
萌发前高温处理对黄茅种子萌发率的影响
2018-11-09 02:45,,,,,,,,,
种子 2018年10期
, , , , , , , , ,
(云南大学生态学与地植物学研究所, 昆明 650500)
干热河谷是对具备干、热2个基本属性河谷带状区域的总称,在此形成了我国西南地区特殊的植被类型——河谷型萨瓦纳,区域内光热资源丰富,气候炎热少雨,水土流失严重,生态十分脆弱[1]。所谓“干热”就是极端水分条件与热量条件的结合。其中,“干”方面,全年的蒸发量大于降水量的3~6倍;“热”方面,年均温>20 ℃和>10 ℃年积温>7 000 ℃。地形深陷封闭而产生的雨影作用以及海拔下降绝热增温和谷地焚风效应是当地气候干热的原因[2]。
干热河谷地区也是全球范围内最易发生火灾的生态系统之一[3-4]。通常认为低强度且周期性的燃烧,对植物的自然更新、丰富度和多样性的维护以及营养循环等方面起着重要作用[5],而火灾强度过大或火烧频率过高则会导致营养物质和总生物量的减少,破坏林分结构和功能,打破生态系统平衡[6-8]。长期进化过程中,许多陆生植物已产生对火的适应性状,其适应性状包括萌生,较厚的树皮,坚硬的种皮,高温或烟诱导种子萌发,果实延迟开放,通过自我修剪降低枯枝等[9-12]。火烧产生的高温将对土壤表面的种子产生一定的影响,有些植物种子会进化形成耐热耐高温能力,过火后保持萌发活力,甚至受到火刺激而获得更高的萌发能力。目前,关于火灾能够促进种子萌发的原因包括火灾能够减少土壤和垃圾中的抑制物质[13-14],火灾烧焦的木头对种子有化学刺激作用[13]以及高温导致种子的硬种皮破碎,刺激种胚,促使种子发芽率提高等[15-17]。种子经高温刺激后提高了萌发率或萌发速度的现象则被称为热冲击效应[18-20]。已有研究发现,非洲萨瓦纳地区——埃塞俄比亚西部实验的21个物种中,有15个物种具有热冲击效应,如:Acacia albida、Acacia Senegal、Acacia seyal、Tamarindus indica等[19]。
黄茅(Heteropogoncontortus)又称地筋,禾本科(Gramineae)黄茅属(Heteropogon),多年生草本,是干热河谷草地土壤种子库和地上植被优势种群[21]。黄茅的须根发达,喜热且抗旱,耐贫瘠土壤,是高温生长响应植物[22]。目前,关于黄茅的研究主要集中在形态特征[23]、干旱环境的适应[24-25]、对其他物种生长的影响[26]等,对于种子是否具有高温适应性一直没有研究。本研究通过高温预处理黄茅种子,了解热冲击对黄茅种子萌发的影响,探讨黄茅种子是否对高温具有适应能力,进而讨论火在干热河谷植被群落构建中所扮演的作用,从而为干热河谷植被保护与恢复提供理论依据。
1 材料与方法
1.1 试验材料
供试材料2016年11月采于四川攀枝花市,对比实验选用的种子于同期采于云南昆明呈贡区。选取颗粒饱满、色泽一致、大小均匀的种子作为试验种子。
1.2 试验方法
1.2.1 材料处理
本试验萌发前的热处理主要参照Ribeiro等[20]的实验进行设计。
选用60 ℃(10,20,40 min)、80 ℃(5,10,20 min)和100 ℃(2.5,5,10 min)3个温度梯度,每组3个加热时间,加上1个常温(25 ℃左右)对照,共计10个处理。每处理5个重复,每个重复20粒种子。
取培养皿3个,每个培养皿中放置100粒种子,单层放置不重叠;设定好烘箱温度,待烘箱温度达到设定的温度后,迅速将装有黄茅种子的培养皿置于烘箱中进行高温处理;达到处理时间后迅速取出,转入备好的常温玻璃皿中;待种子自然冷却至室温后进行萌发实验。
攀枝花、元谋的黄茅种子均按上述方法进行热处理,再进行萌发实验,以进行对比研究。
1.2.2 萌发实验
将种子置于直径为(10±1)cm的铺有湿润滤纸的培养瓶中,放入培养箱进行种子萌发实验。
培养环境设置为:温度25 ℃,光照强度1 250 lx,光照时间14 h/d,适时补充水分,保持培养瓶底部湿润且种子周围不出现水膜。前7 d每2 d记录1次种子的萌发数量,7 d以后3 d记录1次(种子萌发以胚根突破种皮2 mm为标准),记录至连续7 d无种子萌发为止。
1.3 数据处理
参照董沁方等[27]的方法,计算种子萌发率和热冲击效应指数,如下:
萌发率(%)=(n/N)×100%
,
式中:n表示正常萌发数量;N表示供试种子数量。
萌发势(%)=(规定时间(7 d)内发芽的种子数/供试种子总数)×100%。
热冲击效应指数(RI)计算式为:
RI=1-C/T(当T≥C时)
,
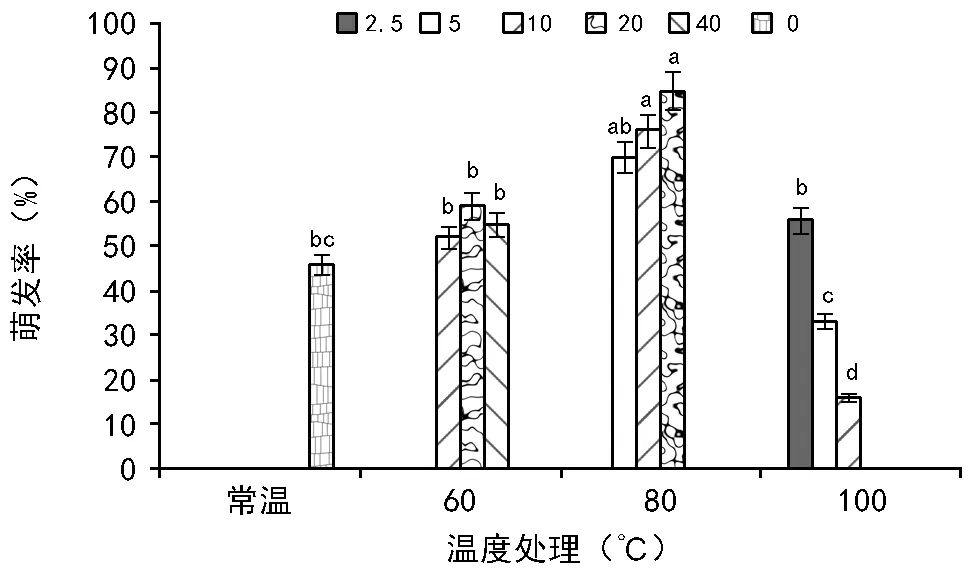
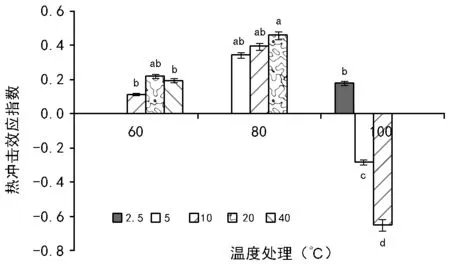
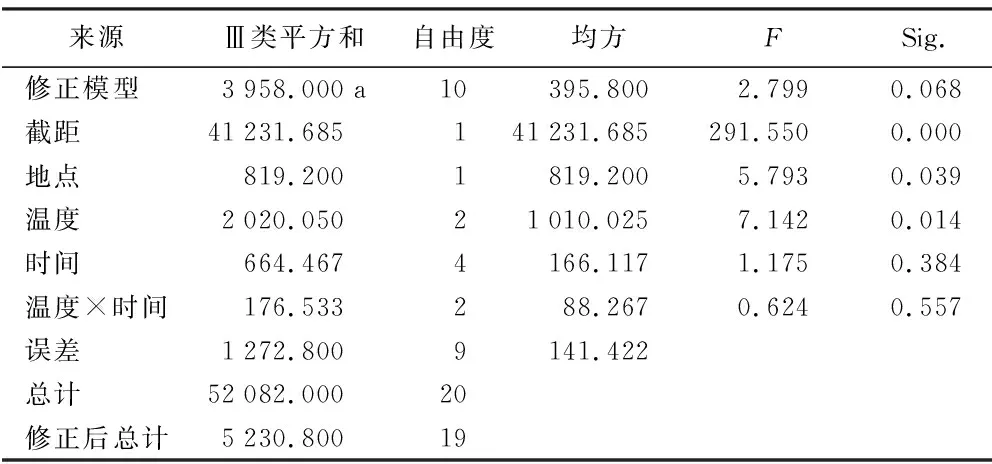
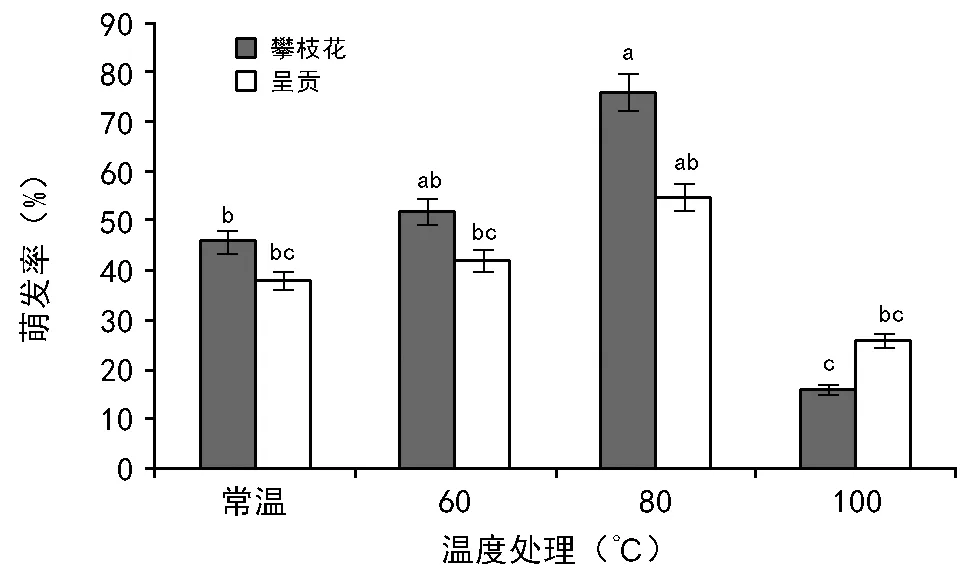
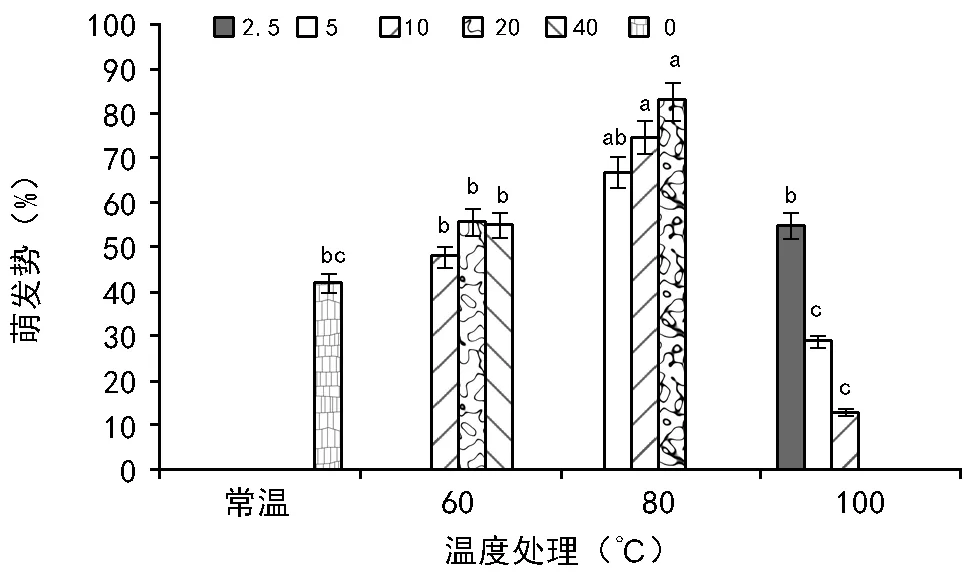
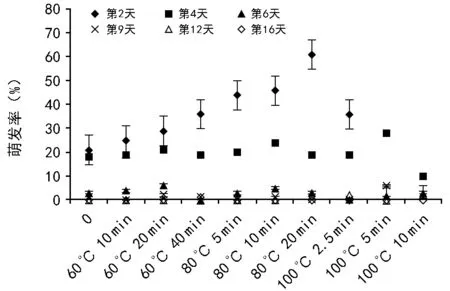
RI=C/T-1(当T , 式中:C为对照值,T为处理值。当RI>0 时,为促进作用;当RI<0 时,为抑制作用。RI的绝对值代表作用强度的大小。 利用Excel 2007软件对数据进行统计分析并作图,利用SPSS 11.0 软件通过95%水平上进行单因素方差(ANOVA)分析,采用Duncan 检验法进行多重比较及差异显著性检验(α=0.05),图表数据为平均值±标准差。 2.1.1 萌发率的变化 萌发前经60 ℃ 10,20,40 min 处理后,其萌发率均高于未处理种子组,分别提高了6%、13%、9%,但差异并不显著(图1)。经高温处理后,萌发率最高可达(77.00±7.55)%,最低仍有(35±20.07)%。 随着温度的升高和处理时间的变化,整体上呈现出先增高后降低的趋势,80 ℃ 10 min和20 min的处理组显著高于其他各组及对照,在80 ℃ 20 min处理时达到最大值(85±4.12)%。100 ℃ 10 min处理显著低于其他各组及对照。 图1 萌发前高温处理对黄茅种子萌发率的影响 2.1.2 热冲击效应指数 热冲击对黄茅种子萌发率的影响可以用热冲击效应指数来表示(图2)。 图2 不同温度处理种子萌发的热冲击效应指数 热冲击效应指数大于0,表明该处理组的萌发率大于未处理组,即促进了种子的发芽。反之,则抑制了种子的萌发。结果表明,在60 ℃、80 ℃的3个时间梯度以及100 ℃ 2.5 min的热冲击效应指数均大于0,表明以上处理促进了黄茅种子的发芽。但在100 ℃ 5,10 min这2个温度-时间处理下,其热冲击效应指数小于0,即抑制了黄茅种子的发芽。从整体上看,随着温度的升高和时间的延长,高温预处理对黄茅种子表现为先促进后抑制的作用。 2.1.3 主效应方差分析 使用SPSS 软件中一般线性模型,对萌发前不同地点经不同温度和时间处理的云南松种子的萌发率进行单变量双因素方差分析(表1)。 结果显示,地点因素和温度因素的主效应显著,但是时间因素以及温度时间两者的互相作用效应并不显著。根据各因素F值的大小可知,萌发前的处理温度对萌发率的影响最大(F=7.142,p=0.014),其次是不同的地点对种子萌发率的影响(F=5.793,p=0.039)。 表1 萌发率主效应方差分析结果 来源Ⅲ类平方和自由度均方FSig.修正模型3958.000a10395.8002.7990.068截距41231.685141231.685291.5500.000地点819.2001819.2005.7930.039温度2020.05021010.0257.1420.014时间664.4674166.1171.1750.384温度×时间176.533288.2670.6240.557误差1272.8009141.422总计52082.00020修正后总计5230.80019 注:R2=0.757(调整后R2=0.486)p<0.01表示差异极显著,p<0.05表示差异显著。 基于主效应方差分析结果,对不同地点样品进行比较。所有处理均有10 min处理方式,因此选取不同地点不同温度处理10 min的数据进行分析(图3)。从图中可以看出:在常温条件下,2个地点的黄茅种子萌发率也很高。在60 ℃和80 ℃高温下处理10 min之后,萌发率显著高于未处理组,均具有热冲击效应。采于攀枝花的种子,在相同处理下,除100 ℃之外萌发率要高于采于呈贡的黄茅种子。在80 ℃处理下,两地的黄茅种子萌发率均达最高。 图3 萌发前高温处理对不同地点黄茅种子萌发率的影响 黄茅在常温条件下,萌发势为(42±1.97)%(图4)。经过不同时间和温度处理之后,黄茅的萌发势呈现出先增加后降低的趋势,80 ℃ 3个时间处理组萌发势均显著高于其余各组,与种子萌发率规律一致,且在80 ℃ 20 min达到萌发势最高值(83±5.22)%。 在60 ℃的处理下,萌发势虽比常温条件下的萌发势高,但并不显著。在经过80 ℃处理下的黄茅种子的萌发势要显著高于常温对照组,100 ℃ 5,10 min处理下的萌发势要显著低于常温对照组,在100 ℃ 10 min处理下达最低值(13±1.19)%。 图4 萌发前高温处理对黄茅种子萌发势的影响 黄茅在高温处理后进行萌发实验的第2天,整体萌发率最高(除100 ℃ 5,10 min未萌发,图5)。并且随着处理时间的变化以及温度的升高,萌发率呈现出先上升后下降的趋势,以80 ℃ 20 min的萌发率为最高。随着时间的推移,黄茅种子的萌发率呈现出时间越长萌发率越低的趋势。前4 d黄茅种子的萌发率相对而言最高。至第6天之后,仍有少量种子萌发,但萌发率不高。 图5 不同温度条件下黄茅种子的萌发进程 Tarrega R等[28]在对Cytisusscoparius和Genistaflorida进行适度的高温处理后,出现了萌发率显著提高的现象。Ribeiro等研究发现,巴西稀树草原的常见物榆叶梧桐(Guazumaulmifolia)经100 ℃,10 min处理后打破了种子休眠,促进了种子的萌发[20]。宋红等研究表明,笃斯越桔(Vacciniumuliginosum)和柳叶绣线菊(Spiraeasalicifolia)种子在适宜的高温处理(120 ℃)下,能够显著提高其种子的萌发率,并且对毛赤杨(Alnussibirica)和珍珠梅(Sorbariasorbifolia)这2个物种的种子萌发率也有一定的促进作用[29]。常云霞等[30]在对云南松(Pinusyunnanensis)种子进行高温处理后,发现云南松的种子萌发前经1,5 min,60 ℃处理后可提高萌发率,存在热冲击效应。本次实验中,黄茅种子经60 ℃ 10,20,40 min以及80 ℃ 5,10,20 min和100 ℃ 2.5 min处理后其萌发率显著提高,具有明显的热冲击效应,与前人研究具有相似的现象,表明黄茅种子对高温具有一定的适应能力。 热冲击不仅能够提高黄茅种子的萌发率,还可以提高其萌发势。在萌发前经60 ℃ 10,20,40 min和80 ℃ 5,10,20 min以及100 ℃ 2.5 min处理后,其萌发势高于对照组,明显缩短了种子萌发的时间(图4)。黄茅随着时间的推移表现出种子萌发率的下降(图5),表明黄茅种子是属于早萌发且萌发快的种子,这可能是黄茅在干热河谷环境中的一种生长适应对策,也可能是由于短暂的高温处理能够有助于打破种子休眠有关,与王海宁等[21]关于黄茅的研究一致。 攀枝花市位于金沙江及其雅砻江的交汇处(101°08′~102°15′E, 26°05′~26°27′N),属典型南亚热带干热河谷气候。据攀枝花市气象资料统计,市区年平均气温20.3 ℃,极端最高温达41.2 ℃,极端最低温为13 ℃,全年日照时数长达2 300~2 700 h,年总降雨量为820~1 100 mm。具有干雨季分明,日照充足,太阳辐射强,蒸发量大,气候垂直差异显著等特征。呈贡区位于云南省滇东高原腹地(102°45′~103 °00′E,24°42′~25 °00′N),属低纬度高原季风型,光照充足。据昆明市气象资料统计,市区多年年均气温14.7 ℃,极端最高温31.2 ℃,极端最低温-7.8 ℃,年平均日照时数2 200 h,全年无霜期285 d,全年平均降雨789.6 mm,气候温和。根据中国气象局的资料显示,在攀枝花及昆明地区均有火灾发生记录(例如,2015年3月11日,云南的怒江、大理、昆明呈贡、石林等地已发生多起森林大火,森林防火形势严峻;2016年4月13日,四川省攀枝花市发生森林火灾)。 中国的干热河谷因其独特的地理环境和气候条件,被称为半自然稀疏草原,即半萨瓦纳植被类型(semi-natural savanna)[2]。目前,关于植物种子对高温适应性状的研究主要集中在非洲以及欧洲的萨瓦纳地区,而中国的干热河谷研究较少。已有研究表明,中国干热河谷的灌木优势种坡柳(Dodonaeaviscosa)在60 ℃ 10,20,40 min,80 ℃ 5,10,20 min以及100 ℃ 2.5,5,10 min的不同温度和时间处理下,其萌发率显著高于常温对照组,具有明显的热冲击效应[31]。说明干热河谷中的植被在长期的高温生长过程之中或者是火灾的威胁之下,已经形成了对高温生境与火生境的积极适应性状,能够在高温刺激下促进种子的发芽。攀枝花、呈贡的黄茅种子萌发前经过10 min的处理之后,在60 ℃和80 ℃的高温下其萌发率高于常温对照组(图3),均在80 ℃时萌发率最高。取自干热河谷攀枝花的居群其萌发率要高于低纬度高原季风型的呈贡地区。但是呈贡地区的黄茅种子也同时表现出了明显的热冲击效应。表明在长期的进化过程之中,热冲击效应已经成为了一种可遗传性状。但从分布区特点来看,干热河谷区气温更高、更易发生火灾,其黄茅居群也表现出更为明显的热冲击效应。此外,坡柳与黄茅种子分别在80 ℃ 10 min以及80 ℃ 20 min萌发率最高,可能是由于在相同的生境中的物种在长期的进化过程之中形成了“趋同进化”的现象,同时也为干热河谷的植被恢复提供了理论指导。
2 结果与分析
2.1 萌发前高温处理对黄茅种子萌发率的影响
2.2 萌发前高温处理对不同地点的黄茅种子萌发率的影响
2.3 萌发前高温处理对黄茅种子萌发势的影响
2.4 不同温度条件下黄茅种子的萌发进程
3 结论与讨论
猜你喜欢
小哥白尼(神奇星球)(2021年9期)2021-11-19
小哥白尼(神奇星球)(2021年5期)2021-07-28
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
哈尔滨轴承(2021年1期)2021-07-21
保鲜与加工(2021年1期)2021-02-06
矿产综合利用(2020年1期)2020-07-24
爆笑show(2015年3期)2015-05-08
汽车维修与保养(2015年12期)2015-04-18
汽车维修与保养(2015年6期)2015-04-17
