
玉米赤霉烯酮对体外培养鸡脾脏淋巴细胞凋亡的影响
2018-08-15 06:08李欣虹郑若愚辜彦霏陶思邑任志华王亚超邓俊良
动物营养学报 2018年8期
李欣虹 郑若愚 辜彦霏 陶思邑 任志华 王亚超 邓俊良*
(1.四川农业大学动物医学院,成都 611130;2.西南科技大学,绵阳 621010)
玉米赤霉烯酮(ZEA)又称F-2毒素,是由镰刀菌(主要是禾谷镰刀菌,此外还有三线镰刀菌、木贼镰刀菌、雪腐镰刀菌以及粉红镰刀菌等)产生的一种类雌激素样霉菌毒素,在霉变的谷类作物和动物性食品中广泛存在。ZEA可促进细胞凋亡。Kim等[1]研究发现,ZEA可导致小鼠精细胞严重损害,出现不同程度的凋亡小体,且随着作用时间延长和浓度增加而加重,精原细胞和精母细胞表现更明显。余增丽等[2]对乳腺癌的MCF-7细胞研究发现ZEA对细胞的增殖活性是通过调节B淋巴细胞瘤-2(Bcl-2)基因和Bcl-2相关X蛋白(Bax)基因的表达抑制了细胞凋亡所获得的,且ZEA可以显著提高细胞色素P450家族1亚科A多肽1(CYP1A1)酶的活性及其mRNA表达。邓友田等[3]也研究得出ZEA能够促进Bc1-2 mRNA和蛋白表达,而抑制Bax的表达,ZEA可提高人乳腺癌细胞系——MCF27细胞增殖活力并促进有丝分裂指数,并通过对Bc1-2和Bax表达的调节作用,抑制雌激素耗尽所诱导的MCF27细胞凋亡。符达[4]给大鼠饲喂不同剂量的ZEA发现,ZEA可对大鼠p53基因第8外显子的构象产生影响,导致外显子中2个条带上碱基对出现以嘧啶与嘌呤的互换突变为主的突变。而p53基因与细胞周期生长的调节、细胞转化的调节、DNA复制及诱导程序性死亡有密切关系,p53基因通过Bal-2家族作用调控细胞凋亡[5]。在Yu等[6]关于ZEA诱导小鼠RAW264.7巨噬细胞死亡机制的研究中,ZEA染毒处理导致细胞线粒体膜电位的损失及Bcl-2和Bax蛋白在线粒体的变化,细胞质释放细胞色素C和凋亡诱导因子,过氧化氢酶抑制ZEA诱导RAW264.7细胞减少。
ZEA作用于免疫系统的主要靶器官是脾脏,可导致脾脏淋巴细胞凋亡,进而使机体免疫功能降低。马勇江等[7]研究ZEA对离体培养脂多糖活化小鼠脾脏淋巴细胞具有显著促凋亡作用,且促进强度与其剂量呈依赖性关系。但目前关于ZEA对鸡脾脏细胞凋亡方面的研究较少。因此,本研究以原代培养鸡脾脏淋巴细胞为模型,从细胞水平上研究镰刀菌毒素ZEA染毒对鸡脾脏淋巴细胞的凋亡及其调控基因的影响,以便阐明ZEA染毒对鸡脾脏淋巴细胞的影响及其分子机制。
1 材料与方法
1.1 试验材料
试验动物:东北农业大学动物医学院动物中心提供的40~60日龄健康的伊莎公鸡。
胎牛血清(FBS)购自Gibco公司;4-羟乙基哌嗪乙磺酸(Hepes)、ZEA及无酚红的RPMI1640培养基均购自Sigma公司;细胞凋亡检测试剂盒[用异硫氰酸荧光素标记的膜联蛋白V(Annexin V-FITC)和碘化丙啶(PI)双染]购自ACTGene公司;2′,7′-二氢二氯荧光黄双乙酸钠(DCFH-DA)与罗丹明123(Rh123)购自Sigma公司;Trizol试剂盒、M-MLV反转录酶、DNA抽提试剂盒及RNA酶均购自Invitrogen公司;rTaq酶等PCR反应试剂购自TaKaRa(大连)生物公司;溴化乙锭(EB)购自Sigma公司;鸡的Bax、Bak-1、Bcl-2、p53及半胱氨酸天冬氨酸蛋白酶-3(caspase-3)酶联免疫吸附测定(ELISA)检测试剂盒均购自上海生工生物公司。
1.2 试验方法
1.2.1 脾脏淋巴细胞悬液的制备、培养及处理
在无菌条件下,将鸡脾脏取出,放入盛有磷酸盐缓冲液(PBS)的培养皿里,用PBS轻轻洗涤脾脏周围的血液残渣,仔细剥去脾脏周围结缔组织,将其移入另一个盛有PBS并浸泡有200目网筛的培养皿中,用镊子将脾脏放在200目铜网上,用20 mL一次性注射器的内芯轻轻研磨,过滤,将滤液适当稀释成一定浓度的细胞悬液,再将细胞悬液移入预先装有鸡淋巴分离液离心管里,即缓缓以1∶1体积比将细胞悬液移入到鸡淋巴分离液上层,2 000 r/min常温离心15 min,用巴氏吸管移取淋巴细胞,加入冷的PBS洗涤,1 500 r/min 4 ℃离心5 min,弃去上清液,加入不含毒素的RPMI1640完全培养液(加FBS)再洗涤1次,重悬,计数,并用台盼蓝检测细胞活力大于95%。
前期试验已检测染毒48 h时,ZEA的半抑制浓度(IC50)为(23.91±4.96) μg/mL。通过IC50筛选出ZEA的作用浓度为对照组0、Z1组0.10 μg/mL、Z2组0.40 μg/mL、Z3组1.60 μg/mL、Z4组6.25 μg/mL和Z5组25.00 μg/mL[8]。
1.2.2 流式细胞仪测定细胞凋亡率和坏死率
染毒培养48 h时,收集细胞,1 500 r/min离心3 min,用PBS洗涤细胞3次。离心后,加500 μL Binding Buffer制备细胞悬液,再加入10 μL Annexin V-FITC和5 μL PI,室温避光反应5~15 min,用流式细胞仪测定细胞凋亡率和细胞坏死率,同时以不加Annexin V-FITC及PI作为试剂对照。激发波长为488 nm,Annexin V-FITC的绿色荧光通过异硫氰酸荧光素(FITC)通道(FL-1,530 nm)检测,PI的红色荧光通过PI通道(FL-2,585 nm)检测。每个样品以10 000个细胞计数进行统计,数据由流式细胞仪的标准计算机程序分析获得。
1.2.3 细胞内活性氧(ROS)含量的测定
染毒培养48 h,收集细胞,1 500 r/min离心3 min,后用PBS洗涤细胞3次。选用对细胞内过氧化氢(H2O2)特异性结合的荧光染料DCFH-DA对细胞内ROS进行检测,用PBS悬浮细胞,加入DCFH-DA染液使其终浓度为100 μmol/L,混匀,37 ℃避光孵育30 min,PBS洗涤3次,在流式细胞仪测定其平均荧光强度(激发波长488 nm,发射波长530 nm)。
1.2.4 细胞线粒体膜电位的测定
染毒培养48 h,收集细胞,1 500 r/min离心3 min,后用PBS洗涤细胞2~3次。选用对线粒体特异性结合的荧光染料罗丹明123对线粒体膜电位进行检测,用PBS悬浮细胞,加入罗丹明123染液使其终浓度为5 μg/mL,混匀,37 ℃避光孵育30 min,PBS洗涤3次,在流式细胞仪测定其平均荧光强度(激发波长488 nm,发射波长530 nm)。
1.2.5 上清液中Bc1-2、p53、Bax、Bak-1和caspase-3含量测定(ELISA法)
染毒培养48 h后,收集细胞培养上清液,3 000 r/min离心20 min,吸取上清,-20 ℃保存备用,按ELISA试剂盒说明书测定上清液中Bc1-2、p53、Bax、Bak-1和caspase-3的含量。
1.2.6 细胞内凋亡调控基因Bc1-2、p53、Bax、Bak-1及caspase-3的mRNA表达测定
按RNA提取试剂盒说明书操作,最终获得组织总RNA;取组织总RNA适量,按PrimeScript RT-PCR Kit说明书进行反转录,将提取的RNA转化为cDNA,cDNA保存于-80 ℃备用或立即用于PCR。根据GenBank中发表的鸡的β-肌动蛋白(β-actin)、Bc1-2、p53、Bax、Bak-1和caspase-3的全基因序列,应用Prime 5.0软件设计特异的上、下游引物,并经GenBank Blast进行同源性检索后由Invitrogen公司(上海)合成。引物序列及参数见表1。

表1 引物序列及参数Table 1 Primer sequences and parameters
相关基因cDNA的PCR反应体系包括cDNA、上游与下游引物、高保真酶、Premix和ddH2O,反应条件为95 ℃预变性30 s,按95 ℃ 变性5 s,60 ℃退火34 s,40个循环,4 ℃终止反应;取PCR扩增产物于1.5%琼脂糖凝胶电泳2 h,凝胶成像仪拍照记录PCR结果。
1.3 数据分析
采用REST软件(Pfaffl)分析毒素处理样品各目的基因mRNA的表达丰度,应用SPSS 13.0软件对数据进行显著性F检验及相关性分析;其中上述各项指标的测定重复3个不同批次的细胞,每批细胞每个样本重复3次,数据以平均值±标准差表示,P<0.05为差异显著,P<0.01为差异极显著。
2 结 果
2.1 ZEA对鸡脾脏淋巴细胞凋亡率和坏死率的影响
ZEA对鸡脾脏淋巴细胞凋亡率和坏死率的影响见表2。ZEA染毒时,细胞凋亡率各组间差异显著(P<0.05)。细胞坏死率除Z2组和Z3组差异不显著(P>0.05)外,其余各组间差异均极显著(P<0.01)。当ZEA浓度达到6.25 μg/mL以上,细胞凋亡和坏死均有所减缓。
除Z5组较Z4组淋巴细胞的凋亡率和坏死率均有所下降外,其余试验组的细胞凋亡率和坏死率均随ZEA浓度的升高而升高,各ZEA组细胞凋亡率和坏死率均极显著高于对照组(P<0.01);且同浓度下,脾脏淋巴细胞的凋亡率均高于坏死率,表明ZEA染毒主要导致脾脏淋巴细胞凋亡。
2.2 ZEA对鸡脾脏淋巴细胞内ROS含量与线粒体膜电位的影响
ZEA对鸡脾脏淋巴细胞内ROS含量与线粒体膜电位的影响见表3。ZEA染毒48 h,鸡脾脏淋巴细胞ROS含量随毒素浓度的升高而增加,各ZEA组ROS含量极显著高于对照组(P<0.01);而线粒体膜电位随毒素浓度的升高而降低(Z3组除外),各ZEA组线粒体膜电位极显著低于对照组(P<0.01)。ROS含量除Z3组与Z4组间差异不显著(P>0.05)外,其余各组间差异显著(P<0.05);ZEA组间线粒体膜电位差异显著(P<0.05)。

表2 ZEA对鸡脾脏淋巴细胞凋亡率和坏死率的影响Table 2 Effects of ZEA on the apoptotic rate and necrotic rate of chicken splenic lymphocytes %
同列数据肩标不同大写字母表示差异极显著(P<0.01),不同小写字母表示差异显著(P<0.05),相同小写字母表示差异不显著(P>0.05)。下表同。
In the same column, values with different capital letter superscripts mean extremely significant difference (P<0.01), and with different small letter superscripts mean significant difference (P<0.05), while with the same small letter superscripts mean no significant difference (P>0.05). The same as below.
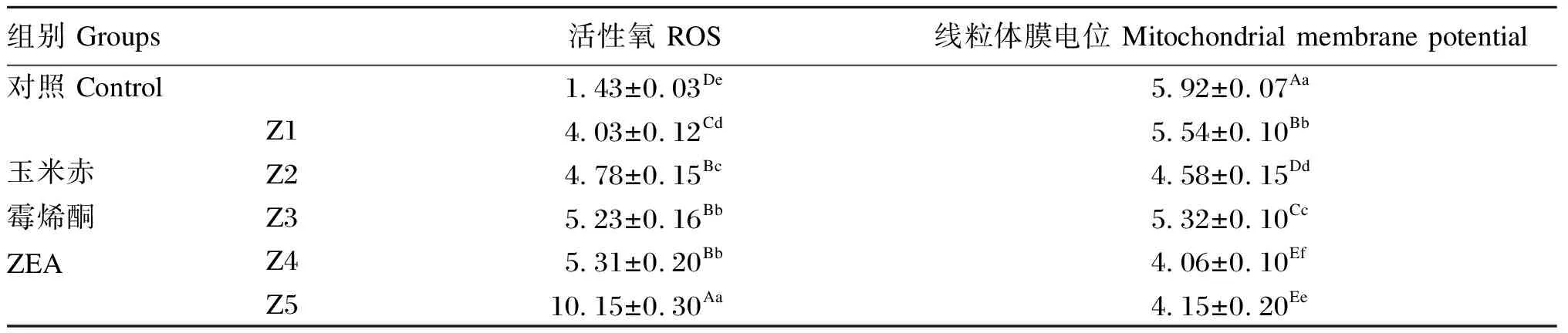
表3 ZEA对鸡脾脏淋巴细胞内ROS含量与线粒体膜电位的影响Table 3 Effects of ZEA on intracellular ROS content and mitochondrial membrane potential in chicken splenic lymphocytes
2.3 ZEA对鸡脾脏淋巴细胞培养上清液中Bc1-2、p53、Bax、Bak-1和caspase-3含量的影响
ZEA对鸡脾脏淋巴细胞培养上清液中Bc1-2、p53、Bax、Bak-1和caspase-3含量的影响见表4。针对Bcl-2含量,除Z1组与Z2组间差异不显著(P>0.05)外,其余各组间差异均极显著(P<0.01);针对p53含量,各组间均差异显著(P<0.05);针对Bax含量,Z1组、Z2组、Z3组和对照组之间差异均不显著(P>0.05),Z4组和Z5组之间及与上述各组间差异极显著(P<0.01);针对Bak-1含量,各组间差异均显著(P<0.05);针对caspase-3含量,除Z3组和Z4组间差异不显著(P>0.05)外,其余各组间均差异显著(P<0.05)。
由此表明,p53、Bax、Bak-1和caspase-3含量随毒素浓度的升高而增加,Bcl-2含量随毒素浓度的升高而降低,具有显著的剂量依赖关系。
2.4 ZEA对鸡脾脏淋巴细胞Bc1-2、p53、Bax、Bak-1和caspase-3 mRNA表达的影响
ZEA对鸡脾脏淋巴细胞Bc1-2、p53、Bax、Bak-1和caspase-3 mRNA表达的影响见表5。针对Bcl-2 mRNA表达量,Z1组、Z2组和Z3组间Bax/Bcl-2值差异不显著(P>0.05),但显著高于对照组(P<0.05),Z4组和Z5组极显著高于其他各组(P<0.01);针对p53 mRNA表达量,各ZEA组极显著高于对照组(P<0.01);针对BaxmRNA表达量,各组间差异显著(P<0.05);针对Bak-1 mRNA表达量,除Z2组和Z3组间差异不显著(P>0.05)外,其余各组间差异极显著(P<0.01);针对caspase-3 mRNA表达量,对照组、Z1组、Z2组和Z3组间差异不显著(P>0.05),Z4组和Z5组极显著高于其他各组(P<0.01),且2组间差异极显著(P<0.01)。
由此表明,除caspase-3 mRNA表达量在Z1组、Z2组和Z3组略小于对照组外,Bax/Bcl-2值以及Bax、Bak-1和caspase-3的mRNA表达量随毒素浓度的升高而增加,具有显著的剂量依赖关系。
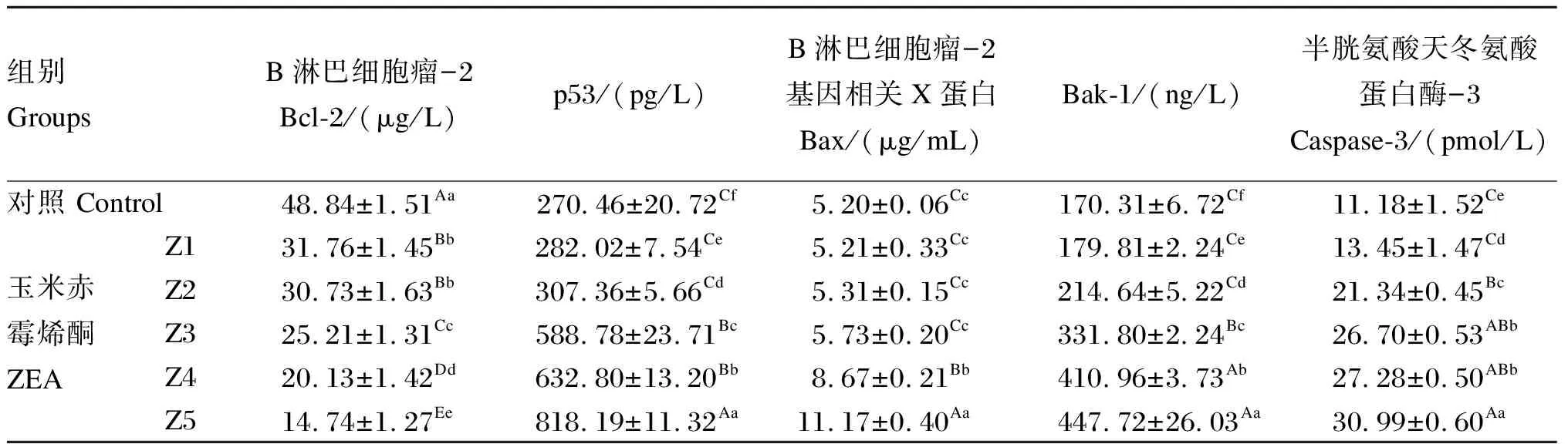
表4 ZEA对鸡脾脏淋巴细胞培养上清液中Bc1-2、p53、Bax、Bak-1和caspase-3含量的影响Table 4 Effects of ZEA on contents of Bc1-2, p53, Bax, Bak-1 and caspase-3 in supernatants of chicken splenic lymphocytes
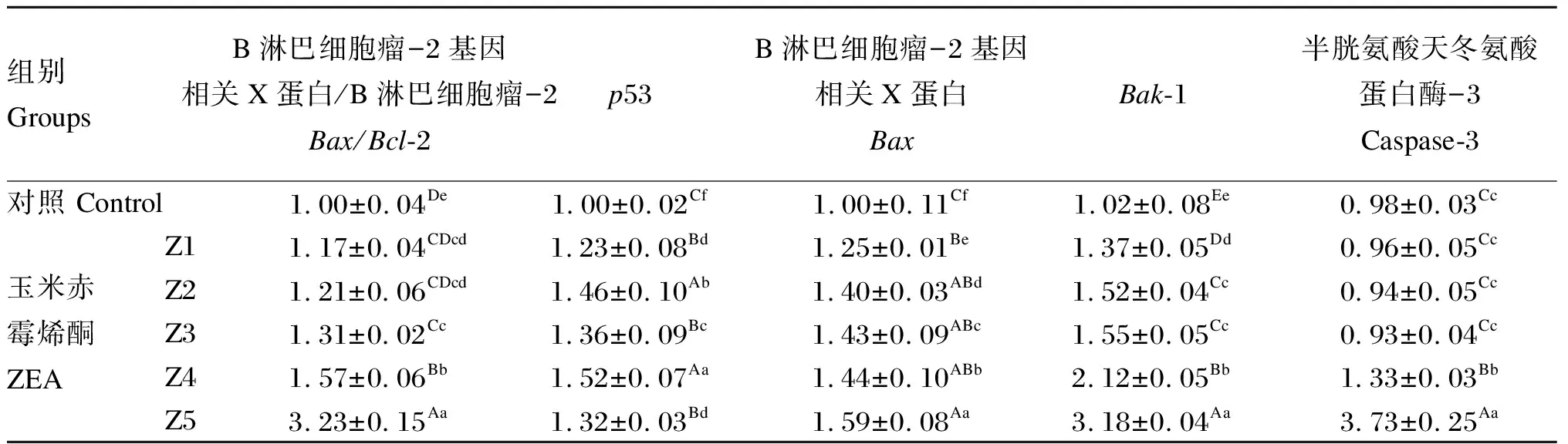
表5 ZEA对鸡脾脏淋巴细胞Bc1-2、p53、Bax、Bak-1和caspase-3 mRNA表达的影响Table 5 Effects of ZEA on mRNA expression of Bc1-2, p53, Bax, Bak-1 and caspase-3 in chicken splenic lymphocytes
3 讨 论
本实验室前期研究表明,ZEA在体内试验中可以诱导小鼠肾脏细胞、脑细胞和肝脏细胞的凋亡[8-11],在体外试验中可以诱导猪脾脏淋巴细胞凋亡[12]。在本试验中,ZEA可以诱导体外培养鸡脾脏淋巴细胞凋亡,结果和前期试验结果一致。
线粒体在氧化代谢过程中产生大量ROS。在自由基产生过多或抗氧化防御系统作用减弱时,机体不能有效清除线粒体内ROS,造成蓄积,过量蓄积的ROS可氧化线粒体渗透性转运孔上的相应氧化还原敏感位点,导致线粒体膜电位降低、线粒体肿胀和进一步产生ROS,进而造成线粒体的氧化损伤[13]。本研究结果表明ZEA染毒使鸡脾脏淋巴细胞内ROS大量积累,线粒体膜电位极显著下降,和上述研究一致。
细胞凋亡的信号传导途径是经过特殊的死亡信号激活半胱氨酸天冬氨酸蛋白酶系统来实现的[14]。caspase-3是在多种诱导剂刺激后导致凋亡的关键酶,它的活化预示着细胞凋亡执行阶段的开始。正常情况下,它以酶原形式存在于胞质中,当细胞接受凋亡刺激时,它被系列反应激活,进而诱导细胞发生凋亡[15-17]。ZEA可诱导猪卵巢颗粒细胞凋亡依赖半胱氨酸天冬氨酸蛋白酶凋亡通路[18]。本研究结果显示,ZEA能够诱导体外培养鸡脾脏淋巴细胞caspase-3的mRNA表达量增加,分泌在上清液中的caspase-3蛋白也增加,且呈剂量效应关系,和上述试验结果一致。
在细胞凋亡过程中,p53蛋白起着至关重要的作用。p53蛋白主要集中于核仁区,能与DNA特异结合,具有明显的细胞转化抑制作用,对保护细胞基因组DNA的完整性具有重要作用。如果DNA受到外来损伤,p53可作为转录因子诱导一系列下游基因的表达,介导细胞停止在G期,使DNA有足够的时间得到修复。如果DNA损伤严重,修复失败、并且不可逆则p53启动凋亡程序诱导细胞凋亡[19]。Ayed Boussema等[20]研究ZEA对人的肝细胞是激活p53,通过p53途径以剂量依赖方式诱导细胞凋亡。Yu等[6]研究ZEA对RAW264.7巨噬细胞的毒性作用,其中p53的激活起到了关键作用。Bouaziz等[21]研究ZEA对人的肝癌细胞的毒性,得出ZEA引发p53依赖的凋亡通路致细胞凋亡的分子机制。本试验结果也表明ZEA从基因水平改变了鸡脾脏淋巴细胞p53的表达,干扰了p53凋亡通路的调控,从而诱发鸡脾脏淋巴细胞凋亡。
Bcl-2家族蛋白在调节通过线粒体使细胞凋亡过程中起着至关重要的作用,Bcl-2蛋白主要分布于线粒体膜、核膜及内质网膜上,参与维持线粒体膜的完整性,防止细胞色素C的释放,阻止凋亡的“内源性激活”,是细胞的抗凋亡成员[22]。当Bax形成同源二聚体Bax-Bax时,诱导细胞凋亡,随着Bcl-2表达量的上升,越来越多的Bax二聚体分开,与Bcl-2形成比Bax-Bax二聚体更稳定的Bax-Bcl-2的异二聚体,从而中和了Bax-Bax二聚体调节细胞凋亡的作用,即细胞内Bax/Bcl-2的值对于决定细胞接受刺激信号后存活与否起关键作用。Bcl-2过量表达细胞存活,Bax过量表达则细胞凋亡[23]。Yuan等[24]研究得出ZEA诱导小鼠雄性生殖细胞的凋亡基因Bax的mRNA的表达增加,Bcl-2的mRNA表达减少。Yu等[25]研究ZEA对人的乳腺癌MCF-7细胞的研究,发现ZEA显著抑制细胞凋亡,存在剂量依赖性,Western blot和多重RT-PCR分析显示,抗凋亡的Bcl-2蛋白和mRNA水平上调,促凋亡的Bax下调,说明ZEA具有雌激素活性,并能促进MCF-7细胞通过细胞周期由G0/G1期减少和S期的显著增加的进展,通过Bcl-2表达调控抑制细胞凋亡。本试验结果显示,在ZEA对鸡脾脏淋巴细胞作用下,Bak-1的表达均有所升高,Bax/Bcl-2值也增加,其蛋白含量也随之增加或减少。
4 结 论
① 本试验表明,随ZEA浓度升高,细胞凋亡调控蛋白p53、Bax、Bak-1和caspase-3的含量升高,Bcl-2含量却下降;ZEA上调Bcl-2家族基因(Bax和Bak-1)、p53基因和caspase-3基因表达,抑制Bcl-2家族基因中Bcl-2基因表达。
② ZEA通过激活p53和半胱氨酸天冬氨酸蛋白酶通路而诱导细胞凋亡增加,并且凋亡是由氧化应激引起。
猜你喜欢
世界科学技术-中医药现代化(2021年7期)2021-11-04
科学导报·学术(2020年19期)2020-07-09
实验动物科学(2019年6期)2019-12-02
教育教学论坛(2019年19期)2019-06-17
环境与生活(2018年7期)2018-09-10
首都公共卫生(2017年3期)2017-11-29
临床医药文献杂志(电子版)(2017年11期)2017-05-17
当代医药论丛(2017年22期)2017-04-12
中国卫生标准管理(2015年15期)2016-01-15
中国药理学通报(2014年2期)2014-05-09
