
κ阿片受体在抑郁中的作用及机制研究进展
2018-06-19 09:32昝桂影李庆林刘景根
神经药理学报 2018年1期
昝桂影 孙 翔 李庆林 刘景根
1. 中国科学院上海药物研究所,上海,201203,中国
2. 安徽中医药大学,合肥,230000,中国
世界卫生组织数据显示,西方人一生罹患抑郁症概率高达16.9%,抑郁症每年消耗欧洲国家1%的国内生产总值[1]。我国约有超过50%的人处于心理亚健康状态[2]。流行病学调查结果显示,我国15岁以上人口中,各类精神疾病患者人数超过1亿人,其中多数是抑郁症、自闭症等精神障碍或心理行为障碍患者[3],给我国人民的身体健康和经济发展带来了沉重的负担。由于抑郁症发病的复杂性,现在仍然不清楚抑郁发病的机制,探索抑郁发病的机制进而研发有效的治疗药物已经成为当下急需解决的问题。
内源性阿片系统是参与应激之后情感和行为反应的重要媒介,主要包括内啡肽、脑啡肽和强啡肽及其受体μ,δ和κ阿片受体。激动μ阿片受体和δ阿片受体能够提升情绪,激动κ阿片受体却会引起焦躁不安和抑郁样行为等负性情绪[4-5]。当接受持续性的神经信号时,强啡肽由致密突触囊泡释放,作用于κ阿片受体。κ阿片受体激活产生负性情绪与中枢神经系统相关,中枢神经系统κ激活参与介导应激和抑郁[6]。强啡肽可以调节神经元兴奋,影响学习、认知,伤害感受和内分泌功能[7-8]。当给予κ阿片受体拮抗剂或者当κ阿片受体和强啡肽原敲除时,应激引起的强迫游泳不动时间升高可以被阻断[9-10],提示强啡肽/κ阿片受体介导应激引起的抑郁样行为。这些抗抑郁行为在重复的应激刺激中尤其明显,说明κ 阿片受体介导应激行为的放大和敏化。
1 强啡肽/κ阿片受体系统和应激
κ阿片受体是一种G蛋白耦联受体,主要通过抑制腺苷酸环化酶及电压门控钙通道,激活电压门控钾通道降低突触传递。κ阿片受体主要存在三种亚型(κ1、κ2、κ3),广泛分布于大脑、脊髓和痛觉神经元[11]。脑室和系统给予特异性κ阿片受体激动剂能够产生良好的镇痛效果,提示激活中枢κ阿片受体可以产生镇痛作用。遗憾的是,κ阿片受体发挥镇痛作用的具体部位目前还没有细致的研究。早在1962年,我国药理学家邹冈和张昌绍先生便发现中央导水管灰质是发挥阿片类物质镇痛作用的重要脑区[12],Wang等发现κ阿片受体在中脑导水管周围灰质区(periaqueductal gray,PAG)大量表达[11],因此,推测κ阿片受体也有可能通过PAG脑区发挥镇痛作用。揭示κ阿片受体发挥镇痛作用的脑区,从而研发该脑区特异性激活κ阿片受体的药物,为将来开发低副作用的镇痛药物提供思路。
临床研究发现,应激是导致抑郁的重要因素[13],急性和慢性应激可以激活下丘脑-垂体-肾上腺(hypothalamic-pituitary-adrenal,HPA)轴,促 进 下 丘脑室旁核的神经元分泌促肾上腺皮质激素释放因子(corticotrophin releasing factor,CRF),CRF 促 进 垂 体前叶促肾上腺皮质激素(adrenocorticotropic-hormone,ACTH)释放,促进肾上腺皮质释放糖皮质激素,进而促进神经肽在周围和中枢神经系统的释放。强啡肽信号通路和HPA轴在应激中存在相互协同作用,CRF和持续的应激环境引起κ阿片受体在背侧中缝核(dorsal raphe nucleus,DRN),杏 仁 核(amygdala),海 马(hippocampus),腹 侧 被 盖 区(ventral tegmental area,VTA)和伏隔核(nucleus accumbens,NAc)的激活。Land等发现强啡肽基因敲除或者κ阿片受体拮抗可以阻断CRF引起的烦躁不安,提示κ阿片受体参与调节应激[14]。应激和κ阿片受体激动时,CRF引起的κ阿片受体磷酸化和谷氨酸脱羧酶(glutamic acid decarboxylase,GAD),胶质纤维酸性蛋白(glial fibrillary acidic portein,GFAP) 和 色 氨 酸 羟 化 酶(tryptophan hydroxylase,TPH)产生共定位[14-15],说明κ阿片受体可能参与调节γ-氨基丁酸(γ-aminobutyric acid,GABA)能神经元,星形胶质细胞和5-羟色胺(5-hydroxytryptamine,5-HT)能神经元的信号传导。
Reyes等发现杏仁核脑区CRF和强啡肽可以共定位,提示CRF可能调控强啡肽的表达[16]。Bruchas 等发现侧脑室给予CRF可以降低实验动物在高架十字迷宫开放臂的时间,并能引起实验动物杏仁核基底外侧核(basal nucleus of the amygdala,BLA)脑区κ阿片受体激活,脑室给予CRF受体1(corticotropin-releasing factor receptor 1,CRF-R1)拮抗剂可以阻断 CRF 引起的焦虑行为,而CRF受体2(corticotropin-releasing factor receptor 2,CRF-R2)拮抗剂则不能,BLA 脑区给予Norbinaltorphimine(norBNI)可以阻断CRF引起的焦虑行为,提示CRF通过激活CRF-R1激活BLA脑区κ阿片受体从而引起焦虑行为[17]。
除 CRF-R1受体外,CRF-R2对强啡肽 /κ阿 片受体系统也存在调控作用[18],杏仁中央核(central amygdaloid nucleus,CeA) CRF受体激活和急性酒精摄入可以增加CeA脑区强啡肽水平,且可以被CRF-R2拮抗剂阻断,CRF-R1受体则不能[19]。Land等发现脑室给予CRF引起的条件位置厌恶可以被κ阿片受体拮抗所阻断,CRF-R2拮抗剂可以阻断CRF引起的条件位置厌恶,而CRF-R1阻断则不能,CRF-R1敲除的实验动物不能产生U50,488H引起的条件位置厌恶[14]。
GABA是哺乳动物中枢神经系统主要的抑制性神经递质,GABA 受体兴奋剂通常具备抗焦虑作用[20]。Kang-Park 等通过脑片膜片钳发现,敲除κ阿片受体或者给予κ阿片受体拮抗剂norBNI可以强化CRF对GABA能神经元抑制性微突触后电流(miniature inhibitory postsynaptic potential,mIPSC)的增强效果,CRF-R1拮抗剂可以阻断norBNI对mIPSC的增强效果,而CRF-R2不能阻断[21]。由这些研究,推测κ阿片受体拮抗剂通过增强GABA能神经元的抑制性突触后传递调控焦虑行为。但是Kang-Park 等同时发现,离体脑片给予外源性CRF可以增加GABA能神经元的抑制性突触后传递,但是CRF却可以引起焦虑样行为,这和苯二氮类通过增强GABA能神经元抑制性突触后传递发挥抗焦虑作用相矛盾,这种不同可能源于CRF的剂量差异,如适当的应激刺激(CRF水平升高)可以增加动力,而较强的应激却能引起焦虑反应,以后研究可以从CRF对GABA能神经传递的作用出发来探究不同程度的应激可以引起动力增加或引起焦虑反应的机制。Bruchas 等发现应激引起κ阿片受体激活介导应激引起的厌恶情绪,应激引起的κ阿片受体磷酸化可以和GABA能神经元标记物GAD产生共标,这项研究侧面支持了κ阿片受体可以调节GABA能神经元的抑制性突触后传递[15]。
目前,CRF调控强啡肽/κ阿片受体系统的分子机制仍不清楚,CRF是直接引起强啡肽释放激活κ阿片受体还是通过中间分子进行调控的,还没有相关的文献报道。CRF受体是一种G蛋白耦联受体,通过有丝分裂原激活蛋白激酶(mitogen-activated protein kinase,MAPK)激活转录因子,进而介导相关蛋白或者多肽的表达。因此,推测CRF可能通过蛋白激酶激活环磷腺苷效应元件结合蛋白(cAMP-response element binding protein,CREB)引起强啡肽原的转录表达增加,强啡肽A表达增加,激活κ阿片受体从而引起行为变化,这个推断还需要具体的实验验证。将来的研究也可以围绕强啡肽/κ阿片受体是否能够调节CRF系统展开,探索CRF系统和强啡肽/κ阿片受体相互作用发挥机体应激反应的具体机制。
2 κ阿片受体的外源性配体调控抑郁行为研究现状
在动物行为上主要通过强迫游泳(forced swim test,FST),悬尾实验(tail suspension test,TST)及颅内自身刺激(intracranial self-stimulation,ICSS)进行实验动物抑郁行为的评价。强迫游泳和悬尾实验是利用实验动物在极端环境中停止挣扎的时间判断实验动物是否发生抑郁,实验动物的不动时间越长,抑郁水平越高。ICSS是一种操作性行为范式,在该实验中,实验动物学会提供简短的电脉冲到自己的大脑的特定奖赏区域(包括外侧下丘脑,内侧前脑,腹侧被盖区等)。在ICSS程序中,升高ICSS阈值反映了刺激对奖赏效应的减弱,即快感缺失,而降低ICSS阈值表明刺激对奖赏效应的增强。大多数滥用药物刺激可以降低ICSS阈值,而包括抑郁在内的负性情绪可以升高ICSS阈值。
目前,常用的κ阿片受体激动剂包括U50,488H、U69593和Salvinorin A等,拮 抗 剂 包 括norBNI、JDTic、GNTI和LY2456302等,κ拮抗剂相对长效。κ阿片受体外源性配体在抑郁样行为中的作用近年来被广泛研究(Tab.1)。研究发现,腹腔给予κ激动剂Salvinorin A能够增加实验动物在ICSS测试上的阈值,并且增加大鼠在强迫游泳中的不动时间[22-23]。在雄性动物中,系统给予κ阿片受体激动剂U50,488H可以剂量依赖的引起快感缺失症状,但是在雌性动物中,这一现象并不明显[24],提示κ激动剂对抑郁样行为的调控存在性别差异。另有个别研究发现激动κ阿片受体可以发挥抗抑郁作用,Harden等发现κ阿片受体激动剂Salvinorin A可以阻断慢性应激引起的快感缺失[25],王等研究发现κ阿片激动剂和μ部分激动剂ATPMET可以发挥抗抑郁和抗焦虑的作用[26-27]。这和大多数研究结果存在矛盾,需要进一步探索其机制。κ阿片受体拮抗剂可以阻断抑郁样行为,系统给予norBNI可以阻断社交应激失败引起的抑郁样行为[28]。初生小鼠暴露于利他林引起实验动物产生蔗糖偏爱减少和强迫游泳不动时间升高的抑郁样行为,伴随给予U50,488H可以加剧抑郁样行为,拮抗剂norBNI可以阻断抑郁样行为[29]。
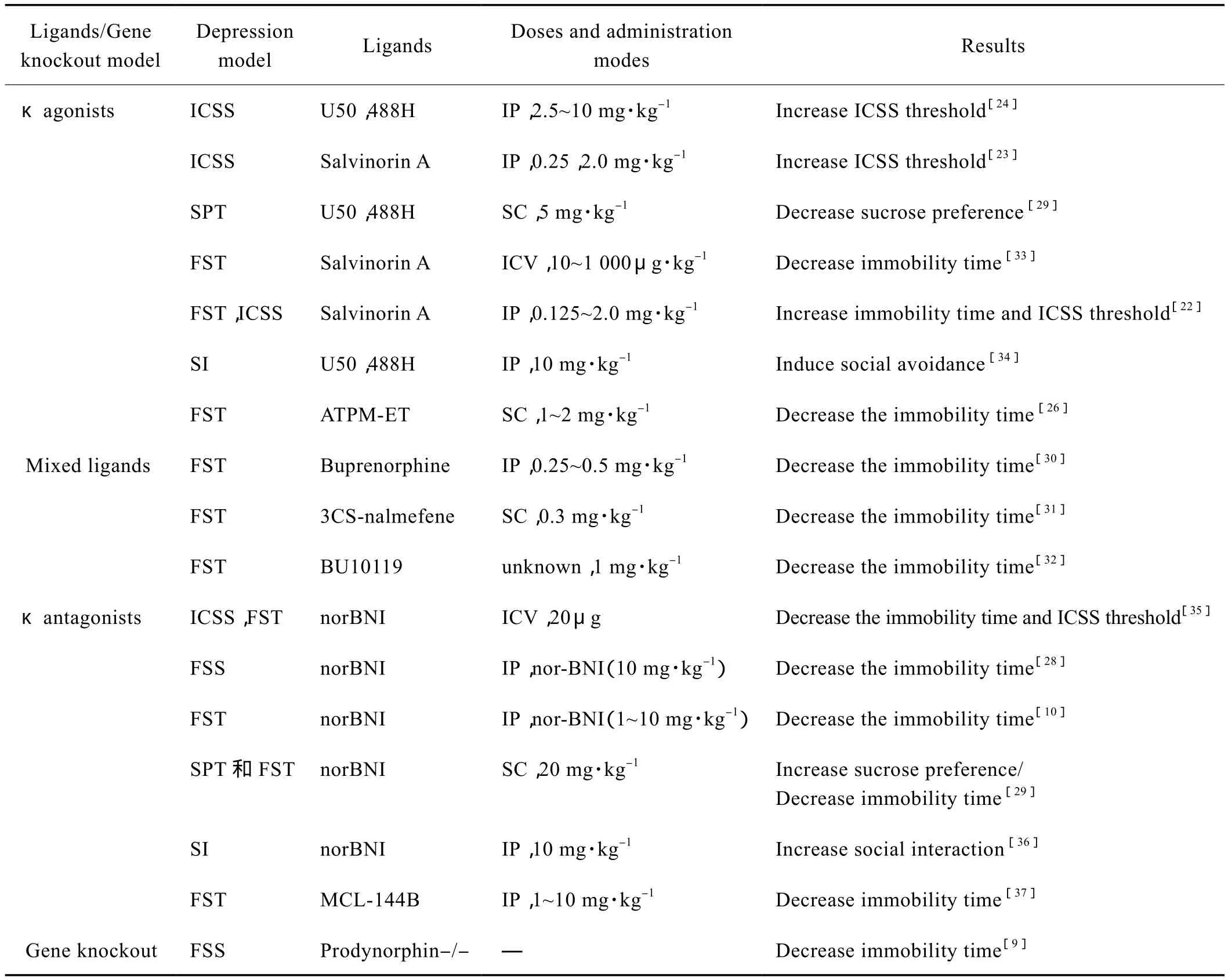
Tab.1 The role of κ ligands and gene knockout approach on animal depression model
以丁丙诺啡为代表的阿片受体部分激动剂近年来得到越来越多的关注,丁丙诺啡是一种κ阿片受体拮抗剂,μ阿片受体部分激动剂。动物实验显示,丁丙诺啡可以明显降低强迫游泳不动时间[30],临床实验显示丁丙诺啡在传统抗抑郁药物治疗无效的抑郁治疗中显示非常有希望的前景。Callaghan等发现κ阿片受体部分激动剂3CS-nalmefene可以明显阻断炎症因子引起的抑郁样行为[31]。Almatroudi等发现新型κ/μ受体混合型拮抗剂BU10119可以降低实验动物在强迫游泳实验中的不动时间[32]。这些研究说明阿片受体混合型拮抗/激动剂在抗抑郁药物的开发中相对更有优势,可以作为将来抗抑郁药物开发努力的方向。
3 κ阿片受体参与调控谷氨酸及单胺类神经递质
谷氨酸是哺乳动物中枢神经系统内重要的兴奋性神经递质,在脑组织中的浓度大约在8~10 mmol·kg-1,显著高于单胺类递质(以μmol·kg-1计量),临床研究显示,抑郁症患者中谷氨酸类神经递质传递出现异常[38]。近年来,谷氨酸递质系统成为抗精神类药物研发的重要靶点[39],以克他命为代表的N-甲基-D-天门冬氨酸(N-methyl-D-aspartate,NMDA)受体拮抗剂在治疗抑郁中疗效显著,与传统的抗抑郁药物相比,克他命发挥药效更迅速。克他命引起谷氨酸递质的大量释放,增加谷氨酸递质传递,这可能是其发挥快速抗抑郁作用的机制[40]。研究表明,强啡肽/κ 阿片受体系统参与调控谷氨酸神经递质传递,脑片电生理表明,U69593孵育伏隔核脑片可以降低该区域的谷氨酸释放[41]。Tejeda等发现内侧前额叶皮层(medial frontal cortex,mPFC)给予U69593可以抑制BLA到PFC的谷氨酸能神经投射,并且该效应可被norBNI阻断,进一步研究表明mPFC脑区给予norBNI可以发挥抗焦虑作用[42]。鉴于之前研究发现BLA脑区κ 阿片受体介导应激引起的焦虑行为,因此推测κ阿片受体可能通过影响BLA至mPFC的谷氨酸神经投射发挥对焦虑行为的作用,需要进一步研究。Crowley等发现脑片给予κ激动剂U69593可以抑制BLA至终纹床核(bed nucleus of the striatal terminalis,BNST)的兴奋性神经投射,光遗传激活κ阿片受体可以抑制谷氨酸的释放[43]。强啡肽/κ阿片受体参与谷氨酸重摄取的调控,在体外培养的海马神经元中,κ阿片受体激动剂U50,488H可以显著增加兴奋性转运蛋白3(excitatory amino acid transporter 3,EAAT3)的表达[44]。κ阿片受体和谷氨酸转运体 1(glutamate transporter 1,GLT1)在神经胶质细胞中均广泛存在,但是κ阿片受体对GLT1表达的调控目前仍不清楚,强啡肽/κ阿片受体系统对于谷氨酸递质系统的调节的行为学意义还没有得到阐明。
5-HT、多巴胺(dopamine,DA)和去甲肾上腺素(noradrenaline,NE)等单胺类递质是经典的抗抑郁药物作用的靶点。5-HT再摄取抑制剂(serotonin specific reuptake inhibitors,SSRIs)是目前抑郁症治疗的一线用药,如氟西汀、帕罗西汀等。SSRIs通过抑制突触前膜对5-HT的再摄取,提高突触间隙内5-HT 的浓度,从而增强5-HT能神经元功能[45]。当抑郁、焦虑发生时,强啡肽和κ阿片受体在中脑边缘区域表达升高,包括中脑多巴胺系统(VTA,PFC)、5-HT能和NE能神经系统、杏仁核、海马和下丘脑。Tao 和Auerbach发现背侧中缝核脑区给予U50,488H可以降低该脑区5-HT的含量[46],Zakharova发现,5-HT 敲除可以阻断 U69593对可卡因引起的自发活动变化的调控[47],提示5-HT系统参与调节U69593引起的行为变化。除影响5-HT释放外,强啡肽/κ阿片受体更能影响5-HT重摄取。Sundaramurthy等发现,在大鼠纹状体及转入5-HT转运体(serotonin transporter,SERT)的 EM4细胞上,κ激动剂预处理能够明显降低SERT在细胞膜上的表达,促进SERT从膜上到胞内的转运,进而降低SERT的重摄取功能[48]。我们认为这是κ激动剂发挥促抑郁样作用的机制。κ阿片受体对SERT功能上的调控具有行为学意义,Schindler等发现,当应激发生时,腹侧纹状体κ阿片受体激活引起SERT的上膜,增加5-HT的重摄取,当SERT敲除时,κ激动剂引起的条件位置厌恶不能形成,提示κ阿片受体可以通过调控5-HT信号通路而影响负性情绪[49]。
Fuentealba等发现系统持续性的κ阿片受体激活可以增加mPFC的DA释放[50],Chudapongse等发现κ激动剂U50,488H可以增加PC12细胞中DA的释放,对于DA重摄取的增加更加明显[51]。Salvinorin A可以增强DA转运体(dopamine transporter,DAT)的活性,降低SERT的活性,对于NE转运体(NE transporter,NET)没有影响[52]。Abraham 等发现强迫游泳应激和κ激动剂U50,488H通过激活DA能神经元上的κ阿片受体发挥行为抑制作用[53]。激动蓝斑κ阿片受体可以增强可卡因引起条件位置偏爱,但是当同时给予NE受体拮抗剂时,该作用即被阻断,说明κ阿片受体通过调节去甲肾上腺素系统而调控成瘾行为[54]。
4 与κ阿片受体调控抑郁行为相关的脑区
4.1 κ阿片受体在杏仁核脑区中的作用
在抑郁患者中,杏仁核发生结构和功能上的改变。研究显示,抑郁患者杏仁核体积明显小于健康人群[55],且杏仁核对于奖赏刺激的反应下降[56]。Stuber等发现杏仁核至伏隔核的兴奋性神经投射介导奖赏效应[57]。Knoll等发现条件化恐惧可以增加杏仁核BLA脑区的κ阿片受体在mRNA水平上的表达,BLA脑区给予拮抗剂norBNI可以降低实验动物的条件化恐惧,并能降低实验动物在高架十字迷宫模型开放臂的时间[58],说明杏仁核(BLA和CeA脑区)强啡肽/κ阿片受体可以调控焦虑行为。Bruchas等发现杏仁核强啡肽/κ阿片受体系统介导CRF引起的焦虑样行为[19]。Nygard等发现BLA脑区κ阿片受体激活介导应激引起的尼古丁复吸[59]。由于成瘾,焦虑和抑郁的发生密切相关,杏仁核脑区在抑郁发生时出现功能和结构上的变化,推测杏仁核脑区强啡肽/κ阿片受体可能调控抑郁行为,值得进一步研究。
4.2 κ阿片受体在海马中的作用
海马是介导应激反应的重要脑区,抑郁患者的海马与健康人群相比明显变小。海马皮质醇类信号传递在HPA轴应激反应中发挥重要作用,长时间的皮质醇信号激活引起海马损伤,包括树突衰退,神经元再生降低、神经可塑性降低[60-61]。 阿片受体在海马中广泛存在[62],在海马脑区,急性强啡肽释放可以降低突触传递和长时程增强[63]。Shirayama 等发现应激反应如强迫游泳、足底电击可以增加海马的强啡肽水平,海马核团给予拮抗剂norBNI可以降低实验动物在习得性无助抑郁模型中的失败次数[64]。在行为测试中,海马CA3脑区给予激动剂引起实验动物在水迷宫和条件化恐惧模型中的记忆损害[65]。重复性的应激刺激引起实验动物产生记忆缺陷,并且可以被κ阿片受体拮抗剂norBNI和强啡肽原敲除所阻断[66]。
4.3 κ阿片受体与中脑DA系统
κ阿片受体在腹侧被盖区及其投射脑区伏隔核中均大量表达。中脑DA系统注射U50,488H可以引起条件位置厌恶[67]。重复给予精神类药物戒断之后伴随CREB和强啡肽水平增加及负性情绪的产生[68-69]。轻微的应激反应可以增加伏隔核DA水平,但是持续性的应激反应却可以降低中脑DA水平[70],系统或者NAc核团给予κ阿片受体激动剂引起伏隔核脑区DA的降低。中脑DA系统是调控奖赏效应的重要环路[71],伏隔核DA的释放和药物奖赏及自然奖赏关系密切,我们推测κ阿片受体引起的伏隔核 DA传递降低可能参与快感缺失。昝等发现慢性吗啡长期戒断可以引起实验动物蔗糖偏好降低和强迫游泳不动时间升高,伴随小鼠伏隔核脑区强啡肽表达增高,伏隔核脑区给予κ拮抗剂norBNI可以阻断吗啡戒断引起的抑郁样行为[72]。
5 κ阿片受体参与抑郁样行为的机制研究
5.1 介导强啡肽抑郁样作用的上游分子
Carlezon 等发现大鼠伏隔核脑区CREB过表达引起实验动物对可卡因奖赏效应的反应降低伴随该脑区强啡肽的表达量升高,伏隔核脑区给予norBNI可以阻断CREB过表达引起的可卡因奖赏效应降低,提示CREB诱导强啡肽的表达,进而影响可卡因的奖赏效应[73]。伏隔核过表达CREB可以增加实验动物在强迫游泳实验中的不动时间,并且可以被伏隔核脑区给予κ阿片受体拮抗剂norBNI阻断[74]。Muschamp等发现伏隔核脑区CREB过表达引起实验动物在ICSS上的阈值升高,相反降低CREB可以升高实验动物在ICSS上的阈值,伏隔核脑区过表达CREB可以降低蔗糖摄入量,转入mCREB可以增加蔗糖摄入量。伏隔核脑区给予κ阿片受体激动剂U50,488H同样可以引起快感缺失,提示伏隔核CREB介导强啡肽表达增加通过激活κ阿片受体发挥快感缺失作用[75]。Chartoff等发现临床抗抑郁药物去甲丙咪嗪可以显著降低应激引起的伏隔核脑区强啡肽原的转录和CREB激活[76],进一步验证了CREB通过调控强啡肽转录介导抑郁样行为。McCarthy等发现,药物刺激引起纹状体CREB丝氨酸133位点磷酸化增强,进而增强CREB对于强啡肽原CRE应答原件的结合能力,最终引起强啡肽原的转录激活增加[77]。这项研究揭示了CREB是如何引起强啡肽原的转录表达增加的,但是目前还不清楚是什么分子引起CREB磷酸化增强,需要进一步研究其具体机制。
5.2 强啡肽/κ阿片受体调控抑郁样行为的分子机制研究
给予激动剂后,κ阿片受体激活Gα,Gα和胞内的信号分子相互作用,进而影响离子通道,持续激活κ阿片受体引起G蛋白耦联受体激酶3(G protein-coupled receptor kinase 3,GRK3)介导的κ阿片受体磷酸化,招募MAPK家族蛋白[78]。MAPK信号通路调节一系列胞内反应,包括细胞增殖、分化、胚胎形成、细胞凋亡、转录因子调控、离子通道磷酸化、蛋白相互作用[79]。有丝分裂原激活蛋白激酶(mitogen-activated protein kinase,MAPK)家族包含12个不同的基因,其中,氨基末端激酶(Jun N-terminal kinase,JNK)、细胞外信号调节激酶(extracellular signal-regulated kinase,ERK)和p38 MAPK的研究比较广泛。MAPK是细胞应激的应答分子,对多种刺激反应,并通过蛋白相互作用把外界刺激转换为胞内信号[80]。
重复强迫游泳刺激引起伏隔核脑区ERK和CREB 磷 酸 化[81],κ阿片受体介导的ERK磷酸化在行为上的意义还不明确。王等研究表明,κ激动剂U50,488H对焦虑行为具有双向调控的作用,高剂量的U50,488H引起抗焦虑反应伴随外侧中膈核(lateral septal nucleus,LSNs)脑区 ERK 激活,而低剂量的U50,488H引起致焦虑行为,伴随伏隔核脑区ERK激活,提示ERK在不同脑区的激活介导U50,488H对焦虑行为的双向调控作用[82]。κ阿片受体介导的p38 MAPK激活的行为学意义得到广泛的研究,p38 MAPK在介导环境压力和炎症反应中发挥重要作用。Bruchas等发现AtT-20细胞系及体外培养的神经元和神经胶质细胞中,U50,488H通过GRK3及arrestin3分子介导p38激活[83],应激可以激活伏隔核脑区GABA能神经元上的p38 MAPK,脑室给予p38抑制剂SB203580可以阻断应激引起的厌恶反应及强迫游泳实验不动时间的升高[14]。昝等发现,U50,488H引起实验动物产生条件位置厌恶,伴随杏仁核脑区p38 MAPK 激活,杏仁核脑区给予p38抑制剂SB203580可以阻断U50488H引起的条件位置厌恶[84]。当SERT敲除时,脂多糖引起的快感缺失即被抑制,表明SERT介导脂多糖引起的快感缺失[85]。在易感快感缺失的小鼠中,前额叶皮层SERT表达量升高[86]。Bruchas等发现特异性敲除5-HT能神经元的p38α亚基,实验动物不能形成社交失败应激引起的抑郁样行为[87],p38α亚基参与介导应激引起的 SERT 转运。Zhu 等发现p38通过PP2A作用于SERT增加5-HT重摄取[88],提示强啡肽/κ阿片受体激活p38,通过PP2A作用于SERT,增加5-HT重摄取而发挥致抑郁作用。
6 总结和展望
κ阿片受体在抑郁模型中的作用已经得到广泛的研究,如在行为绝望、社交失败应激等模型中发挥促进抑郁样效果,激活κ阿片受体引起焦虑行为。κ阿片受体激活可以增加实验动物在ICSS模型中的阈值,引起快感缺失。但是也有文献报道κ阿片受体激动剂可以阻断慢性应激引起的快感缺失,在高架十字迷宫模型上发挥抗焦虑作用。王等发现高剂量U50,488H引起抗焦虑的行为。这些现象出现并不是偶然的,需要进一步探索这些行为反应分歧出现的机制。此外,快感缺失又分为动机型快感缺失和满足型快感缺失,目前基于κ阿片受体在快感缺失中作用的研究大多是关于DA系统紊乱引起的动机型快感缺失,但是对于满足型快感缺失还没有相关文献报道。κ阿片受体介导抑郁的分子机制还有待于进一步研究。激动κ阿片受体所引起的急性反应如快感缺失,延迟反应如强迫游泳不动值,需要进一步研究激动κ阿片受体之后发生急性反应及延迟反应的具体内在机制。CREB在海马和在杏仁核的作用相反,即CREB的作用存在脑区特异性,那么作为CREB的下游信号分子,强啡肽的功能是否存在脑区特异性呢,这个问题值得进一步深究,目前这方面的研究不多。κ阿片受体的研究大多集中在中脑DA系统和杏仁核,其作用比较确切。同样,强啡肽在海马分布很多,它在海马中作用的研究大多基于学习记忆的缺失,在抑郁相关模型中的研究还相对较少,值得进一步探索。
[1] Ronald C Kessler,Evelyn J Bromet. The epidemiology of depression across cultures [J]. Annu Rev Publi Health,2013,34:119-138.
[2] 何海然,薛占霞. 抑郁症相关发病机制的研究进展 [J]. 神经药理学报,2016,6(2):20-25.
[3] 我国精神疾病患者人数超1亿多数是抑郁症患者[Z]. 人民日报(2016-10-14),北京。
[4] George T Taylor,Francesca Manzella. Kappa opioids,Salvinorin A and major depressive disorder [J]. Curr Neuropharmacol,2016,14(2):165-176.
[5] William A Carlezon,Cecile Beguin,Allison T Knoll,et al.Kappa-opioid ligands in the study and treatment of mood disorders [J]. Pharmacol Ther,2009,123(3):334-343.
[6] Allison T Knoll,William A Carlezon Jr. Dynorphin,stress,and depression [J]. Brain Res,2010,1314:56-73.
[7] Bruchas M R,Land B B,Chavkin C. The dynorphin/kappa opioid system as a modulator of stress-induced and proaddictive behaviors [J]. Brain Res,2010,1314:44-55.
[8] 谢小虎,周文华,杨国栋. κ阿片受体研究新进展[J]. 中国药物依赖性杂志,2000,(3):166-169.
[9] McLaughlin J P,Marton-Popovici M,Chavkin C. Kappa opioid receptor antagonism and prodynorphin gene disruption block stress-induced behavioral responses [J]. J Neurosci,2003,23(13):5674-5683.
[10] Carr G V,Bangasser DA,Bethea T,et al. Antidepressantlike effects of kappa-opioid receptor antagonists in Wistar Kyoto rats [J]. Neuropsychopharmacology,2010,35(3):752-763.
[11] Wang Yu-jun,Khampaseuth Rasakham,Huang Peng,et al. Sex difference in κ-opioid receptor (KOPR)-mediated behaviors,brain region KOPR level and KOPR-mediated guanosine 5’-O-(3-[35S]thiotriphosphate) binding in the guinea pig [J]. J Pharmacol Exp Ther,2011,339(2):438-450.
[12] 邹冈,张昌绍. 脑室内或脑组织内微量注射吗啡的镇痛效应[J].生理学报,1962,25:119-128.
[13] 李凌江,马宁. 应激和抑郁[J]. 临床精神医学杂志,2014,1:69-70.
[14] Benjamin B Land,Michael R Bruchas,Julia C Lemos,et al. The dysphoric component of stress is encoded by activation of the dynorphin kappa-opioid system [J]. J Neurosci,2008,28(2):407-414.
[15] Michael R Bruchas,Benjamin B Land,Megumi Aita,et al. Stress-induced p38 mitogen-activated protein kinase activation mediates kappa-opioid-dependent dysphoria [J].J Neurosci,2007,27(43):11614-11623.
[16] Beverly A S Reyes,Guy Drolet,E J Van Bockstaele.Dynorphin and stress-related peptides in rat locus coeruleus:contribution of amygdalar efferents [J]. J Comp Neurol,2008,508(4):663-675.
[17] Michael R Bruchas,Benjamin B Land,Julia C Lemos,et al. CRF1-R activation of the dynorphin/kappa opioid system in the mouse basolateral amygdala mediates anxiety-like behavior [J]. PLoS One,2009,4(12):e8528.
[18] Katia Gysling. Relevance of both type-1 and type-2 corticotropin releasing factor receptors in stress-induced relapse to cocaine seeking behavior [J]. Biochem Pharmacol,2012,83(1):1-5.
[19] Minh P Lam,Christina Gianoulakis. Effects of corticotropinreleasing hormone receptor antagonists on the ethanol-induced increase of dynorphin A1-8 release in the rat central amygdala [J]. Alcohol,2011,45(7):621-630.
[20] 龚雪. γ-氨基丁酸对小鼠焦虑样行为及认知功能的影响[D].上海:复旦大学,2013.
[21] Kang-Park M,Kieffer B L,Roberts A J,et al. Interaction of CRF and kappa opioid systems on GABAergic neurotransmission in the mouse central amygdala [J]. J Pharmacol Exp Ther,2015,355(2):206-211.
[22] William A Carlezon,Cecile Béguin,Jennifer A DiNieri ,et al. Depressive-like effects of the kappa-opioid receptor agonist salvinorin A on behavior and neurochemistry in rats [J]. J Pharmacol Exp Ther,2006,316(1):440-447.
[23] Stephanie R Ebner,Mitchell F Roitman,David N Potter,et al. Depressive-like effects of the kappa opioid receptor agonist salvinorin A are associated with decreased phasic dopamine release in the nucleus accumbens[J]. Psychopharmacology (Berl),2010,210(2):241-252.
[24] Shayla E Russell,Anna B Rachlin,Karen L Smith,et al.Sex differences in sensitivity to the depressive-like effects of the kappa opioid receptor agonist U-50488 in rats [J].Biol Psychiatry,2014,76(3):213-222.
[25] Mitch Harden,Staci E Smith,Jennifer A Niehoff,et al.Antidepressive effects of the kappa-opioid receptor agonist salvinorin A in a rat model of anhedonia [J]. Behav Pharmacol,2012,23(7):710-715.
[26] Wang Qian,Long Yu,Hang Ai,et al. The anxiolytic- and antidepressant-like effects of ATPM-ET,a novel κ agonist and μ partial agonist,in mice [J]. Psychopharmacology(Berl),2016,233(12):2411-2418.
[27] 龙玉. ATPM-ET的抗焦虑、抗抑郁作用评价及机制研究[D]. 大连:大连医科大学,2016.
[28] Jay P McLaughlin,Li Shuang,Joseph Valdez,et al.Social defeat stress-induced behavioral responses are mediated by the endogenous kappa opioid system [J].Neuropsychopharmacology,2006,31(6):1241-1248.
[29] Matthew D Wiley,Laura B Poveromo,John Antapasis,et al. Kappa-opioid system regulates the long-lasting behavioral adaptations induced by early-life exposure to methylphenidate [J]. Neuropsychopharmacology,2009,34(5):1339-1350.
[30] Edgardo Falcon,Kaitlyn Maier,Shivon A Robinson,et al. Effects of buprenorphine on behavioral tests for antidepressant and anxiolytic drugs in mice [J].Psychopharmacology (Berl),2015,232(5):907-915.
[31] Charlotte K Callaghan,Jennifer Rouine,Reginald L Dean,et al. Antidepressant-like effects of 3-carboxamido seconalmefene (3CS-nalmefene),a novel opioid receptor modulator,in a rat IFN-α-induced depression model [J].Brain Behav Immun,2017,67:152-162.
[32] Abdulrahman Almatroudi,Mehrnoosh Ostovar,Christopher P Bailey,et al. Antidepressant-like effects of BU10119,a novel buprenorphine analogue with mixed kappa/mu opioid receptor antagonist properties,in mice[J]. Br J Pharmacol,2017,DOI:10.1111/bph.14060.
[33] Daniela Braida,Valeria Capurro,Alessia Zani,et al.Potential anxiolytic- and antidepressant-like effects of salvinorin A,the main active ingredient of Salvia divinorum,in rodents [J]. Br J Pharmacol,2009,157(5):844-853.
[34] Cindee F Robles,Marissa Z McMackin,Katharine L Campi,et al. Effects of kappa opioid receptors on conditioned place aversion and social interaction in males and females [J].Behav Brain Res,2014,262:84-93.
[35] Elena Chartoff,Allison Sawyer,Anna Rachlin,et al.Blockade of kappa opioid receptors attenuates the development of depressive-like behaviors induced by cocaine withdrawal in rats [J]. Neuropharmacology,2012,62(1):167-176.
[36] Lalanne L,Ayranci G,Filliol D,et al. Kappa opioid receptor antagonism and chronic antidepressant treatment have beneficial activities on social interactions and grooming deficits during heroin abstinence [J]. Addict Biol,2017,22 (4):1010-1021.
[37] Reindl J D,Rowan K,Carey A N,et al. Antidepressant-like effects of the novel kappa opioid antagonist MCL-144B in the forced-swim test [J]. Pharmacology,2008,81(3):229-235.
[38] 李婧,孙建栋,苑玉和,等.谷氨酸能神经传递在抑郁症发病机制中作用的研究进展[J]. 神经药理学报,2014,3(1):20-24.
[39] Danielle M Gerhard,Eric S Wohleb,Ronald S Duman.Emerging treatment mechanisms for depression:focus on glutamate and synaptic plasticity [J]. Drug Discov Today,2016,21(3):454-464.
[40] Ronald S Duman. Pathophysiology of depression and innovative treatments:remodeling glutamatergic synaptic connections [J]. Dialogues Clin Neurosci,2014,16(1):11-27.
[41] Gregory O Hjelmstad,Howard L Fields. Kappa opioid receptor activation in the nucleus accumbens inhibits glutamate and GABA release through different mechanisms[J]. J Neurophysiol,2003,89(5):2389-2395.
[42] Hugo A Tejeda,Ashley N Hanks,Liam Scott,et al.Prefrontal cortical kappa opioid receptors attenuate responses to amygdala inputs [J]. Neuropsychopharmacology,2015,40(13):2856-2864.
[43] Nicole A Crowley,Daniel W Bloodgood,J Andrew Hardaway,et al. Dynorphin controls the gain of an amygdalar anxiety circuit [J]. Cell Rep,2016,14(12):2774-2783.
[44] Guo Ming-yan,Cao De-xiong,Zhu Si-yu et al. Chronic exposure to morphine decreases the expression of EAAT3 via opioid receptors in hippocampal neurons [J]. Brain Res,2015,1628 (Pt A):40-49.
[45] 薛占霞,彭亮. 情感性精神障碍疾病治疗药物的研究现状[J]. 神经药理学报,2011,1(1):55-64.
[46] Tao Rui,Sidney Auerbach. mu-Opioids disinhibit and kappa-opioids inhibit serotonin efflux in the dorsal raphe nucleus [J]. Brain Res,2005,1049(1):70-79.
[47] Elena Zakharova,Stephanie L Collins,Maria Aberg,et al. Depletion of serotonin decreases the effects of the kappa-opioid receptor agonist U-69593 on cocainestimulated activity [J]. Eur J Pharmacol,2008,586(1-3):123-129.
[48] Santhanalakshmi Sundaramurthy,Balasubramaniam Annamalai,Devadoss J Samuvel,et al. Modulation of serotonin transporter function by kappa-opioid receptor ligands [J]. Neuropharmacology,2017,113 (Pt A):281-292.
[49] Abigail G Schindler,Daniel I Messinger,Jeffrey S Smith,et al. Stress produces aversion and potentiates cocaine reward by releasing endogenous dynorphins in the ventral striatum to locally stimulate serotonin reuptake [J]. J Neurosci,2012,32(49):17582-17596.
[50] Fuentealba J A,Gysling K,Andrés M E. Repeated treatment with the κ-opioid agonist U-69593 increases K+-stimulated dopamine release in the rat medial prefrontal cortex [J]. Synapse,2010,64(12):898-904.
[51] Nuannoi Chudapongse,Seong-Youl Kim,Robert Kramer,et al. Nonspecific effects of the selective kappa-opioid receptor agonist U-50,488H on dopamine uptake and release in PC12 cells [J]. J Pharmacol Sci,2003,93(3):372-375.
[52] Bronwyn Kivell,Zeljko Uzelac,Santhanalakshmi Sundaramurthy. Salvinorin A regulates dopamine transporter function via a kappa opioid receptor and ERK1/2-dependent mechanism [J]. Neuropharmacology,2014,86:228-240.
[53] Antony D Abraham,Harrison M Fontaine,Allisa J Song.Kappa opioid receptor activation in dopamine neurons disrupts behavioral inhibition [J]. Neuropsychopharmacology,2017,43:362-372.
[54] Ream Al-Hasani,Jordan G McCall,Audra M Foshage,et al. Locus coeruleus kappa-opioid receptors modulate reinstatement of cocaine place preference through a noradrenergic mechanism [J]. Neuropsychopharmacology,2013,38(12):2484-2497.
[55] 夏军,陈军,周义成,等. 抑郁症患者海马及杏仁核容积异常的 MRI研究[J]. 中华放射学杂志,2005,(2):28-31.
[56] Philip Gorwood. Neurobiological mechanisms of anhedonia[J]. Dialogues Clin Neurosci,2008,10(3):291-299.
[57] Garret D Stuber,Dennis R Sparta,Alice M Stamatakis,et al. Excitatory transmission from the amygdala to nucleus accumbens facilitates reward seeking [J]. Nature,2011,475(7356):377-380.
[58] Allison T Knoll,John W Muschamp,Stephanie E Sillivan,et al. Kappa opioid receptor signaling in the basolateral amygdala regulates conditioned fear and anxiety in rats [J].Biol Psychiatry,2011,70(5):425-433.
[59] Stephanie K Nygard,Nicholas J Hourguettes,Gabe G Sobczak,et al. Stress-induced reinstatement of nicotine preference requires dynorphin/kappa opioid activity in the basolateral amygdala [J]. J Neurosci,2016,36(38):9937-9948.
[60] 王一赫,江虹,李颖,等. 不同时程温和应激对大鼠海马神经元、T淋巴细胞亚群的动态影响 [J]. 中国临床心理学杂志,2013,21(5):731-734.
[61] 成翔,张蕾,姚莉红,等. 慢性应激抑郁状态对大鼠海马神经元再生的影响 [J]. 神经解剖学杂志,2013,29(4):431-434.
[62] Carrie T Drake,Charles Chavkin,Teresa A Milner. Opioid systems in the dentate gyrus [J]. Prog Brain Res,2007,163:245-263.
[63] John J Wagner,Gregory W Terman,Charles Chavkin.Endogenous dynorphins inhibit excitatory neurotransmission and block LTP induction in the hippocampus [J]. Nature,1993,363(6428):451-454.
[64] Yukihiko Shirayama,Hisahito Ishida,Masaaki Iwata,et al. Stress increases dynorphin immunoreactivity in limbic brain regions and dynorphin antagonism produces antidepressant-like effects [J]. J Neurochem,2004,90(5):1258-1268.
[65] Stephanie Daumas,Alexandre Betourne,Helene Halley,et al. Transient activation of the CA3 Kappa opioid system in the dorsal hippocampus modulates complex memory processing in mice [J]. Neurobiol Learn Mem,2007,88(1):94-103.
[66] Carey A N,Lyons A M,Shay C F,et al. Endogenous kappa opioid activation mediates stress-induced deficits in learning and memory [J]. J Neurosci,2009,29(13):4293-4300.
[67] Sante A B,Manoel J Nobre,Marcus L Brandão. Place aversion induced by blockade of mu or activation of kappa opioid receptors in the dorsal periaqueductal gray matter[J]. Behav Pharmacol,2000,11(7-8):583-589.
[68] George F Koob. Neurobiological substrates for the dark side of compulsivity in addiction [J]. Neuropharmacology,2009,56(Suppl 1):18-31.
[69] 俞纲. 中脑腹侧背盖区κ-阿片受体系统对吗啡依赖的调控作用[D].北京:中国人民解放军军事医学科学院,2009.
[70] Elizabeth N Holly,Klaus A Miczek. Ventral tegmental area dopamine revisited:effects of acute and repeated stress [J].Psychopharmacology (Berl),2016,233(2):163-186.
[71] William A Carlezon Jr,Mark J Thomas. Biological substrates of reward and aversion:a nucleus accumbens activity hypothesis [J]. Neuropharmacology,2009,56(Suppl 1):122-132.
[72] Zan Gui-ying,Wang Qian,Wang Yu-jun,et al. Antagonism of κ opioid receptor in the nucleus accumbens prevents the depressive-like behaviors following prolonged morphine abstinence [J]. Behav Brain Res,2015,291:334-341.
[73] William A Carlezon Jr,Johannes Thome,Valerie G Olson,et al. Regulation of cocaine reward by CREB [J].Science,1998,282(5397):2272-2275.
[74] Andrea M Pliakas,Richard R Carlson,Rachael L Neve,et al.Altered responsiveness to cocaine and increased immobility in the forced swim test associated with elevated cAMP response element-binding protein expression in nucleus accumbens [J].J Neurosci,2001,21(18):7397-7403.
[75] John W Muschamp,Ashlee Van’t Veer,Aram Parsegian,et al. Activation of CREB in the nucleus accumbens shell produces anhedonia and resistance to extinction of fear in rats [J]. J Neurosci,2011,31(8):3095-3103.
[76] Elena H Chartoff,Maria Papadopoulou,Matt L MacDonald,et al. Desipramine reduces stress-activated dynorphin expression and CREB phosphorylation in NAc tissue [J].Mol Pharmacol,2009,75(3):704-712.
[77] Michael J McCarthy,Anne-Marie Duchemin,Norton H Neff,et al. CREB involvement in the regulation of striatal prodynorphin by nicotine[J]. Psychopharmacology,2012,221(1):143-153.
[78] Michael R Bruchas,Charles Chavkin. Kinase cascades and ligand-directed signaling at the kappa opioid receptor [J].Psychopharmacology (Berl),2010,210(2):137-147.
[79] Raman M,Chen W,Cobb M H. Differential regulation and properties of MAPKs [J]. Oncogene,2007,26(22):3100-3112.
[80] Karandikar M,Cobb M H. Scaffolding and protein interactions in MAP kinase modules [J]. Cell Calcium,1999,26(5):219-226.
[81] Jay P McLaughlin,Xu Mei,Ken Mackie,et al.Phosphorylation of a carboxyl-terminal serine within the kappa-opioid receptor produces desensitization and internalization [J]. J Biol Chem,2003,278(36):34631-34640.
[82] Wang Yu-jun,Hang Ai,Lu Yu-chen,et al. κ Opioid receptor activation in different brain regions differentially modulates anxiety-related behaviors in mice [J].Neuropharmacology,2016,110(Pt A):92-101.
[83] Michael R Bruchas,Tara A Macey,Janet D Lowe,et al.Kappa opioid receptor activation of p38 MAPK is GRK3-and arrestin-dependent in neurons and astrocytes [J]. J Biol Chem,2006,281(26):18081-18089.
[84] Zan G Y,Wang Q,Wang Y J,et al. p38 mitogen-activated protein kinase activation in amygdala mediates κ opioid receptor agonist U50,488H-induced conditioned place aversion [J]. Neuroscience,2016,320:122-128.
[85] Floor van Heesch,Jolanda Prins,Jan Pieter Konsman,et al. Lipopolysaccharide-induced anhedonia is abolished in male serotonin transporter knockout rats:an intracranial self-stimulation study [J]. Brain Behav Immun,2013,29:98-103.
[86] Yvonne Couch,Daniel C Anthony,Oleg Dolgov,et al.Microglial activation,increased TNF and SERT expression in the prefrontal cortex define stress-altered behaviour in mice susceptible to anhedonia [J]. Brain Behav Immun,2013,29:136-146.
[87] Michael R Bruchas,Abigail G Schindler,Haripriya Shankar,et al. Selective p38α MAPK deletion in serotonergic neurons produces stress resilience in models of depression and addiction [J]. Neuron,2011,71(3):498-511.
[88] Zhu Chong-Bin,Ana M Carneiro,Wolfgang R Dostmann,et al. p38 MAPK activation elevates serotonin transport activity via a trafficking-independent,protein phosphatase 2A-dependent process [J]. J Biol Chem,2005,280(16):15649-15658.
猜你喜欢
磁共振成像(2022年6期)2022-07-30
音乐生活(2020年12期)2020-01-05
智富时代(2019年6期)2019-07-24
智富时代(2019年6期)2019-07-24
中国老年学杂志(2017年13期)2017-07-18
中国药理学与毒理学杂志(2017年4期)2017-06-01
中华老年多器官疾病杂志(2016年7期)2016-04-28
中国社区医师(2015年14期)2015-12-24
医学研究杂志(2015年12期)2015-06-10
文史博览·文史(2014年11期)2015-01-14
