
喷施GA3和6-BA对“富士”苹果顶芽内源激素及成花成枝的影响
2018-06-23 06:59张满让
西北植物学报 2018年5期
马 玲,张 鑫,孟 莹,赵 静,张满让*
(1 西北农林科技大学 园艺学院,陕西杨陵,712100;2 西北农林科技大学 园艺学院科学研究中心,陕西杨陵,712100)
“富士”是中国苹果第一主栽品种,其种植面积占苹果总面积的65%以上[1-3]。但在生产中,“富士”品种容易出现树势过旺,成花困难,从而导致“大小年”现象严重。花芽分化是成花所必须的生理过程,对开花结果起着至关重要的作用,其过程受到多种因素(如激素、温度、枝条类型等)调控[4]。研究发现,激素是调控成花过程的一种有效的方式,在花芽孕育期经GA3处理的“红富士”中CTK和IAA水平都高于对照组,且对花芽孕育有明显的促进作用,但过高浓度的GA3处理会抑制花芽孕育[5]。Bangerh等人[6]研究发现,苹果上采用的多种抑花措施(如喷赤霉素等)均是通过诱导梢尖IAA浓度的增加来抑制花芽分化的。
赤霉素(GAs)是果树抑花最常用的激素之一,其抑制作用在樱桃、苹果和杏树中表现尤为明显,在樱桃上GA3甚至可以调整树体“大小年”现象,提高座果率[7-8]。关于赤霉素的抑花作用已有一系列研究证实,王中原等[9]人发现,在9月中下旬和10月上中旬喷施赤霉素可延迟“金水一号”梨花期4~6 d;陈梅等[10]指出GA3(100 mg/L)对芝麻花序分化具有显著抑制作用。但关于赤霉素对苹果树的抑花效果及其内部机理还需要更加深入的研究。同样,细胞分裂素(CTK)也被证实可促进花芽分化,在花芽分化时期CTK水平往往呈上升趋势,并于花芽形态学分化初期达到顶峰。其中6-BA是第一代细胞分裂素,用于调控细胞生长、分裂、横向萌芽、开花等生理过程[11-12]。钱桦等[13]报道,使用6-BA可促使春石斛花芽分化进程提前并且促进花芽形成及花芽数量增多。里程辉等[14]研究表明6-BA还能降低岳帅苹果座果率,有较好的疏果作用。
可见GA3和6-BA均参与了果树花芽分化的调控,但其在苹果树上的作用效果与机理还需要更深入的探究。众多研究只表明单次喷施激素会影响花芽分化,而多次喷施是否能促进调控效果却涉及很少,且不同激素对不同类型枝条(短枝和中长枝)的作用效果是否存在差异,也鲜有报道。因此,本试验以4年生“富士”树体为材料,在不同生长发育时期、同浓度的GA3、6-BA喷施处理树体和以不同浓度、不同次数GA3、6-BA喷施处理树体,定期取样以测定不同类型枝条顶芽中内源激素(GA3、IAA、ABA、ZT)含量的动态变化,同时统计成花率、成枝率和枝条伸长的程度,来探究GA3和6-BA对于苹果“大小年”现象的可能调控作用以及其对花芽分化的作用。以期为“富士”品种隔年结果现象的调节和苹果花芽分化的进一步研究奠定基础。
1 材料和方法
1.1 植物材料
试验于2016年4~8月在西北农林科技大学千阳苹果试验站进行,材料为4年生、长势一致、砧木为M26的“富士”果树树体。
1.2 试验药品和仪器
GA3、IAA、ABA和ZT标准品均购自Sigma公司,纯度为99%。色谱甲醇为美国天地公司生产。乙酸乙酯、石油醚(沸点30~60 ℃)、丙酮、冰乙酸和纯水均为国产分析纯。仪器:氮吹仪MD200-2、超高效液相色谱LC-30A(日本岛津,分析柱:Shim-pack XR-ODSⅢ(1.6 μm,2.0 mm I.D.*75 mm),检测器:SPD-M30A)。
1.3 处理与采样
1.3.1不同喷施次数在2016年4月,选择生长势、负载量一致的“富士”130棵,随机分成两组;(1)处理组包含:GA350 mg/mL、GA3100 mg/mL、6-BA 50 mg/mL、6-BA 100 mg/mL(依据文献报道[13]、[15])四个小组。从2016年5月1日(花后7 d)开始喷施第一次,标记为1-10号;5月21日喷施第二次,标记为11-20号;6月6日喷施第三次,标记21-30号。每个处理小组每次喷施10棵树,每次喷施时间间隔为20 d,每小组30棵树。(2)对照组,清水,共10棵。
1.3.2不同喷施时间选择生长势、负载量一致的“富士”120棵,随机分为3组,每组含40棵;其中,第一组喷施75 mg/mL GA3,第二组喷施75 mg/mL 6-BA,第三组喷施清水作为对照。每组树体依次标记为1~40号,喷施处理时间为花后1个月(1~10号)、2个月(11~20号)、3个月(21~30号)和4个月(31~40号)。
1.3.3采样在喷施激素2 d后,选取树体0.8~2 m之间的短枝和中长枝分别采取花芽20个,5次重复,同时统计开花率及枝组比例。若处理或采样遇雨天,则提前或推后1 d。采样完成后立即带回实验室液氮冷冻保存。
1.4 内源激素含量测定
1.4.1标准溶液的制备分别称取GA3、IAA、ABA和ZT标准品各0.1 g(精确到0.01 g),配制成10 μg/mL的标准标准贮备液,密封,于-20 ℃储存。分别用甲醇稀释成0、100、200、400、800、1000(ng/mL)的浓度梯度的标准溶液,进行UPLC分析,得出标准曲线。
1.4.2GA3、IAA和ABA的提取与色谱条件参照黄岛平等[16]的方法,略有改动。称0.3 g样品于离心管,加入20 mL体积比为70%的预冷丙酮溶液冷浸12 h,在4 ℃ 8 000 r/min条件下离心10 min,留上清液,以氮吹仪浓缩至少许残留后加入30 mL石油醚萃取,弃醚相留酯相,再加入20 mL乙酸乙酯萃取,留酯相,并将酯相放入氮吹仪浓缩至无残留,加入1.5 mL甲醇溶解,以0.22 μm滤器过滤后定容至进样瓶,4 ℃保存备用。色谱条件:流动相为甲醇:0.8% 冰乙酸=28:72,流速为0.1 mL/min,柱温为25 ℃,进样量为3 μL。
1.4.3ZT提取分离与色谱条件提取方法参照贾鹏禹等[17]的方法略有改变。取0.5 g样品于离心管中,加入5 mL 80%冷甲醇,浸提12 h,12 000 r/min离心30 min,取上清于另一离心管中,并加入3 mL 80%冷甲醇,浸提6 h后,在12 000 r/min离心10 min,上清液以氮吹仪至无残留,然后用1 mL甲醇溶解,0.22 μm滤器过滤,定容至进样瓶,待测。色谱条件:流动相为甲醇、0.6%冰乙酸、水溶液(35∶60∶5),流速为0.1 mL/min,柱温为25 ℃,进样量为3 μL。
1.4.4激素浓度的计算方法待进行UPLC分析后,根据每一种激素的出峰面积结合标准曲线计算出该激素浓度。
1.5 成花率、成枝率及枝条长度的统计
2016年秋季统计每组树体短枝和中长枝数量及长度,计算成枝率。2017年果树盛花期统计每组树体成花率。
1.6 数据分析
使用SPSS 17.0软件进行数据统计分析,用Excel 2007绘图。
2 结果与分析
2.1 激素喷施次数和时期对不同类型枝条顶芽内源GA3含量的影响
如图1,A和1,B所示,随着GA3和6-BA喷施次数的增加,各处理组枝条顶芽中GA3含量的变化趋势与CK组基本保持一致,但是含量显著大于CK组(P<0.05)。单次喷施后,不同处理组短枝顶芽中GA3含量均较CK组增加,且GA3处理增加的幅度更大,50和100 mg/mL GA3处理分别比CK组增加了32.25%和44.38%。与单次喷施相似,多次喷施GA3、6-BA(2次、3次)处理组短枝顶芽中GA3含量也较CK组高,但是GA3含量增幅与喷施次数变化呈相反趋势。同样,单次喷施GA3和6-BA后,中长枝顶芽中GA3含量显著高于CK组,50和100 mg/mL GA3处理分别高于CK组67.54%和111.17%。多次喷施处理组含量也显著高于CK组,但增加幅度较单次喷施低。可见,不论在短枝还是中长枝中,GA3和6-BA喷施次数与GA3含量的增加幅度都呈反比关系。
在短枝中,随着“富士”发育时期的推进,GA3和6-BA处理组顶芽中GA3含量呈先上升后下降的趋势,而CK组GA3含量呈一直降低的趋势(图1,C)。在整个观察时期,GA3处理组含量始终显著高于对照组,而6-BA处理组在花后1个月时低于CK组,后期才增加至显著高于CK组的水平(P<0.05)。在中长枝中(图1,D),6-BA处理组顶芽GA3含量呈在花后2个月内增加之后下降的趋势,且整体含量较对照组低;GA3处理组顶芽中GA3含量除在花后2个月略有降低外,整体增高,且含量始终高于CK组,之间差异显著(P<0.05)。
2.2 激素喷施次数和时期对不同类型枝条顶芽内源IAA含量的影响
如图2,A所示,随着喷施次数的增加,GA3和6-BA处理组短枝顶芽中内源IAA含量的变化趋势与CK组相同,都随着次数的增多含量逐渐增加,但处理组含量要显著小于CK组(P<0.05)。单次喷施后,处理组短枝顶芽中IAA含量均较CK组小;多次喷施后,外源GA3处理组顶芽中IAA含量仍较CK组小,但6-BA处理使得顶芽中IAA含量较CK组高。在中长枝中(图2,B),单次喷施GA3和6-BA后,顶芽中IAA含量与CK组接近,无显著性关系(P<0.05)。多次喷施后处理使顶芽IAA含量增加且显著高于CK组(P<0.05)。综上,无论枝条的类型,顶芽中IAA的含量与激素的喷施次数成正比关系,即随着喷施次数的增加含量的积累增多,但IAA含量增加的幅度随着激素种类和浓度的不同而有所差异。
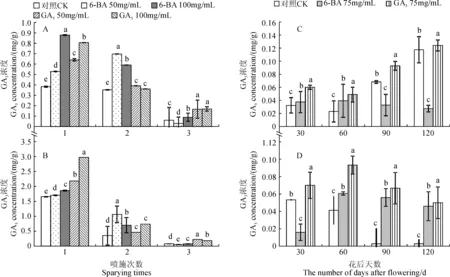
图1 不同GA3和6-BA喷施次数和时期下“富士”短枝(A、C)和中长枝(B、D)顶芽的内源GA3含量Fig.1 The endogenous GA3 concentration in flowering buds of short branches(A,C) and long branches (B,D)of “Fuji” spraying with GA3 and 6-BA at different times and stages
由图2,C可知,在短枝顶芽中,6-BA处理组在花后3个月内基本保持稳定,第4个月时急剧增加,达到峰值。GA3处理组呈先下降后上升再降低的趋势,而CK组随着时间的变化略有降低,在花后4个月增加至峰值。花后1个月GA3处理组IAA含量与CK组基本相同,但6-BA处理组显著低于CK组。之后随着处理天数的增加,在花后2个月时6-BA处理组和GA3处理组IAA含量呈相反的变化趋势,其后两个处理组含量均显著高于CK组,直至观察末期,CK增加至一个显著高于处理组的水平(P<0.05)。 与短枝顶芽激素含量变化不同,除GA3处理在花后4个月略有增加外,处理组和CK组IAA含量随着发育时期的增加呈整体减小的趋势(图2,D),且两个处理组IAA含量在花后1个月和2个月时显著低于CK组,但各处理组之间并无显著相关关系(P<0.05)。之后,随着观察天数的增加,处理组IAA含量积累显著大于CK组,且两处理组之间差异显著(P<0.05)。
2.3 激素喷施次数和时期对不同类型枝条顶芽内源ABA含量的影响
随着喷施次数的增加,处理组短枝顶芽中ABA含量与CK组的变化类似,呈先减小后增加的趋势(图3,A)。单次喷施后处理组短枝顶芽中ABA含量较CK组增加,且6-BA处理增幅较GA3大,50和100 mg/mL 6-BA分别高于CK组35.94%和63.10%;多次喷施后处理组顶芽中ABA含量整体显著低于CK组(P<0.05)。在中长枝中,随着喷施次数的增加,处理组和CK组顶芽中ABA含量呈逐渐上升的趋势(图3,B)。单次喷施后,处理组中顶芽中ABA含量与CK组差异显著,且同激素不同浓度和同浓度不同激素处理之间差异也十分显著;多次喷施后处理组ABA含量较CK组显著增加,且增加幅度与喷施次数呈正比(P<0.05)。可见,不同喷施次数对顶芽中ABA含量均有明显的增加。
随着果树发育时期的推进,6-BA处理组短枝顶芽中ABA含量与CK组变化趋势相同,先增加后减小,而GA3处理组ABA含量逐渐减小(图3,C)。中长枝顶芽中(图3,D), 6-BA处理对顶芽中ABA含量在整个观察时期都无明显的改变作用,而GA3处理组ABA的含量与CK组保持一致,呈逐渐增加的趋势,且在花后4个月时达到峰值。由此可知,外源激素的喷施对于不同类型枝条顶芽中ABA含量的积累整体呈促进作用。

图2 不同GA3和6-BA喷施次数和时期下“富士”短枝(A、C)和中长枝(B、D)顶芽的内源IAA含量Fig.2 The endogenous IAA concentration in flowering buds of short branches(A,C) and long branches (B,D)of “Fuji” spraying with GA3 and 6-BA at different times and stages
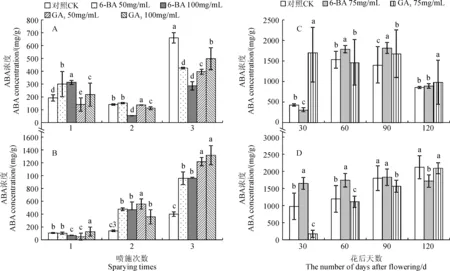
图3 不同GA3和6-BA喷施次数和时期下“富士”(A、C)和中长枝(B、D)顶芽的内源ABA含量Fig.3 The endogenous ABA concentration in flowering buds of short branches(A,C) and long branches (B,D)of “Fuji” spraying with GA3 and 6-BA at different times and stages
2.4 激素喷施次数和时期对不同类型枝条顶芽内源ZT含量的影响
随着喷施次数的增加,处理组短枝顶芽中ZT含量先减小后增加,CK组与其变化趋势相同(图4,A)。单次喷施后,处理组短枝顶芽中ZT含量显著高于CK组,且GA3处理组增幅更大,分别高于对照组30.60%和81.55%;多次喷施后,短枝顶芽ZT含量均不同程度显著高于CK组(P<0.05)。随着喷施次数的增加,GA3和6-BA处理组中长枝顶芽ZT含量变化趋势与短枝相同(图4,B)。单次喷施后,高浓度使ZT含量增加而低浓度使之减小。GA3处理组表现与6-BA处理组相反,高浓度使ZT含量降低,低浓度使之增加。多次喷施后,除2次喷施中50 mg/mL GA3处理组略高于CK组外,其余处理组顶芽中ZT含量整体显著小于CK组(P<0.05)。
处理组短枝顶芽中ZT含量在花后2个月内显著高于CK组,之后则表现相反,显著低于CK组(P<0.05)。在中长枝顶芽中(图4,D),随着树体的发育,GA3处理组顶芽中ZT含量先降低最后增加,CK组变化趋势与之相反。6-BA处理组在花后3个月内基本无变化,观察末期,有所增加,但始终低于CK组。由此可知,在同一发育时期,同处理的ZT在短枝顶芽中的含量要明显低于中长枝。
2.5 激素喷施次数和时期对不同类型枝条成花率、成枝率及枝条长度的影响
2.5.1成花率随着喷施次数的增加,处理组短枝顶芽的成花率均不同程度地低于CK组(图5,A)。单次喷施GA3和6-BA后,短枝顶芽成花率均不同程度的减小,且外源GA3处理的减幅更大;多次喷施后,GA3处理组短枝顶芽的成花率接近30%,相比CK组降低了54%。随着激素喷施次数的增加,中长枝顶芽的成花率逐渐增加(图5,B)。单次喷施后,GA3处理组中长枝顶芽成花率分别比CK组减小了19.71%和28.81%。多次喷施后GA3处理组的减幅依旧大于外源6-BA处理, GA3处理的成花率接近40%,而6-BA处理组成花率高于50%,但显著低于CK组(P<0.05)。可见,在相同喷施次数下同一处理下短枝中成花率要高于中长枝。
随着果树的发育,GA3处理组短枝顶芽的成花率逐渐减小,比率始终低于CK组;6-BA处理呈减小增加的波动变化趋势,且比率始终高于CK组;而CK组在花后2个月内基本无变化,之后略有降低再增加(图5,C)。中长枝顶芽的成花率在外源激素处理下的表现与短枝不同(图5,D),GA3处理组成花率呈先抑制后促进最后抑制的趋势,其成花率始终显著低于CK组(P<0.05);6-BA处理组随树体发育的推进,呈抑制成花率的作用,其成花率在花后3个月内高于CK组,观察末期降低至低于CK组的水平。可见,短枝成花率在同一发育时期高于中长枝,并且外源激素对于不同类型的枝条上成花率的作用不同。

图4 不同6-BA和GA3喷施次数和时期下“富士”短枝(A、C)和中长枝(B、D)顶芽的内源ZT含量Fig.4 The endogenous ZT concentration in flowering buds of short branches(A,C) and long branches (B,D)of “Fuji” spraying with GA3 and 6-BA at different times and stages

图5 不同GA3和6-BA喷施次数和时期下“富士”树体短枝(A、C)和中长枝(B、D)的成花率Fig.5 The flowering rate of short branches(A,C) and long branches(B,D) of “Fuji” tree spraying with GA3 and 6-BA at different times and stages

图6 不同GA3和6-BA喷施次数和时期下“富士”树体短枝(A、C)和中长枝(B、D)的成枝率Fig.6 The branching rate of short branches(A,C) and long branches(B,D) of “Fuji” tree spraying with GA3 and 6-BA at different times and stages
2.5.2成枝率如图所示6,A,随着喷施次数的增加,外施激素对于短枝成枝率的影响整体呈现先促进然后抑制的作用。单次喷施后,处理组中的成枝率整体高于CK组,且100 mg/mL的GA3对成枝率的促进作用最显著短枝数占总体枝数的比例达6.5%以上;2次喷施后,除100 mg/mL GA3显著高于单次处理外,其余均无明显变化,但3次喷施处理均不同程度地抑制了短枝的形成,即成枝率降低,且整体显著小于对照组(P<0.05)。随着喷施次数的增加,外源激素对于中长枝成枝率的作用不同于短枝(图6,B)。单次喷施后,处理组中的成枝率均要不同程度的高于CK组;多次喷施处理组的成枝率逐渐减小并在观察末期整体小于CK组。综上所述,在相同激素、喷施次数处理下中长枝的成枝率要高于短枝,并且外施激素对不同类型枝条的作用效果相同。
如图6,C所示,随着发育时期的推进,GA3处理组的短枝成枝率在花后2个月内基本无变化,之后略有增加再迅速减小,其值在花后1个月内显著低于CK组,之后与CK组无显著差异(P<0.05); 6-BA处理组表现出在花后三个月内略有上升的趋势,而之后迅速降低,其值与CK组无显著差异(P<0.05)。 在中长枝中(图6,D),GA3处理组在花后三个月内无明显变化,之后迅速减小,且其值始终显著低于CK组(P<0.05);6-BA处理组的成枝率呈先减小后增加的趋势,其值在花后三个月内与CK组无显著差异,之后显著高于CK组(P<0.05)。
2.5.3枝条长度在短枝中(图7,A),无论喷施次数多少,处理组枝条长度均要低于CK组,但每个处理之间差异不显著(P<0.05)。在中长枝中(图7,B),单次喷施后,GA3处理组与CK组间无显著差异(P<0.05); 6-BA处理组促进枝条伸长,分别高于CK组72.01%和19.76%;多次喷施后,处理组枝条长度减小并与CK组接近。如图7,C所示,随着果树生育期的变化, GA3处理对于短枝枝条长度无明显作用;而6-BA处理使枝条长度减小并显著小于CK组(P<0.05)。 在中长枝中(图7,D),GA3处理组与CK组无显著差异; 6-BA处理组显著高于CK组(P<0.05)。可见,GA3和6-BA对于不同类型枝条的伸长有抑制的作用。

图7 不同GA3和6-BA喷施次数和时期下“富士”树体短枝(A、C)和中长枝(B、D)的枝条长度Fig.7 The branch length of short branches(A,C) and long branches(B,D) of “Fuji”tree spraying GA3and 6-BA with different times and stages
3 讨 论
多项研究表明植物成花受内源激素调控[18-20],而内源激素水平可以通过外源激素的使用来调节。魏钰等[21]研究发现:外源GA3和6-BA作用于郁金香鳞茎可导致内源GA3和IAA的含量明显升高,说明外施GA3和6-BA可影响内源IAA和GA3等激素水平。在柑桔的生理分化前喷施6-BA显著促进了内源GA3与ZR的积累而明显抑制了ABA的合成[22]。高红兵等人[23]研究发现樱桃苗组培中6-BA的使用使得内源IAA和ZR含量增加,GA3和CTK含量下降。在本研究中,喷施外源GA3和6-BA对苹果顶芽中内源激素(GA3、IAA、ABA、ZT)含量有明显的调控作用。主要表现在:内源GA3整体增加、IAA呈增加趋势、ABA含量整体逐渐增加而ZT总体呈减小趋势。此外,还有研究报道外施6-BA会对植物体内CTK水平产生影响[24],但在“富士”苹果中是否有变化,还有待进一步研究。
植物激素对成花有着至关重要的作用[25-28]。多项研究证实外施GA3对大部分果树成花均有显著的抑制作用[29-30],例如在枣、荔枝、苹果、芒果等树种的花芽分化期间,GA3都发挥了显著的抑制作用[31-32]。有研究表明:GA3可通过促进营养梢的伸长来影响果树花芽分化,从而促进枝条的营养生长,抑制其生殖生长[33]。IAA对于植物成花的作用众说纷纭,有学者认为生长素可以促进成花,但Owens[34]的试验表明生长素抑制剂的使用可促进成花,Lewis也认为生长素不能促花。然而关于生长素在花芽分化中的作用机理,人们更倾向于认为它的存在可能与营养的输入有关。ABA对成花作用的研究目前还不完善,Rakngan认为ABA可促进成花,而Hoad则认为其在成花中起负调节作用。在对“苹果梨”的研究中发现,在生理分化期间,低浓度的ABA有利于花芽分化,而高水平的ABA含量有利于形态学分化,针对不同植物探索有效的浓度阈值并调节ABA至适宜浓度对其调控花芽分化具有重要意义[35]。ZT属于细胞分裂素的一种,在苹果、梨的生理分化期、花蕾分化期、花萼分化期都需要大量的细胞分裂素去促进花原基的形成和发育。研究表明高水平的细胞分裂素有利于花芽分化,而低浓度的细胞分裂素则不利于花芽分化。但也有研究表明,喷施细胞分裂素吡效隆会抑制苹果的花芽分化过程。在花芽分化期中,大多数的学者认为细胞分裂素有促进作用。在本研究中不同发育时期(花后1、2、3、4个月)喷施GA3,对不同类型枝条顶芽的成花均存在抑制作用,且抑制作用和施用时期的早晚呈正相关关系。这是由于喷施外源GA3可促进内源GA3含量的迅速增加,从而强烈的抑制了成花,这与前人的研究成果一致,同时,GA3也会影响內源IAA、ABA、ZT含量的变化,进一步加强对成花的抑制作用。6-BA作为第一代人工合成的细胞分裂素,其对果树成花发育有不同程度的促进效果[36],也被证实可用于促进果树花芽形成[37]。在本研究中,对“富士”使用外源6-BA可对果树的短枝成花产生显著的促进作用(P<0.05),这是因为外源6-BA的使用调节了顶芽中内源激素水平,促进短枝顶芽中GA3、IAA的积累。本研究结果显示连续多次喷施6-AB对于枝条的成花率有着明显的抑制作用,但曾有试验表明了连续喷施5次50 mg/L和2 000 mg/L的6-BA可以增加 “Baldwin”苹果复花率[38],这与本研究的结论相悖,可能是因为随着喷施次数的增加,反转了6-BA对于成花的作用,并且6-BA的浓度对成花有重要的影响,浓度较小时对成花具有抑制作用,当浓度增加到一个界限时,6-BA对于成花的作用会从抑制转变为促进,而这个浓度值目前尚未明确,需要进一步的研究。
果树的萌芽成枝也与内源激素的变化有着密切的关系。曾有报道显示6-BA和GA3均可促进油茶穗条的营养生长,但其效果随着激素浓度的升高而减小[39]。本研究的结果与其相似,在“富士”树体上使用外源GA3和6-BA整体上促进了枝条成枝率。潘根生等人[40]的研究表明经过外源GA3处理后,茶树新梢内源 ABA含量下降,GA3和ZT含量增加,ABA/GA3比值减小。研究认为 GA3有利于ZT的生物合成 ,从而加速新梢生长。在本试验中,喷施外源GA3对ZT的合成也有促进作用,但效果并不显著。内源激素突然急速的变化是成枝率提高的内在诱因,而在本试验中内源GA3的增加和ABA含量的减小也是导致成枝率改变的一个原因,这与牛自勉等[41-42]的研究一致。本试验还发现6-BA的使用对成枝率有显著的促进作用,这与姚春潮等[43]的研究结果一致,经6-BA处理后,枝条伸长受到抑制并不是因为节间数减少,而是因为节间变短造成的。
综上所述,不同喷施次数时,外源激素对于不同类型枝条顶芽中内源激素的调节有着不同的作用,但GA3和6-BA始终抑制枝条成花,与枝条类型无关;GA3和6-BA在单次喷施时都会促进枝条的成枝率,而随着喷施次数的增加成枝率呈降低趋势;GA3和6-BA抑制短枝的伸长,但是随着喷施次数的增加,二者对于中长枝的伸长也表现出抑制作用。对于不同时期喷施外源激素,GA3处理对枝条的成花率、成枝率有明显抑制作用,而对枝条的伸长无明显作用;6-BA在生理分化期前对成花、成枝以及枝条伸长起到了促进作用。所以,若是在生产实践中使用外源激素来进行疏花疏果,可以选择GA3,其抑制成花的效果明显,且随着喷施次数和浓度的增加,抑制作用也更加显著。
参考文献:
[1] 翟 衡,史大川,束怀瑞.我国苹果产业发展现状与趋势[J].果树学报,2007,24(3): 355-360.
ZHAI H, SHI D C,SHU H R. History of the mountains the status and development trend of apple industry in China[J].JounalofFruitScience, 2007,24(3): 355-360.
[2] 尚志华,魏钦平,孙丽珠,等.乔砧“富士”苹果改良高干开心形树冠郁闭的评判参数[J].中国农业科学,2010,43(1):132-139.
SHANG Z H, WEI Q P, SUN L Z,etal. Joe “Fuji” Apple modified high anvil shaped canopy happy evaluation parameters[J].ScientiaAgricultureSinica,2010,43(1): 132-139.
[3] 李慧峰,吕德国,秦嗣军,等.我国苹果产业现状及其可持续发展对策[J].沈阳农业大学学报(社会科学版)2005,7(2):203-204.
LI H F, LV D G, QIN S J,etal. The current situation of apple industry in China and the countermeasures for sustainable development[J].JournalofShenyangAgriculturalUinversity:SocialScienceEdition, 2005,7(2): 203-204.
[4] KOUTINAS N, PEPELYANKOV G, LICHEV V. Flower induction and flower bud development in apple and sweet cherry [J].Biotechnology&BiotechnologicalEquipment,2010,24(1): 1549-1558.
[5] 李天红,黄卫东,孟昭清.苹果花芽孕育机理的探讨[J].植物生理学报,1996,22(3): 251- 257.
LI T H, HUANG W D,MENG Z H. Discussion on the mechanism of flower bud inoculation in apple [J].PlantPhysiologyJournal, 1996,22(3): 251- 257.
[6] BANGERTH P K. Can regulatory mechanism in growth and development be elucidated through the study of endogenous hormone concentration [J].ActaGort,1989,463: 77-78
[7] 刘丙花,姜远茂,彭福田,等.花期喷激素对红灯樱桃坐果率的影响[J]. 落叶果树,2007, 2: 10-11.
LIU B H, JIANG M Y, PENG F T,etal. Effect of blooming hormone on fruit setting rate of red cherry [J].DeciduousFruit,2007,02:10-11.
[8] 杨文莉,周伟权,赵世荣,等. GA3对轮台白杏坐果及果实品质的影响[J]. 新疆农业科学,2017, 4: 597-604.
YANG W L, ZHOU W Q, ZHAO S R,etal. Effects of GA3on fruit setting and fruit quality of Luntai white apricot [J].XinjiangAgriculturalSciences, 2017, 4: 597-604.
[9] 王中原,朱学俭,张永祥.赤霉素、2,4—D对金水一号梨推迟物候期、提高座果率的试验[J]. 落叶果树,1990, 2: 29.
WANG Z Y, ZHU X J, ZHANG Y X. Tests of gibberellins 2,4-D on delaying phenology and increasing fruit setting reat of Jinshui No.1 pear[J].DeciduousFruit, 1990,2: 29.
[10] 陈 梅,李培旺,蒋丽娟.外源激素对蓖麻营养生长及花芽分化的影响[J].中南林业科技大学学报,2011,7: 86-90-114.
CHEN M, LEI P W, JIANG L J. Effects of exogenous hormones on vegetative growth and flower bud differentiation of castor [J].JournalofCentralSouthUniversityofForestry&Technology, 2011,7: 86-90-114.
[11] SUN D, ZHANG H. Voltammetric determination of 6-benzylaminopurine (6-BAP) using an acetylene black-dihexadecyl hydrogen phosphate composite film coated glassy carbon electrode[J].AnalChimActa, 2006,557, 64-9.
[12] OHASHI F, UEDA S, TAGURI T,etal. Antimicrobial activity and their most ability of silver 6-benzylaminopurine montmorillonite[J].ApplClaySci, 2009,46, 296-9.
[13] 钱 桦,刘 燕,郑勇平,等.施用6-BA对春石斛花芽分化及内源激素的影响[J]. 北京林业大学学报,2009, 6: 27-31.
QIAN Y, LIU Y, ZHENG Y P,etal. Application of 6-BA effect on Dendrobium flower bud differentiation and endogenous hormones [J].JournalofBeijingForestryUniversity, 2009, 6: 27-31.
[14] 里程辉,刘 志,王宏,等.不同化学疏花剂对岳帅苹果疏花疏果及果实品质的影响[J]. 江苏农业科学,2014, 11: 180-182.
LI C H, LIU Z, WANG H,etal.The effects of additives on different chemical thinning Yueshuai Apple thinning and fruit quality [J].JiangSuAgriculturalSciences, 2014, 11: 180-182.
[15] 赵 莉,潘远智,朱 峤. GA3、IBA和6-BA对香水百合开花特性及内源激素的影响[J]. 湖北农业科学,2012,51(7): 1 385-1 389.
ZHAO L, PAN Y Z, ZHU A. Effects of GA3,IBA and 6-BA on flowering characteristics and Endogenous hormones of Perfume Lily [J] .HuBeiAgriculturalScience: 2012,51(7): 1 385-1 389.
[16] 黄岛平,徐 慧,蒋艳芳. 超高效液相色谱法测定广西蜜梨花芽中4种植物激素[J]. 大众科技,2011, (10): 184-185+195.
HUANG D P, XU H, JIANG Y F. Determination of 4 plant hormones in flower bud of Guangxi honey pear by ultra high performance liquid chromatography [J].Popularscience&technology, 2011, (10): 184-185+195.
[17] 贾鹏禹,曾明飞,冯乃杰,等. 在线固相萃取-高效液相色谱-串联质谱法同时检测大豆不同部位的4种植物激素[J]. 分析化学,2014,42(12): 1 743-1 749.
JIA P Y, ZENG M F, FENG N J,etal. Online solid-phase extraction, high performance liquid chromatography tandem mass spectrometry simultaneous detection of 4 plant hormones in different parts of soybean[J].AnalysisChemistry, 2014,42(12): 1 743-1 749.
[18] 曹尚银,汤一卒,张俊昌. GA3和PP333对苹果花芽形态建成及其内源激素比例变化的影响[J]. 果树学报,2001, 6: 313-316.
CAO S Y, TANG Y C, ZHANG J C. GA3and PP333on apple flower bud morphogenesis and the changes of endogenous hormone ratio [J].JournalofFruitScience, 2001, 6: 313-316.
[19] 杜 研,李建贵,侍 瑞,等. BA、GA3和IAA对“富士”苹果果形形成的影响 [J]. 植物生理学报,2013, 9: 895-901.
DU Y, LI J G, SHI R,etal.BA, GA3and IAA on the formation of fruit shape of “Fuji” apple [J].PlantPhysiologyJournal, 2013, 9: 895-901.
[20] 吴 俊,钟家煌,徐 凯,等. 外源GA3对藤稔葡萄果实生长发育及内源激素水平的影响[J]. 果树学报,2001, 4: 209-212.
WU J, ZHONG J H, XU K,etal. Effects of exogenous GA3on growth and endogenous hormone levels of Fujiminori grape fruit [J].JournalofFruitScience, 2001, 4: 209-212.
[21] 魏钰,张辉,孟昕,等. 外源GA3和6-BA对郁金香鳞茎内3种激素含量变化的影响[J].植物生理学报,2013,49(2): 161-166.
WEI Y, ZHANG H, MENG X,etal.Effects of exogenous GA3and 6-BA on the contents of three hormones in tulip bulb [J].ActaGort. 2013,49(2): 161-166.
[22] 李学柱,邓 烈. 喷布BA对柑桔内源激素及花芽分化的调控[J]. 园艺学报,1992, (4): 314-318.
LI X Z, DENG L. BA regulation of endogenous hormones and flower bud differentiation in citrus [J].ActaHorticulturaeSinica, 1992, (4): 314-318.
[23] 高红兵,王朋飞,刁绍启,等. 6-BA对酸樱桃组培苗4种内源激素质量分数动态变化的影响[J]. 东北林业大学学报,2007, (7): 46-48.
GAO HB, WANG PF, DIAO SQ,etal.Effect of 6-BA on dynamic changes of four 4 endogenous hormones in tissue culture seedlings of sour cherry [J].JournalofNortheastForestryUniversity. 2007, (7): 46-48.
[24] 王云莉,王成荣,王 然,等.细胞分裂素类生长调节剂对青花菜采后衰老的影响[J].园艺学报,2009,36(11): 1 619-1 626.
WANG Y L, WANG C R, WANG R,etal. Effects of cytokinin growth regulators on postharvest senescence of broccoli[J].ActaHorticulturaeSinica.2009,36(11): 1 619-1 626.
[25] 邱学思,刘国成,吕德国,等杏花芽分化期叶片内源激素含量的变化[J].安徽农业科学,2006,34(9): 1 798-1 800.
QIU X S, LIU G C, LV D G,etal.Changes of endogenous hormones in leaves of apricot during flower bud differentiation [J]. Journal of Anhui Agricultural Sciences, 2006,34(9): 1 798-1 800.
[26] 苏明华,刘志成,庄伊美.椭张龙眼结果母枝内源激素含量变化对花芽分化的影响[J].热带作物学报,1998, (2): 66-77.
SU M H, LIU Z C, ZHUANG Y M. The changes of maternal branch elliptic longan endogenous hormone content influence on flower bud differentiation[J].ChineseJournalofTropicalCrops, 1998, (2): 66-77.
[27] 曹尚银,张俊昌,江爱华,等.苹果花芽孕育调控的最佳时期的研究[J].中国果树,2000,(1): 14-17.
CAO S Y, ZHANG J C, JIANG A H,etal. Study on the optimum period of flower bud regulation in Apple [J].ChineseFruit, 2000, (1): 14-17.
[28] 李秉真,孙庆林,张建华,等.‘苹果梨’花芽分化期叶片激素及核酸含量变化[J].园艺学报,1999,26(3): 188-190.
LI B Z, SUNQ L, ZHANG J H,etal. Changes of leaf hormones and nucleic acid contents during flower bud differentiation of “apple pear”[J].ActaHorticultureSinica, 1999,26(3): 188-190.
[29] TROMP J. Flower-bud formation in pome fruits as affected by fruit thinning[J].PlantGrowthRegul, 1998, 31, 27-34.
[30] SOUTHWICK S M, FRITTS R. Commercial chemical thinning of stone fruit in California by gibberellins to reduce flowering[J].ActaHortic,2001,394, 135-147.
[31] 王学军,郝宝锋.赤霉素对枣树花芽分化和采前落果的影响[J].河北果树,2005,13(1): 13.
WANG X J, HAO B F. Effects of gibberellin on flower bud differentiation and pre-harvest fruit drop of jujube [J].HeBeiFruit,2005,13(1): 13 .
[32] 吴志祥,周兆德,陶忠良,等.妃子笑与鹅蛋荔枝花芽分化期间内源激素的变化[J].热带作物学报,2005,26(4): 42-45.
WU Z X, ZHOU Z D, TAO Z L,etal.Changes of ehormones during flower bud differentiation between Feizixiao and Goose Egg Litchi [J].JournalofTropicalcrops. 2005,26(4): 42-45.
[33] CLINE MG. The role of hormones in apical dominance:New approaches to an old problem in plant development[J].PhysiolPlant,l994,90: 230-237.
[34] OWENS J N.Flowering and seed set.In:A.S.Rahavenmendra(ed.)[M].Physiologyoftrees.Wiley:NewYork,1991:247-271.
[35] 李秉真,孙庆林,张建华,等.苹果梨花芽分化期内源激素含量的变化(简报)[J].植物生理学通讯,2000,(1): 27-29.
LI B Z, SUN Q G, ZANG J H,etal. Changes of endogenous hormones in apple pear during flower bud differentiation (brief) [J].PlantPhysiology,2000,(1): 27-29.
[36] HOAD G V. Hormonal regulation of fruit-bud formation in fruit trees[J].FlowerFruitSetFruitTrees, 1984,149: 13-24.
[37] KRASNIQI A L, DAMEROW L, KUNZ A, BLANKE M M. Quantifying key parameters as elicitors for alternate fruit bearing in cv. ‘Elstar’apple trees[J].PlantSci, 2007,212: 10-14.
[38] MCLAUGHLIN J M, GREENE D W. Fruit and hormones influence on flowering of apple. I. Effect of cultivar[J].JournaloftheAmericanSocietyforHorticulturalScience,1991,116(3): 446-449.
[39] 陈国臣,曾雯珺,金颐熙,等. 6-BA和赤霉素对油茶穗条生长发育及内源激素的影响[J]. 广西林业科学,2014,43(1): 5-9.
CHEN G C, ZENG W J, JIN Y X,etal.Effects of 6-BA and gibberellin on growth and Endogenous hormones of Camellia oleifera [J].GuangXiForestryScience: 2014,43(1): 5-9.
[40] 潘根生,钱利生,吴伯千,等. 茶树新梢生育的内源激素水平及其调控机理(第四报)外源激素对茶树内源激素的影响及其与新梢生长的关系[J]. 茶叶,2001, (2): 25-29.
PAN G S, QIAN L S, WU B Q,etal. The effects of exogenous hormones on the growth of new shoots of tea plants and the relationship between endogenous hormones and shoot growth [J].JournalofTea, 2001,(2): 25-29.
[41] 牛自勉,李 全,部晓梦.苹果短枝形品种树形结构对果实质量的影响[J].山西果树,1994, (2): 4-5.
NIU Z M, LI Q, BU X M. Effects of Apple spur type varieties of tree structure on fruit quality[J].ShanxiFruits, 1994, (2): 4-5.
[42] 牛自勉,房姐仁,许月明.包膜对越冬期苹果枝条脱落酸和赤霉素含量变化的影响[J].华北农学报,1993,8(3): 38-43.
NIU Z M, FANG J R, XU Y M. Effects of abscisic acid and gibberellin change Capsule on the content of winter apple branches[J].ActaAgriculturalBoreali-Sinica, 1993,8(3): 38-43.
[43] 姚春潮,王西芳,张有平. BA不同浓度和喷布次数对新红星一年生苗木侧枝形成的影响[J]. 西北农业学报,1998, (3): 82-84.
YAO CC, WANG XF, ZHANG YP. Effects of different concentration of BA and times of spraying on lateral branch formation of annual seedlings of Xinghongxing [J].ActaAgricultureBoreal-occidentalisSinica,1998, 3: 82-84.
猜你喜欢
金山(2021年10期)2021-11-02
安徽农业科学(2021年11期)2021-07-16
安徽农学通报(2021年10期)2021-06-17
农民致富之友(2020年22期)2020-09-02
热带生物学报(2019年1期)2019-04-18
现代园艺(2017年23期)2018-01-18
现代农业科技(2016年9期)2016-10-20
浙江农业科学(2016年11期)2016-05-04
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
中国果业信息(2013年1期)2013-01-23
