
莱茵衣藻CrDRBs基因的克隆和生物信息学分析
2018-09-26 09:23娄素琳黄思敏胡章立
深圳大学学报(理工版) 2018年5期
娄素琳,林 鑫,黄思敏,李 辉,4,胡章立,4
1) 深圳大学生命与海洋科学学院,深圳市海洋生物资源与生态环境重点实验室,广东深圳 518060;2) 深圳大学光电工程学院,光电子器件与系统教育部/广东省重点实验室,广东深圳 518060;3) 广东省植物表观遗传学重点实验室,广东深圳 518060;4) 深圳大学龙华生物产业创新研究院,深圳市海洋藻类生物工程技术研究中心,广东深圳 518060
双链小核糖核酸(micro ribonucleic acid, microRNA 或miRNA)结合蛋白(double-stranded RNA-binding protein, DRB)是一类广泛存在于真核细胞、细菌和病毒中,含有双链RNA结合结构域(dsRNA-binding domain,dsRBD)的蛋白.dsRBD 结构域一般由约70个氨基酸残基组成,具有一定种属保守性,dsRBD的结构为αβββα折叠[1-4].DRB在miRNA的生物合成途径中与核酸酶Dicer协同作用,共同精确剪切前体而生成成熟miRNA[5-6].研究表明,DRB在miRNA选择何种作用方式中也起着重要作用[7-8].
在动植物中,一般存在多个DRB蛋白成员,且每个DRB在miRNA途径的分工略有不同[9-11].拟南芥DRB蛋白家族共有5个成员DRB1~DRB5,其中, 研究最透彻的是DRB1(即HYPONASTIC LEAVES1,HYL1).多项研究表明,在拟南芥miRNA 调控途径中,DRB1作为DCL1的重要互作因子,可共同作用、精确有效地从初级转录本剪切生成miRNA,并把其带至AGO1催化的RNA诱导沉默复合物RISC[12-13],执行剪切靶基因mRNA序列或抑制翻译的作用.拟南芥DRB1参与miRNA介导的靶mRNA剪切,且是剪切所必需的,然而DRB2是miRNA介导的翻译抑制所必需的[14].此外,DRB2也是DCL1的互作蛋白,DRB2还抑制DRB1的表达.通过对突变体drb1和drb2蛋白组学的研究,表明DRB1参与的miRNA调控mRNA剪切,广泛存在于各个代谢过程,包括合成代谢和分解代谢,尤其脂肪酸降解途径[14].反之,DRB2相关的翻译抑制显得不太普遍,只是特异性地响应生物或非生物胁迫.
拟南芥DRB1和DRB2均含有两个dsRBD结构域,DRB1的dsRBD结构域和苔藓有50%同源性,而DRB2的dsRBD结构域在两者中有80%同源性,DRB2比DRB1更加保守.文献[6]研究认为,DRB1和DRB2之所以分工不同,主要是蛋白结构域的差异.DRB1的氨基酸序列和DRB2非常不同,分析drb1的转基因株系揭示了DRB1和DRB2的第1个dsRBD结构域在功能上类似,而第2个dsRBD结构域在功能上明显不同,生物信息分析表明DRB2的C-末端结构域在miRNA途径中起功能作用,而DRB1在该位置的结构域是非必须的.
miRNAs调控途径在高等多细胞真核生物已得到广泛深入的研究,但对低等单细胞真核生物的研究非常少[15].莱茵衣藻是一种重要的单细胞模式微藻,同时也是一种极具经济价值的能源微藻.而miRNAs作为一类重要的转录后调控因子,虽然在莱茵衣藻中的研究起步略晚,但其重要地位不容小觑.所在课题组近年对莱茵衣藻miRNAs的研究取得了一系列成果,不仅在莱茵衣藻中发现了一些新的miRNAs存在,且还发现miRNAs在莱茵衣藻缺硫产氢过程中的重要调控作用[16-17].目前莱茵衣藻miRNA作用机制的研究非常少,关于莱茵衣藻CrDRB蛋白在miRNA途径作用的研究更少.2016年底首次报道了莱茵衣藻的DRB蛋白(DUS16),证明了DUS16在pri-miRNA的加工过程起着重要作用,发现DUS16蛋白定位在细胞核和细胞质,且与CrDCL3相互作用[18-19].本研究拟挖掘莱茵衣藻所有DRB蛋白,并对其进行生物信息学分析,为下一步全面揭示莱茵衣藻CrDRB的功能,及其在miRNA调控途径的不同分工奠定基础.
1 实验方法
1.1 莱茵衣藻藻株及培养条件
莱茵衣藻藻株 cc849 购自莱茵衣藻资源中心(https://www.chlamycollection.org/).衣藻在TAP培养基培养至对数生长期,离心收集藻细胞用于总RNA的提取.
1.2 总RNA的提取
① 收集藻体后用液氮研磨成粉末,转至1.5 mL离心管中(粉末不超过管身一半),加入1 mL Trizol抽提液,充分摇匀,漩涡振荡10 min;② 加200 μL氯仿,混匀5 min,室温放置15 min,于10 ℃以下,12 000 r/min离心15 min;③ 吸取上清于另一灭菌的离心管中(上清约600 μL),加入400 μL预冷的异丙醇,-20 ℃沉淀1~2 h,或于-70 ℃沉淀10 min,然后12 000 r/min离心10 min;④ 倒掉上清,保留沉淀,加入1 mL体积分数为75%乙醇(焦碳酸二乙酯,diethyl pyrocarbonate,DEPC),轻弹使沉淀悬起,7 500 r/min离心5 min,倒掉上清,再重复1次;⑤ 室温风干RNA,加入20~30 μL DEPC处理水溶解,-70 ℃下保存备用.
1.3 cDNA的获得
反转录具体步骤参照Promega M-MLV反转录试剂盒说明书,略有改动.① 取5 μL RNA,加5 μL 浓度为10 μmol/L的引物Oligo (dT)20,混匀,70 ℃反应5 min,打开RNA的二级结构,立即置于冰上5 min;② 往上述反应液加入5 μL 5×M-MLV缓冲液、1.25 μL浓度为10 mmol/L的dNTP 混合物、1 μL M-MLV 反转录酶和7.5 μL DEPC处理水,混匀,25 ℃反应5 min,然后42 ℃反应1 h,最后置于70 ℃孵育10 min使酶失活;③ cDNA产物-20 ℃保存.所得的反转录产物可直接作为聚合酶链式反应(polymerase chain reaction,PCR)的模板.
1.4 实时荧光定量PCR
实时荧光定量PCR (quantitative real-time PCR,qRT-PCR)是应用SYBR Premix Ex TagTMⅡ试剂盒(Takara)的反应体系,按说明书操作.PCR反应条件为:95 ℃ 3 min,1个循环;95 ℃ 10 s,58 ℃ 15 s (取决于不同引物的Tm值),72 ℃ 20 s,共45个循环.在65~95 ℃,每0.5 s读取荧光强度分析熔解曲线.每个样品重复3次.实时荧光定量PCR检测的数据采用相对定量的二阶导数法分析(2-△△Ct).
△△Ct=Ctsample-Ctactin
(1)
其中,Ctsample和Ctactin分别是目的基因和内参基因actin的Ct值,Ct为荧光阈值.
1.5 进化树构建
从Phytozome数据库(https://phytozome.jgi.doe.gov/pz/portal.html)下载莱茵衣藻和拟南芥的所有 DRB 基因序列,在美国国立生物技术信息中心(National Center for Biotechnology Information, NCBI)数据库预测DRB的dsRBD结构域,并获取其序列,使用 MUSCLE软件对所有 dsRBD 序列进行对位排列,利用 MEGA7.0 软件构建邻接法(neighbor joining)系统进化树.
2 结果与分析
2.1 莱茵衣藻CrDRB的dsRBD结构域位置
从Phytozome 数据库下载莱茵衣藻的全基因组序列,从蛋白质功能结构域 Pfam 数据库中下载DRB基因的结构域模块PFAM00035,使用 HMMER 软件基于 DRB 结构域检索莱茵衣藻全基因组,获得莱茵衣藻中所有含 dsRBD 结构域的 DRB 蛋白共4个,其中,1个为已经报道的DUS16(Cre05.g232004.t1.1)[18-19].将另外3个DRB蛋白分别命名为CrDRB1(Cre06.g280500.t1.1)、CrDRB2(Cre13.g574650.t1.1)和CrDRB3 (Cre03.g199550.t1.2). 莱茵衣藻的4个DRB蛋白大小为518~2 037个氨基酸残基不等,dsRBD结构域在蛋白的位置如图1.
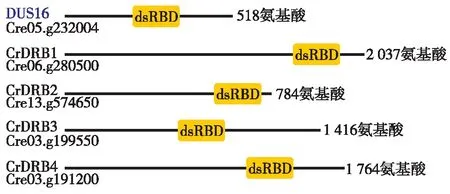
图1 莱茵衣藻CrDRB蛋白的dsRBD结构域位置Fig.1 The location of dsRBD domain of Chlamydomonas CrDRB
2.2 莱茵衣藻CrDRB基因的克隆
根据Phytozome 数据库中莱茵衣藻4个CrDRBs基因的cDNA编码区序列,设计引物,PCR分别扩增4个基因,产物电泳结果如图2,条带大小均符合预期,即DUS16为1 557碱基对(base pair,bp),CrDRB1为6 042 bp,CrDRB2为2 355 bp,CrDRB3为4 251 bp.
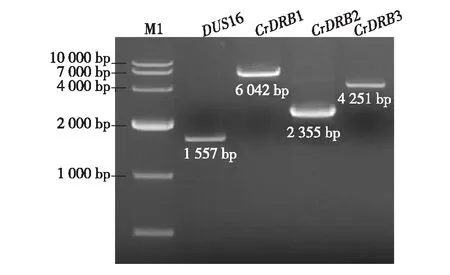
图2 PCR检测莱茵衣藻4个CrDRBs基因Fig.2 PCR detection of four CrDRBs genes in Chlamydomonas
2.3 莱茵衣藻CrDRB基因的表达分析
为分析比较莱茵衣藻中4个CrDRBs基因的mRNA表达水平.收集对数生长期的衣藻细胞,提取mRNA,反转录获得cDNA.每个CrDRBs基因设计3~4对qRT-PCR引物,测试之后选取一对溶解曲线最佳,特异性最好的引物做后续基因表达分析.以actin基因为内参基因,DUS16与actin的表达量比值设为1,其他几个基因的相对表达量分别如图3所示.由图3可见,基因CrDRB3和CrDRB1的相对表达水平显著高于已报道的DUS16[18], 而CrDRB2的相对表达水平则显著低于DUS16.
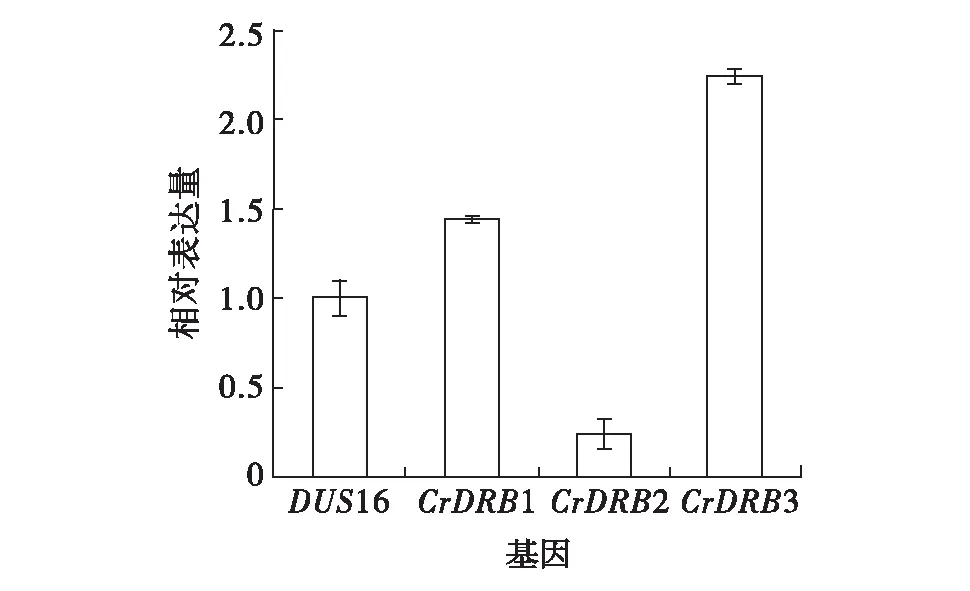
图3 qRT-PCR分析莱茵衣藻4个CrDRBs基因的相对表达水平Fig.3 qRT-PCR analysis of the relative expression of four CrDRBs in Chlamydomonas
2.4 莱茵衣藻CrDRBs蛋白的dsRBD结构域分析
通过NCBI的Protein Blast及结构域的预测分析,获取莱茵衣藻4个CrDRBs蛋白的dsRBD序列,分别将该4段序列通过软件Phyre2在线预测二级结构,方法参考文献[20].莱茵衣藻4个CrDRBs蛋白的dsRBD结构域如图4,CrDRB1、CrDRB3和DUS16蛋白的dsRBD结构域均可形成αβββα的拓扑结构,这是DRB蛋白特有的结构.但是CrDRB2蛋白的dsRBD结构域中β折叠不明显.
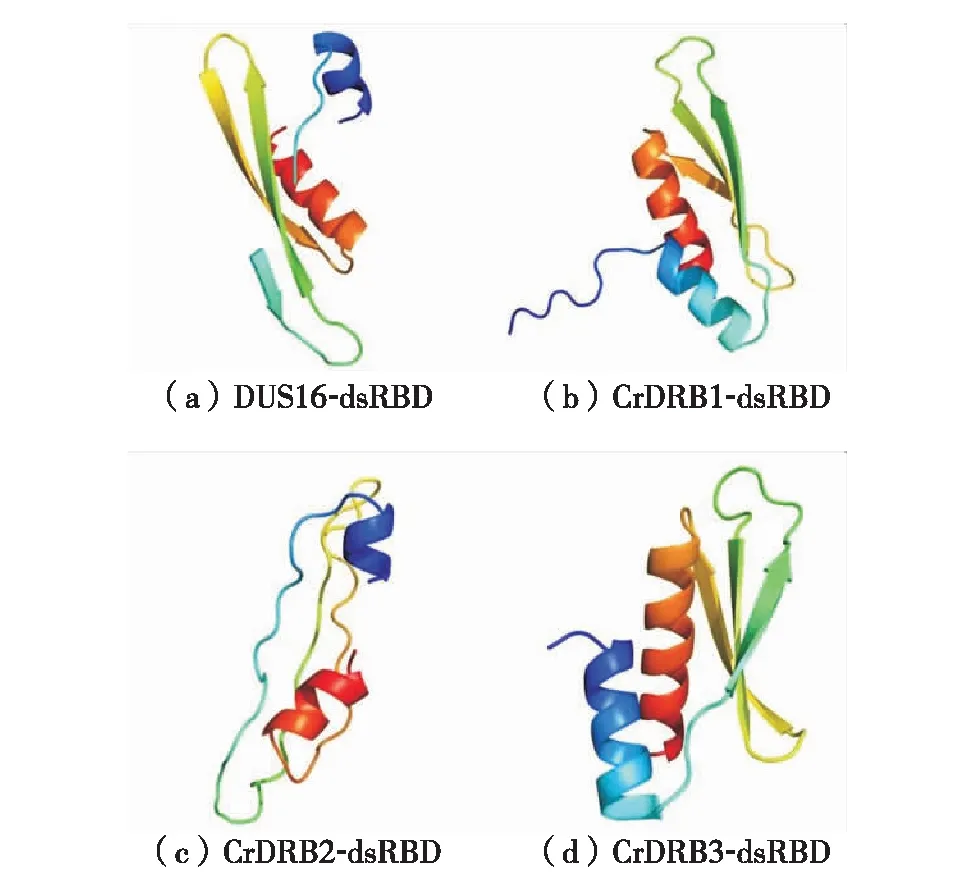
图4 莱茵衣藻4个CrDRBs蛋白的dsRBD结构域Fig.4 dsRBD domain of four CrDRBs in Chlamydomonas
2.5 同源比对分析莱茵衣藻各个CrDRBs基因序列
将莱茵衣藻4个CrDRBs的dsRBD序列,通过BioEdit软件的ClustalW multiple alignment多列比对分析(图5).结果发现,4个dsRBD序列有一定的同源性,其中有3个保守的氨基酸,分别是1个亮氨酸L和2个甘氨酸G,均处于N端.
2.6 莱茵衣藻和拟南芥dsRBD的进化分析
拟南芥中5个DRBs蛋白在miRNA途径的作用有明确分工,为探究莱茵衣藻中4个CrDRBs在miRNA途径可能的功能,及其与拟南芥的异同.将莱茵衣藻CrDRBs的dsRBD结构域与拟南芥5个DRBs成员的10个dsRBD结构域(每个拟南芥DRB均有两个dsRBDs)构建系统进化树(图6).从进化树的结果看出,大致分为了莱茵衣藻CrDRBs与拟南芥DRBs两大簇,其中,莱茵衣藻CrDRB2与DUS16在进化上较相近,而CrDRB3与其他衣藻CrDRBs较远.
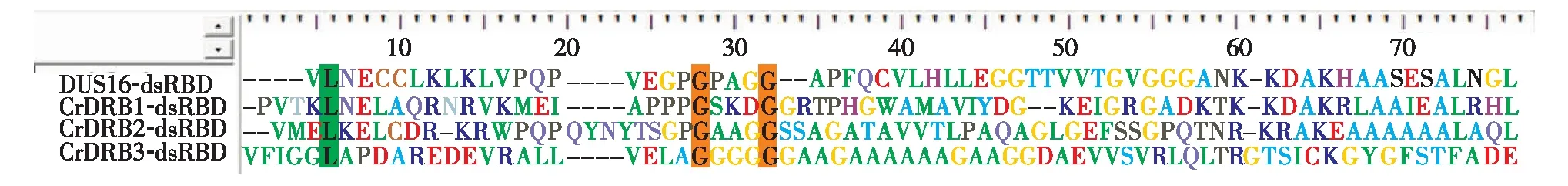
图5 莱茵衣藻CrDRBs蛋白dsRBD结构域同源性比较Fig.5 Sequence alignment of dsRBD domain of CrDRBs in Chlamydomonas
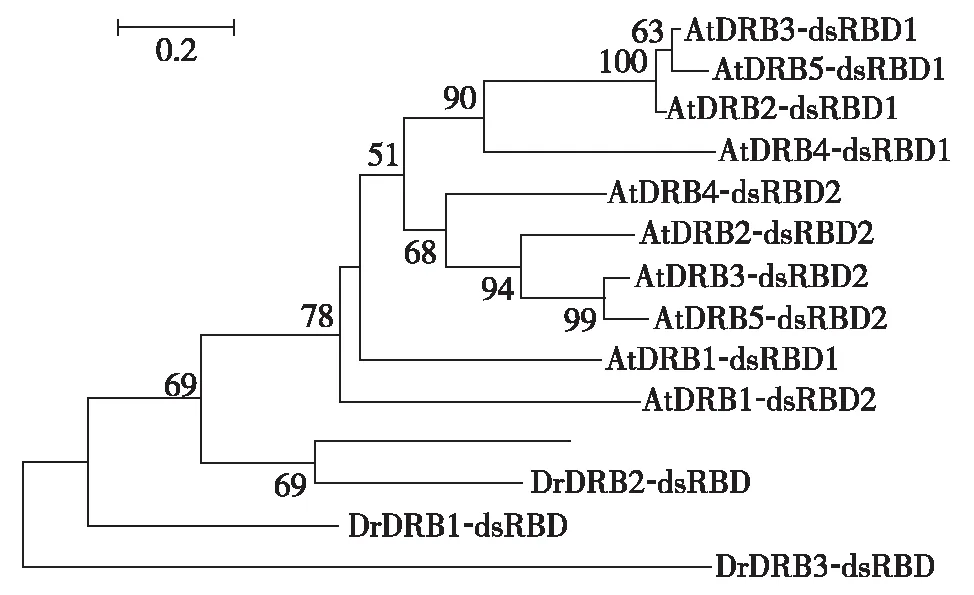
图6 莱茵衣藻与拟南芥dsRBD结构域的进化分析Fig.6 Phylogenetic analysis of dsRBD domain in Chlamydomonas and Arabidopsis
3 讨 论
DRB是一类含有dsRBD的蛋白,除参与RNA的加工和翻译调控外,另一重要功能是,在miRNA的生物合成和作用方式选择方面起着重要作用[21].一般情况,拟南芥或其他高等植物的DRB蛋白大多具有两个或以上的dsRBD结构域,但从图1结果看出,莱茵衣藻的4个CrDRBs蛋白均只有1个dsRBD结构域.造成这种差异的原因可能是莱茵衣藻属于低等生物,在由低等生物到高等植物的进化过程中,dsRBD由1个拷贝演变到多个拷贝.同源比对分析莱茵衣藻4个CrDRBs蛋白的dsRBD结构域,发现相似性不是特别高,有3个氨基酸是保守的.然而,拟南芥DRB成员的dsRBD同源性高达90%.究其原因可能是dsRBD结构域在不同物种之间,尤其是在高等植物和低等微藻之间的差异较大.此外,发现莱茵衣藻CrDRB2的dsRBD结构域不具有典型的完整αβββα拓扑结构,对此下一步将通过RNA结合实验来确定CrDRB2是否为双链RNA结合蛋白.
从文献[9]可知拟南芥的5个DRB蛋白均小于500个氨基酸,最大的DRB2为434个氨基酸.但从图1和2可见,莱茵衣藻的CrDRB蛋白普遍较大,CrDRB1为2 037个氨基酸,CrDRB3为1 416个氨基酸,最小的DUS16为518个氨基酸.莱茵衣藻CrDRBs不仅蛋白较大,且GC含量也较高,导致基因的克隆和蛋白的表达难度也增加了.拟南芥5个DRB蛋白的功能已经很明确,其在miRNA途径的分工也很清晰.但在莱茵衣藻中,目前只有DUS16有相关报道,DUS16与CrDCL3相互作用,共同参与pri-miRNA的剪切.莱茵衣藻其他3个CrDRBs是否也参与了miRNA途径,目前尚不清楚,它们是否参与决定miRNA剪切或抑制翻译的调控方式也尚待进一步研究.
利用qRT-PCR分析莱茵衣藻CrDRBs的基因表达水平,CrDRB3的相对表达量最高,是DUS16的2倍多;CrDRB1的表达水平也较高,约为DUS16的1.5倍;CrDRB2的相对表达量最低.由此说明在莱茵衣藻对数生长时期,CrDRB3 和CrDRB1基因相对较活跃,可能参与了多个生物学过程.
综上可见,DRB是一类双链RNA结合蛋白,在miRNA调控过程起着重要作用.本研究获得莱茵衣藻中4个含 dsRBD 结构域的 CrDRBs 蛋白相关信息,其中包括已经报道的RNA结合蛋白DUS16.分析4个CrDRBs基因的表达情况,发现CrDRB3和CrDRB1的相对表达较高.预测4个CrDRBs的dsRBD二级结构,发现除CrDRB2之外,其他3个CrDRBs蛋白均可形成DRB蛋白特有的αβββα拓扑结构.将莱茵衣藻CrDRBs的dsRBD与拟南芥的5个DRB成员构建系统进化树分析,初步探讨分析莱茵衣藻CrDRBs各成员的的进化保守性.为下一步全面揭示莱茵衣藻CrDRB蛋白功能,及其在miRNA调控途径的具体分工奠定了基础.
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
山西师范大学学报(自然科学版)(2021年3期)2021-11-02
思维与智慧·下半月(2021年4期)2021-04-29
实用肿瘤学杂志(2020年4期)2020-12-08
文萃报·周二版(2020年42期)2020-11-28
进出口经理人(2020年11期)2020-11-24
进出口经理人(2020年10期)2020-11-17
红领巾·探索(2015年9期)2015-09-10
中国老年学杂志(2015年9期)2015-01-31
植物营养与肥料学报(2014年1期)2014-03-11
