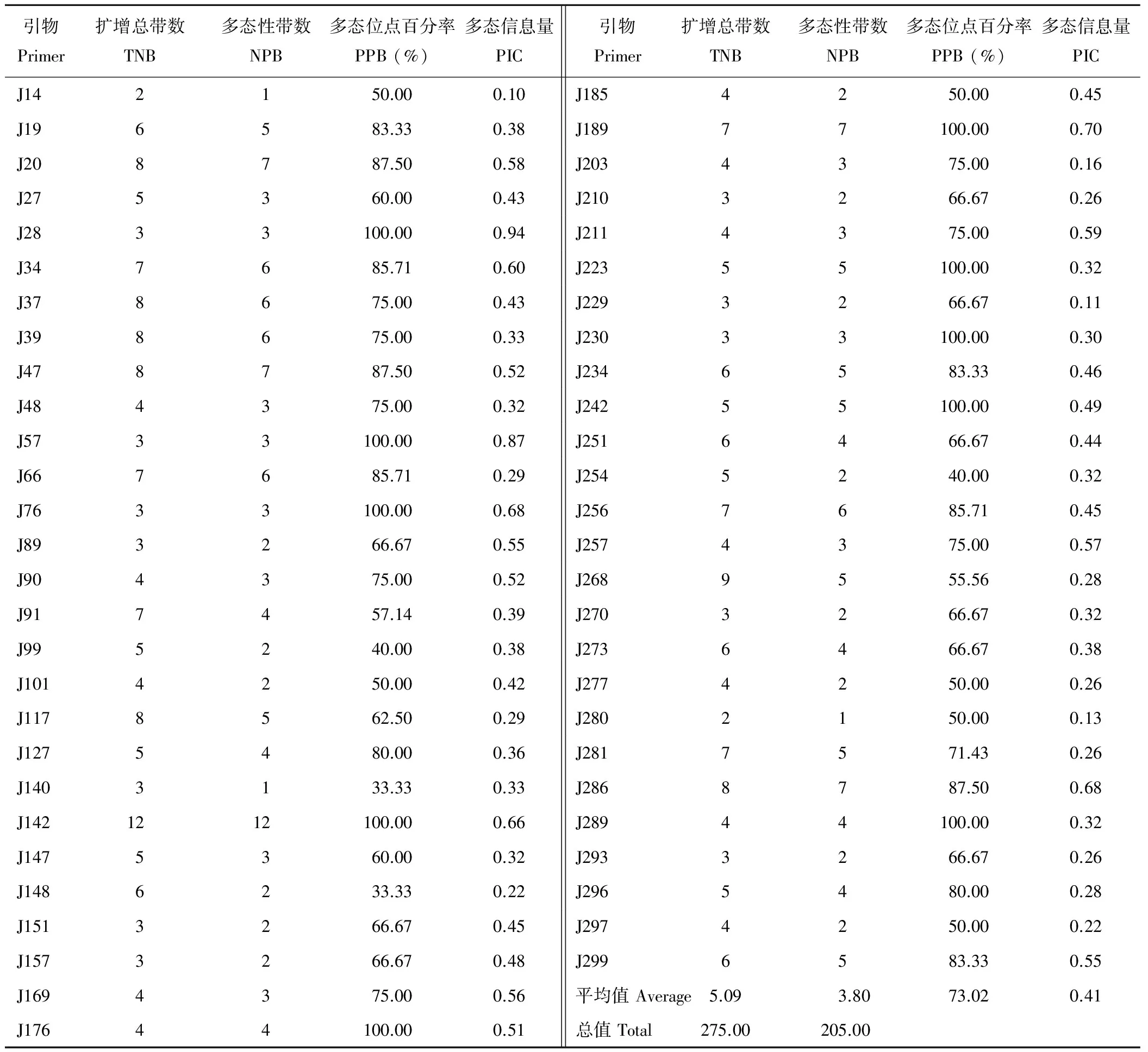
表5 54对引物扩增结果Table 5 The amplification results of 54 SSR primer pairs
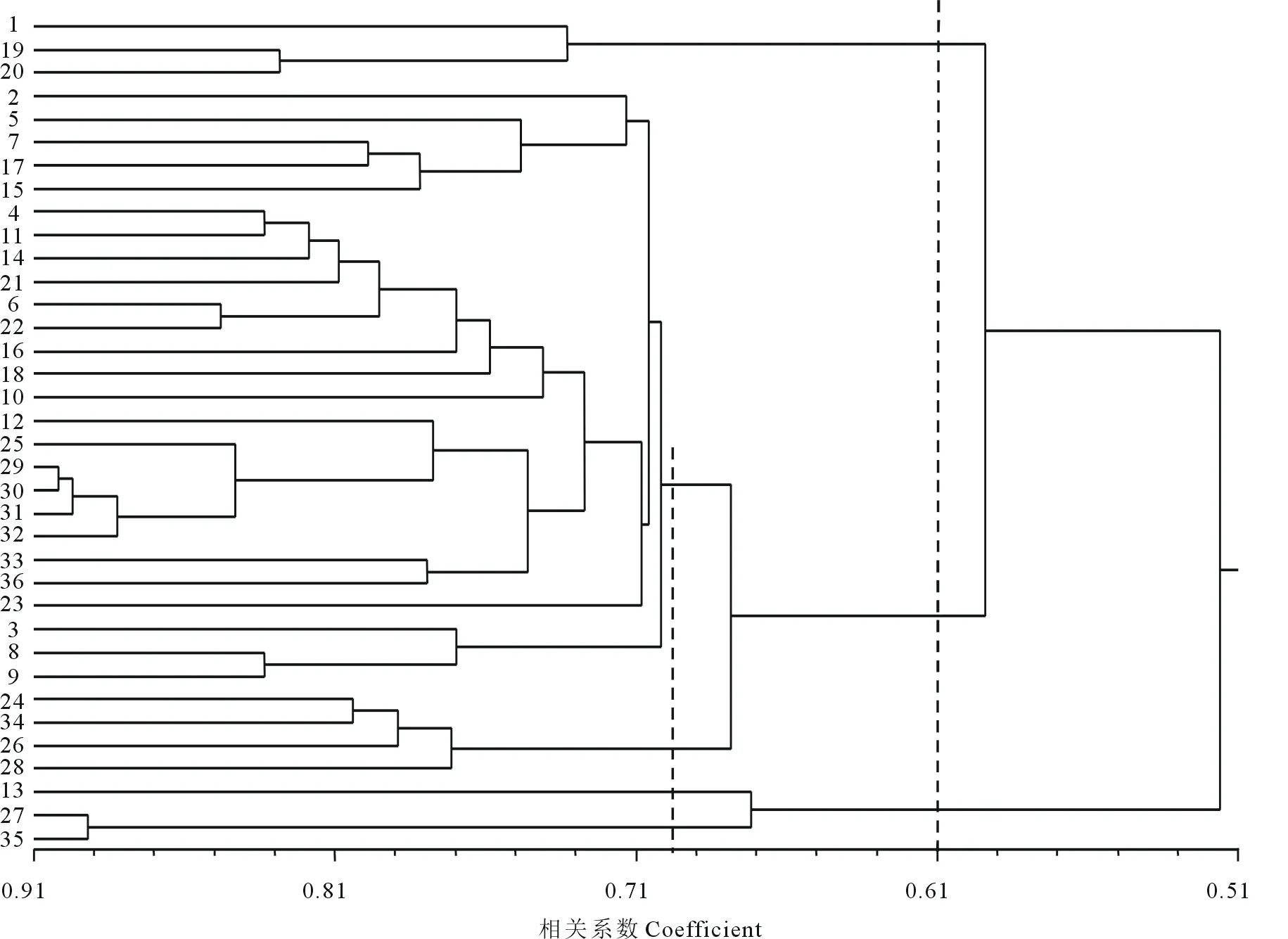
图2 EST-SSR对36份材料的UPGMA聚类分析Fig.2 UPGMA cluster of 36 accessions based on EST-SSR markers
根据EST-SSR是对基因内部变异进行直接评价的这一特点,本研究利用包含不同重复基元的54对多态性引物应用于34份苏丹草和2份高丹草材料中进行分子标记扩增分析,得到较多的变异位点,其中73.05%的扩增片段能够揭示材料间的遗传差异,均值为0.41的多态信息含量(PIC)反映了供试材料间的遗传多样性水平。一般来说,PIC含量越高,表明群体变异程度越大,物种的遗传多样性越丰富[31]。这说明基于转录组开发的EST-SSR对苏丹草遗传多样性的检测具有一定的有效性,同时也证明了供试材料具有较为丰富的遗传变异类型。
基于遗传相似系数(GS)的分析也能印证以上结论。36份供试材料的平均GS为0.6471,表明供试材料间具有较远的亲缘关系,而27和35号两份高丹草材料其GS为0.8922,聚类结果也归为一类,表明苏丹草转录组开发的SSR引物扩增出的特异位点能够有效地将苏丹草与高丹草材料进行区别。另外,采自于川渝地区的29、30、31和32号4份材料间的GS值较高,且其聚类结果与地理来源基本一致,说明来自于同一地区的材料遗传变异较小。以上结果均表明根据苏丹草转录组开发的SSR标记对苏、高丹草遗传变异和亲缘关系的鉴别能力具有一定的可行性,可有效应用于苏、高丹草种质资源的遗传多样性评价中。
根据EST-SSR标记具有很好的通用性这一特点,EST-SSR引物不仅可运用于近源物种的遗传多样性研究,如将高粱EST-SSR标记应用于苏丹草及高丹草[13],将高丹草EST-SSR标记应用于苏丹草和高粱[31]等。此外,陈永霞等[32]还将玉米(Zeamays)、水稻(Oryzasativa)和高粱3种禾谷类作物的EST-SSR引物成功用于扁穗牛鞭草(Hemarthriacompressa)的跨种扩增研究其遗传多样性水平。本研究根据苏丹草转录组数据开发大量本草种EST-SSR分子标记,通过遗传多样性分析揭示了所开发的引物具有良好的特异性,在这一基础上,以后可将这54对特异性SSR引物运用到高粱、高丹草以及其他禾本科牧草、草坪草甚至是禾谷类作物等的遗传多样性研究工作中。由于EST-SSR来源于序列相对保守的基因组编码区,相同引物因与不同材料的基因组互补DNA片段数目和位点而不同,扩增产物大小和数目也存在一定差异,故扩增产物的多态性反映了材料的遗传多样性,为基因功能多样性的研究提供了可能[33]。因此,利用EST-SSR标记技术对草种质资源进行遗传多样性分析,可以更好地开发和利用优良基因资源,保护和鉴定基因的多样性,并可通过对材料的亲缘关系进行梳理[34],为杂交育种选育新品系奠定基础,提高品种选育工作的准确性。
4 结论
本研究基于苏丹草转录组测序分析明确了苏丹草SSR位点分布的总体特征,利用SSR位点数目和种类均较为丰富的EST序列开发了54个特异的苏丹草SSR分子标记,建立了高效开发苏丹草特异标记的新途径,为今后苏丹草及近源物种遗传组成的鉴定、遗传多样性研究、“三性”测试(DUS)和分子标记辅助育种奠定了重要基础。
参考文献References:
[1] Zhang X D, Liu J R, Ye T Y. The growth performance and cultivation techniques ofSorghumsudanense. Bulletin of Agricultural Science and Technology, 2004, (2): 38-39.
张希弟, 刘建荣, 叶挺云. 苏丹草的生长表现及栽培技术. 农业科技通讯, 2004, (2): 38-39.
[2] Ke M, Zhu H, Liang W W,etal. Grey correlation analysis of main agronomic characters and its yield and quality traits inSorghumsudanense. Pratacultural Science, 2016, 33(5): 949-955.
柯梅, 朱昊, 梁维维, 等. 苏丹草农艺性状与产量、品质间的灰色关联度分析. 草业科学, 2016, 33(5): 949-955.
[3] Xu Y P, Wu Z X, Zhao Z X. The adaptability and the developing foreground of Sudan grass in the produce of agriculture and animal husbandry in China. Pratacultural Science, 2003, (7): 23-25.
徐玉鹏, 武之新, 赵忠祥. 苏丹草的适应性及在我国农牧业生产中的发展前景. 草业科学, 2003, (7): 23-25.
[4] Li C J, Fu Y B, Wan J C,etal. Genetic diversity of agronomic characteristics of 30Sorghumsudanensegermplasm. Pratacultural Science, 2015, 32(1): 85-93.
李陈建, 付彦博, 万江春, 等. 30份苏丹草种质资源农艺性状的遗传多样性分析. 草业科学, 2015, 32(1): 85-93.
[5] Zhan Q W. Prospect analysis of Sorghum-sudangrass hybrids in bio-energy utilization. Seed, 2009, 28(8): 77-79.
詹秋文. 高粱-苏丹草杂交种生物能源利用的前景分析. 种子, 2009, 28(8): 77-79.
[6] Wang J, Gao Q, Yang G F,etal. Fingerprint constructing and genetic diversity analyzing ofSorghumsudanenseandSorghumbicolor×Sorghumsudanensewith SSR markers. Acta Agrestia Sinica, 2016, 24(1): 156-164.
王杰, 高秋, 杨国锋, 等. 国审苏丹草和高丹草品种SSR指纹图谱构建及遗传多样性分析. 草地学报, 2016, 24(1): 156-164.
[7] Wang Y W, Cao Z Z, Han J G,etal. Evaluation and cluster analysis on forage and seed performances of nineSorghumsudanensevarieties. Acta Pratacultural Sinica, 2005, (5): 117-123.
王赟文, 曹致中, 韩建国, 等. 9个苏丹草品种生产性能的评价与聚类分析. 草业学报, 2005, (5): 117-123.
[8] Kalia R K, Rai M K, Kalia S,etal. Microsatellite markers: an overview of the recent progress in plants. Euphytica, 2011, 177(3): 309-334.
[9] Agarwal M, Shrivastava N, Padh H. Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Reports, 2008, 27(4): 617-631.
[10] Luo R, Wu W L, Zhang Y,etal. SSR marker and its application to crop genetics and breeding. Genomics and Applied Biology, 2010, 29(1): 137-143.
罗冉, 吴委林, 张旸, 等. SSR分子标记在作物遗传育种中的应用. 基因组学与应用生物学, 2010, 29(1): 137-143.
[11] Zhan Q W, Zhang T Z, Wang B H,etal. Diversity comparison and phylogenetic relationships ofS.bicolorandS.sudanenseas revealed by SSR markers. Plant Science, 2008, 174(1): 9-16.
[12] Lu J, Zhan Q W, Lu J B,etal. Development of EST-SSR maker in Sorghum and Sudangrass. Acta Laser Biology Sinica, 2009, 18(3): 389-394.
卢杰, 詹秋文, 陆景标, 等. 高粱与苏丹草EST-SSR标记的开发. 激光生物学报, 2009, 18(3): 389-394.
[13] Li J Q, Wang L H, Zhan Q W,etal. Establishment of sorghum EST-SSR marker and its preliminary application to sudangrass. Pratacultural Science, 2010, 27(3): 112-117.
李杰勤, 王丽华, 詹秋文, 等. 高粱EST-SSR标记的建立及其在苏丹草中的应用初探. 草业科学, 2010, 27(3): 112-117.
[14] Kane N C, Rieseberg L H. Selective sweeps reveal candidate genes for adaptation to drought and salt tolerance in common sunflower,Helianthusannuus. Genetics, 2007, 175(4): 1823-1834.
[15] Yu X Q, Bai G H, Luo N,etal. Association of simple sequence repeat (SSR) markers with submergence tolerance in diverse populations of perennial ryegrass. Plant Science, 2011, 180(2): 391-398.
[16] Nie G, Huang L K, Zhang X Q,etal. Marker-trait association for biomass yield of potential bio-fuel feedstockMiscanthussinensisfrom Southwest China. Frontiers in Plant Science, 2016, 7: 802.
[17] Huang H Y, Du H Y, Wu Y T N,etal. Development of SSR molecular markers based on transcriptome sequencing ofEucommiaulmoides. Scientia Silvae Sinicae, 2013, 49(5): 176-181.
黄海燕, 杜红岩, 乌云塔娜, 等. 基于杜仲转录组序列的SSR分子标记的开发. 林业科学, 2013, 49(5): 176-181.
[18] Liu Z P, Chen T L, Ma L C,etal. Global transcriptome sequencing using the Illumina platform and the development of EST-SSR markers in autotetraploid alfalfa. PLoS One, 2013, 8(12): e83549.
[19] Huang L K, Yan H D, Zhao X X,etal. Identifying differentially expressed genes under heat stress and developing molecular markers in orchardgrass (DactylisglomerataL.) through transcriptome analysis. Molecular Ecology Resources, 2015, 15(6): 1497-1509.
[20] Nie G, Tang L, Zhang Y J,etal. Development of SSR markers based on transcriptome sequencing and association analysis with drought tolerance in perennial grassMiscanthusfrom China. Frontiers in Plant Science, 2017, 8: 801.
[21] Li J Q, Wang L H, Zhan Q W,etal. Transcriptome characterization and functional marker development inSorghumsudanense. PloS One, 2016, 11(5): 1-12.
[22] Nei M. Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences, 1973, 70(12): 3321-3323.
[23] Li J, Huang L K, Jin M Y,etal. Development and verification of orchardgrass Genomic-SSR. Molecular Plant Breeding, 2017, (10): 4071-4079.
李季, 黄琳凯, 金梦雅, 等. 鸭茅基因组Genomic-SSR标记开发. 分子植物育种, 2017, (10): 4071-4079.
[24] Wang D, Cao L Y, Gao J P. Data mining of simple sequence repeats inCodonopsispilosulatranscriptome. Chinese Traditional and Herbal Drugs, 2014, 45(16): 2390-2394.
王东, 曹玲亚, 高建平. 党参转录组中SSR位点信息分析. 中草药, 2014, 45(16): 2390-2394.
[25] Yates S A, Swain M T, Hegarty M J,etal.Denovoassembly of red clover transcriptome based on RNA-Seq data provides insight into drought response, gene discovery and marker identification. BMC Genomics, 2014, 15(1): 453.
[26] Huang X, Yan H D, Zhang X Q,etal.Denovotranscriptome analysis and molecular marker development of twoHemarthriaspecies. Frontiers in Plant Science, 2016, 7: 496.
[27] Cardoso-Silva C B, Costa E A, Mancini M C,etal. De novo assembly and transcriptome analysis of contrasting sugarcane varieties. PloS One, 2014, 9(2): e88462.
[28] Kaur S, Pembleton L W, Cogan N O I,etal. Transcriptome sequencing of field pea and faba bean for discovery and validation of SSR genetic markers. BMC Genomics, 2012, 13(1): 104.
[29] Kantety R V, La Rota M, Matthews D E,etal. Data mining for simple sequence repeats in expressed sequence tags from barley, maize, rice, sorghum and wheat. Plant Molecular Biology, 2002, 48(5): 501-510.
[30] Gao L F, Tang J F, Li H W,etal. Analysis of microsatellites in major crops assessed by computational and experimental approaches. Molecular Breeding, 2003, 12(3): 245-261.
[31] Wen Y, Lu X P, Ren R,etal. Development of EST-SSR marker and genetic diversity analysis inSorghumbicolor×Sorghumsudanenes. Hereditas, 2013, 35(2): 225-232.
温莹, 逯晓萍, 任锐, 等. 高丹草EST-SSR标记的开发及其遗传多样性. 遗传, 2013, 35(2): 225-232.
[32] Chen Y X, Zhang X Q, Xie W G,etal. Genetic diversity ofHemarthriacompressagermplasms from southwestern China based on EST-SSR markers. Acta Prataculturae Sinica, 2011, 20(6): 245-253.
陈永霞, 张新全, 谢文刚, 等. 利用EST-SSR标记分析西南扁穗牛鞭草种质的遗传多样性. 草业学报, 2011, 20(6): 245-253.
[33] Wang R J, Li P Y, Zhang Y H. Development ofElytrigiarepensEST-SSR marker and its application. Pratacultural Science, 2016, 33(8): 1526-1535.
王瑞晶, 李培英, 张延辉. 偃麦草EST-SSR标记开发及应用. 草业科学, 2016, 33(8): 1526-1535.
[34] Wang F, Gao Q, Wang J,etal. Analysis of genetic diversity inSorghumgermplasm collections using SSR markers. Acta Prataculturae Sinica, 2016, 25(5): 125-133.
王芳, 高秋, 王杰, 等. 利用SSR标记分析高粱属种质资源的遗传多样性. 草业学报, 2016, 25(5): 125-133.
DOI:10.11686/cyxb2017262http://cyxb.lzu.edu.cn
成启明, 格根图, 尹强, 等. 苜蓿干草捆安全贮藏条件的研究. 草业学报, 2018, 27(5): 190-200.
Cheng Q M, Ge G T, Yin Q,etal. Effect of moisture, bale density and CaO on storage life and quality of alfalfa hay. Acta Prataculturae Sinica, 2018, 27(5): 190-200.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
昆明医科大学学报(2021年3期)2021-07-22
趣味(语文)(2019年10期)2020-01-14
环球时报(2019-04-12)2019-04-12
环球人物(2018年7期)2018-04-23
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
癌变·畸变·突变(2014年2期)2014-03-01
中国神经精神疾病杂志(2013年4期)2013-03-11
