
多花黄精种子后熟过程生理研究
2018-11-09 02:45,,,,,,,,
种子 2018年10期
, , , , , , , ,
(1.西南中药材种质创新与利用国家地方联合工程研究中心, 云南 昆明 650201;2.云南农业大学资源与环境学院, 昆明 650201; 3.金平慧达农林开发有限公司, 云南 红河 661500)
多花黄精(PolygonatumcyrtonemaHua.)为百合科黄精属多年生草本植物,药用历史有两千多年,俗称姜形黄精,同时还具有食用性。李时珍曰:“黄精为服食要药”,体现其药食两用性[1]。多花黄精以黄精多糖、甾体皂苷为主要有效成分[2],具有补气养阴、益肾填精、生津补脾等功效[3-5]。现代研究发现,黄精有降血糖、血脂、血压和抗衰老的作用,对治疗心血管疾病、糖尿病等方面也有较好效果[6-7]。中国该属植物有30多种。多花黄精主产于四川、湖南、云南等省,是中国的特有植物。
种子休眠是植物种子对外界环境的一种适应机制[8],很多学者对种子休眠原因及打破种子休眠的方法进行了大量的研究[9-16]。发现种子休眠的类型及原因包括两大类:第一类是胚本身的因素导致的。包括胚发育未完成;生理上没成熟;缺少必须的激素或者抑制种子萌发物质的存在。种子需要通过后熟过程,完成一系列生理生化变化后才能萌发。第二类是外源因素导致的,即胚以外的各种组织导致休眠,比如种壳产生的机械阻碍或存在抑制物,种子不透水、不透气等[17-20]。
黄精种子具有休眠现象,研究表明黄精种子休眠属于胚本身原因引起的休眠,即种胚有一个后熟期,也可能是种胚内内源激素GA和CK相对较少,种子处于休眠状态,即使有适宜的环境条件种子仍不能萌发。导致黄精种子休眠的原因包括胚的后熟以及种子结构。
张跃进等研究表明黄精种子属于综合休眠。多花黄精与黄精同为黄精属,其形态特征和内环境等都具有相似性,因此都具有休眠性,发芽率较低。多花黄精单独作为一个种,但关于多花黄精种子繁殖技术的报道较少。张旺凡[21]和赵致等[22]分别利用适宜浓度的外源6-BA和GA3处理和低温贮藏的方法来打破多花黄精种子休眠,都得到较好的发芽效果。刘保才将多花黄精种子在不同条件下贮藏,发现在低温和常温下层积种子打破休眠效果较好[23]。周建金等为打破黄精种子休眠、提高种子发芽率,利用组织培养技术,对多花黄精种子进行不同浓度GA3处理和不同时间的低温贮藏处理[24]。张旺凡认为,多花黄精种子休眠属于胚本身原因引起的休眠,即种胚有一个后熟期[21]。
多花黄精种子具有休眠性,发芽率低,使得多花黄精资源开发利用、人工育种栽培遇到困难。所以本研究探讨了层积后熟过程中多花黄精种子生理生化的变化,通过测定多花黄精种子后熟过程中内源激素及关键酶活性的变化,明确胚发育与内源激素及呼吸途径关键酶的动态变化规律,为实际生产中采用生化手段调控胚休眠,及促进后熟提供理论指导,完善多花黄精种子休眠机制理论。同时为多花黄精种苗育种及资源更合理的利用提供便利。
1 材料与方法
1.1 试验材料
种子采自云南省金平县大老塘种植基地的二年生多花黄精植株,经鉴定为百合科植物多花黄精的果实和种子。采收果实后密封1周揉搓去掉果皮,洗净获得多花黄精种子,用滤纸吸干外表水分备用。实验在西南生物多样性实验室进行。
1.2 试验方法
1.2.1 多花黄精种子后熟期间生理生化指标测定
以干燥贮藏的种子(袋装自然存放)为对照,将种子按20 ℃层积、20 ℃与4 ℃(每24 h交替1次)交替层积、4 ℃层积、-20 ℃层积4种方法处理(将种子与湿砂按体积1∶5层积,河砂湿度手捏刚出水而不落,定期喷水保湿)。以后每10 d测定不同贮藏条件下多花黄精种子后熟期间的下列各项指标,各项测定均重复3次。
种子激素测定方法:脱落酸含量、赤霉素含量、细胞分裂素含量、生长素含量采用酶联免疫分析(ELISA)试剂盒测定。
1.2.2 多花黄精种子发芽实验
以干燥贮藏种子为对照,将种子按20 ℃层积、20 ℃与4 ℃(每24 h交替1次)交替层积、4 ℃层积、-20 ℃层积4种方法处理,贮藏70 d后进行种子萌发实验。
萌发前,种子先于20 ℃培养箱清水浸泡24 h,再播种于内径为15 cm的培养皿内,内放3层滤纸,蒸馏水培养, 25 ℃黑暗条件下智能人工培养箱培养,每处理均重复3次,每重复50粒种子。以种胚突破种皮为标准,间隔20 d统计1次发芽数,共统计3次。
2 结果与分析
2.1 不同贮藏条件下种子萌发情况
多花黄精种子具有休眠特性,需要经过一段时间的层积处理完成后熟才能萌发。作为对照,在干燥贮藏条件下,多花黄精种子层积50 d后生活力完全丧失,种子失去发芽力。与对照相比,层积70 d后,4 ℃层积、交替层积和20 ℃层积这3种贮藏条件下多花黄精种子的生活力分别为72.33%、70.67%和 70.67%,将不同贮藏条件下的种子分别置于25 ℃黑暗条件并给予一定水分之后,20 d后种子陆续开始萌发,萌发率分别为62%、59%和55%,4 ℃条件下萌发率最高;而-20 ℃贮藏条件下的种子,层积70 d后,种子生活力降为0,将种子置于25 ℃并给予一定水分之后,未能萌发。
表1 不同贮藏条件下种子生活力和发芽率
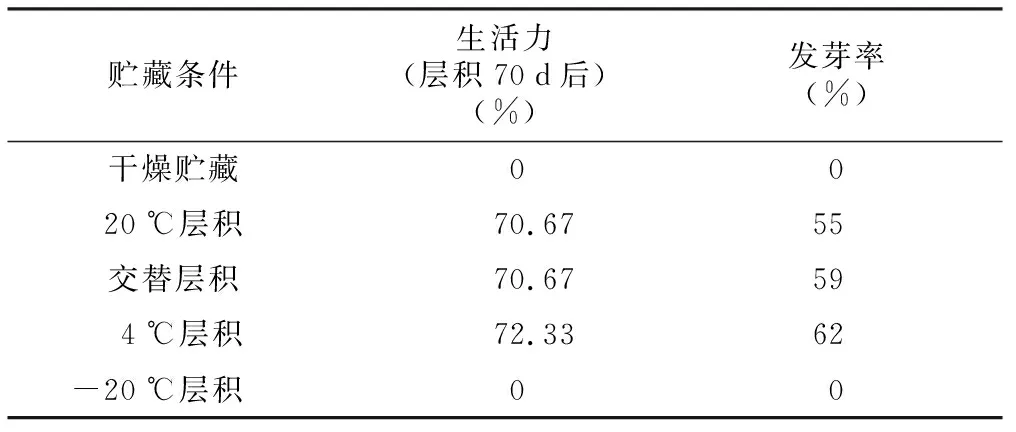
贮藏条件生活力(层积70d后)(%)发芽率(%)干燥贮藏0020℃层积70.6755交替层积70.67594℃层积72.3362-20℃层积00
2.2 不同贮藏条件下种子内源激素含量的变化
GA更多的被认为是促进种子解除休眠的必要因子,GA主要通过2种途径来促进种子解除休眠: 1) 动员种子内的贮藏物质,刺激胚的生长; 2) 増强软化屏障组织水解酶的表达,降低根尖的机械阻力,促进胚根突破种皮。
在多花黄精后熟过程中,4 ℃、交替和20 ℃ 3种层积方式的GA3含量都随时间的推移表现为先降低后升高(如图1),其中4 ℃层积和20 ℃层积条件下种子在30 d时降为后熟过程中最低,分别为0.523 pmol/g和0.614 pmol/g;交替条件种子在40 d时达最低值(为0.621 pmol/g)。如图1表明,层积处理对赤霉素无显著影响。
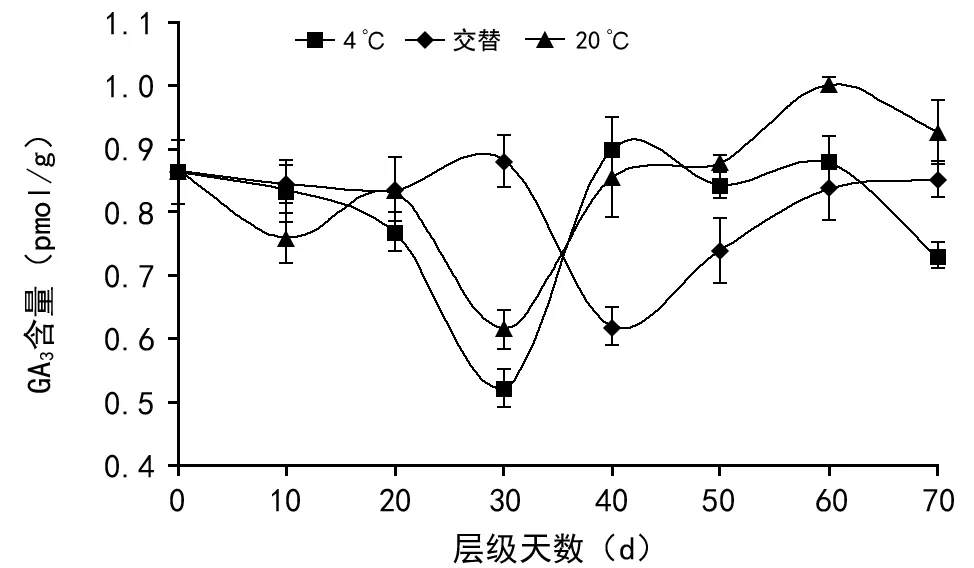
图1 不同贮藏条件下多花黄精种子后熟期间赤霉素含量的变化
在种子后熟过程中(如图2), 随着贮藏时间延长,4 ℃条件下的IAA含量随时间的推移表现为先降低后升高再降低的趋势,在30 d和40 d分别达到整个后熟过程中的最低值和最高值,分别为0.141μmol/g和0.201μmol/g,后持续下降直到层积结束;20 ℃条件下,种子在前40 d 有小幅度的升降,为0.18μmol/g,在50~70 d开始下降,70 d时为0.165μmol/g;交替条件下,在10 d时IAA含量由最初的0.175μmol/g降低为0.154μmol/g,在20 d时上升后再下降,在40 d时达到后熟过程中的最低值0.123μmol/g,后有小幅升高直至层积结束,为0.131μmol/g。4 ℃条件下的波动显著可能说明4 ℃条件下IAA参与了种子后熟某些机制的调控,或者是对ABA的抑制并且反应活跃。
在种子后熟过程中(如图3),在20 ℃层积的贮藏条件下,层积20 d后CTK含量从191.21 ng/g 上升为267.00 ng/g,30 d快速下降为188.36 ng/g,随着时间小幅变化,70 d时上升为288.03 ng/g;在20 ℃层积、4 ℃层积这2种贮藏条件下,种子的CTK含量变化趋势基本一致,均随着层积时间的延长而表现出先降低后升高再降低的趋势,不同的是,不同贮藏条件下种子CTK含量在整个层积过程中达到峰值的时间先后略有差异,在4 ℃条件下,CTK含量出现2个峰值,层积30 d后达到第1个峰值,为293.73 ng/g,之后逐渐下降,层积50 d后有小幅回升,到层积60 d时达到第2个峰值,为301.32 ng/g,之后逐渐下降直到层积结束;在20 ℃条件下,2个双峰出现在20 d和40 d,分别为267.00 ng/g和285.13 ng/g,随后又开始下降再升高,到70 d层积结束时为 288.03 ng/g。
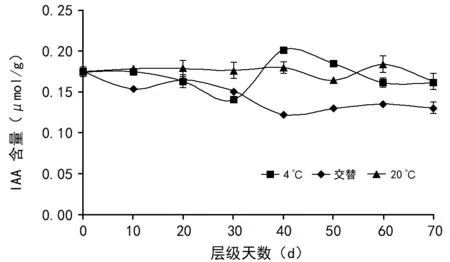
图2 不同贮藏条件下多花黄精种子后熟期间生长素含量的变化

图3 不同贮藏条件下多花黄精种子后熟期间细胞分裂素含量的变化
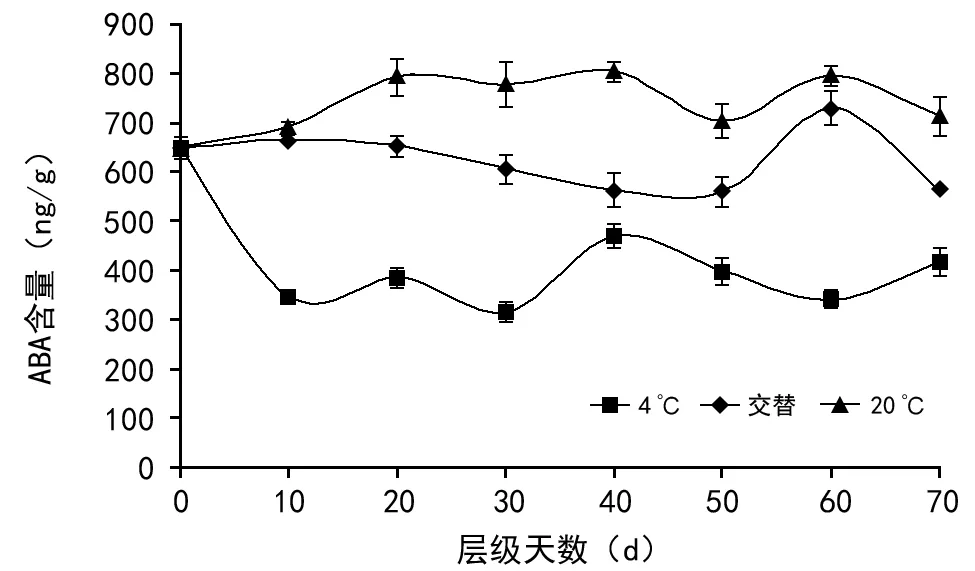
图4 不同贮藏条件下多花黄精种子后熟期间脱落酸含量的变化
在种子后熟过程中(如图4),在3种不同贮藏条件下,ABA含量差异明显。在4 ℃层积贮藏条件下,层积10 d后ABA含量从651.23 ng/g 下降为345.74 ng/g,随着时间小幅变化,40 d时上升为469.82 ng/g,随后下降再上升,层积结束时为417.20 ng/g;在20 ℃层积贮藏条件下,种子的ABA含量随着层积时间的延长而表现出先小幅度升高后下降再升高,50 d时为后熟过程中最小值,为703.85 ng/g,层积结束时为714.06 ng/g;在交替层积贮藏条件下,随着层积时间的延长而表现出先小幅度降低后在50 d快速升高,在60 d达到最大值,为795.73 ng/g,后下降,层积结束时为714.06 ng/g。
3 讨 论
多花黄精种子具有胚后熟的习性,发芽率低,对多花黄精种子育苗与育种工作的开展造成阻碍,但目前有关多花黄精种胚后熟的机制方面的研究较少。通过对多花黄精种胚后熟过程中ABA、GA3、IAA和CTK等几种主要内源激素含量的变化规律研究,探索缩短或延长种胚发育时期的方法和途径,为种胚后熟机理的深入研究和生产中的种苗繁殖提供依据。
3.1 GA3, IAA对多花黄精种子后熟的调控作用不明显
在多数休眠种子中,GA3含量和ABA含量之间的平衡关系是种子破除休眠的关键条件。但是在本实验的多花黄精种子层积过程中,种子内检测到的GA3和IAA含量都非常低,且变化不明显,可以推测多花黄精种子休眠解除与GA3和IAA水平的变化关系不大。GA3在整个层积过程含量较低,变化不明显,一直维持在0.5~1.0 pmol/g之间,凡是能够正常发芽的种子均存在具有生理活性的赤霉素,但是存在生理活性的种子不一定能发芽[25],仍需要满足其他利于种子发芽的生理条件,说明赤霉素可能不是打破多花黄精种子休眠主要因素,这与史静等[26]的研究结果类似。另一方面,考虑到GA3和IAA难提取和易分解的特性,故不能排除在本实验中GA3和IAA因提取和保藏方法不当,可能造成检查结果存在偏差。
3.2 ABA、CTK可能参与了多花黄精种子的后熟生理调控
多花黄精种子在层积处理0~70 d中,在4 ℃贮藏条件下,CTK的含量表现出先升高后下降再升高再下降的趋势,出现了2个峰值,分别为第30天和第60天,含量分别为293.73 ng/g和301.32 ng/g,而ABA含量在贮藏30 d和60 d时为后熟过程中的2个谷底值,可以发现CTK和ABA含量的变化趋势相反,说明这2种激素在调控着种子的休眠与萌发;同样在20 ℃层积和交替层积贮藏条件下,在第60天,种子CTK含量出现迅速下降的同时发现ABA的含量出现上升的情况,再次说明CTK和ABA这2种激素在互相影响,调控着种子的休眠与萌发。故推测CTK和ABA可能参与了多花黄精种子胚后熟生理调控。在肉苁蓉种子层积处理破除休眠的研究[27-28]中也曾发现细胞分裂素含量的显著升高,并指出细胞分裂素促进种子萌发相关酶活性的增强,还可能对拮抗种子内含萌发抑制物有利。试验还发现,在层积过程中,与其它2种贮藏条件相比,4 ℃贮藏条件下的ABA的含量最低,CTK含量也相对较高,可能表示4 ℃条件下进行层积处理更适于多花黄精种子打破休眠,这与种子萌发试验中4 ℃贮藏条件下种子萌发率最高的结果相一致。
3.3 多花黄精种子在胚后熟过程中的关键期
在20~40 d期间,多花黄精种子激素CTK/ABA含量的比值在整个层积过程中表现出升高的现象,有利于种子萌发。因此可以认为,在整个层积处理过程中,20~40 d是多花黄精种子打破休眠的一个关键期。
4 结 论
本研究通过对不同贮藏条件下多花黄精种子植物激素含量及种子成活率的动态测定,结果表明:
多花黄精种子层积第20~40天是胚后熟的关键期,种子代谢最为活跃。
4 ℃层积贮藏最有利于多花黄精种子萌发,发芽率达72.33%。超低温条件贮藏不利于种子生活力的保持。
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
中老年保健(2021年11期)2021-08-22
花卉(2020年24期)2021-01-04
防护林科技(2020年11期)2020-12-30
东北农业大学学报(2020年11期)2020-12-17
Digital Chinese Medicine(2020年3期)2020-11-03
云南农业科技(2020年5期)2020-10-14
西南农业学报(2019年1期)2019-03-01
中国食品学报(2019年3期)2019-01-13
天津农业科学(2018年3期)2018-03-24
