
不同比例的生物饲料对罗氏沼虾生长、消化酶和免疫酶活性的影响
2018-12-29 07:27杨树浩陈建酬洪允成
饲料工业 2018年2期
■杨树浩 冯 艺 陈建酬* 洪允成
(1.佛山科学技术学院动物营养实验室,广东佛山528000;2.佛山市三水白金水产种苗有限公司,广东佛山528000)
罗氏沼虾又称马来西亚大虾,产自厄瓜多尔沿岸附近,是一种生长发育快、食性杂、体格大、肉质鲜美、经济实惠的大型淡水虾。20世纪90年代以来,全球产量持续增长[1]。到2016年上半年,我国罗氏沼虾养殖主产区分布在沿海省市,养殖面积达到60万亩,占全国养殖产值的1.5%,是我国主要经济虾类之一[2]。生物饲料通过微生物发酵剂菌种的发酵能消除饲料中抗营养因子,分解原料大分子蛋白,产生多种有机酸[3],具有改善饲料品质、促进动物生长、提高饲料消化率、增强免疫力等作用[4]。近年来已应用在克氏原螯虾(Procambarus clarkii)[5]、中华绒螯蟹(Eriocheir sinensis H.Milne-Edwards)[6]等水产动物中,能够促进动物生长,提高机体免疫酶活性,在罗氏沼虾养殖中尚未见报道。本试验给罗氏沼虾投喂不同比例的生物饲料,探究生物饲料对罗氏沼虾生长、消化酶和免疫酶活性的影响,为生物饲料推广应用提供参考。
1 材料与方法
1.1 试验材料
胰蛋白酶(TPS)、脂肪酶(LPS)、淀粉酶(AMS)、纤维素酶(CL)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、碱性磷酸酶(AKP)、酸性磷酸酶(ACP)、谷丙转氨酶(GPT)和谷草转氨酶(GOT)测定试剂盒,均购于南京建成生物工程研究所。
1.2 试验动物及饲料
试验用的罗氏沼虾来自于佛山市三水白金水产种苗有限公司,在水泥池暂养一周,投喂广东旺海饲料实业有限公司生产的基础饲料。
基础饲料和生物饲料的成分分别见表1和表2。生物饲料∶水=2∶5混合,常温密封发酵72 h,室内低温干燥后,按饲养试验要求与基础饲料充分混合粉碎过60目筛,4℃保存备用。

表1 基础饲料的组成(干物质,%)

表2 生物饲料的组成(干物质,%)
1.3 试验设计及饲养管理
选择平均体重为(0.02±0.01)g健康的罗氏沼虾28 800尾,按饲养试验要求随机分为4组,每组设3个重复,每个重复2 400尾虾,分别饲养在水泥池中(5.3 m×3.5 m×1 m),池底设有微孔增氧和换水系统,放置遮蔽物,防止沼虾自相残杀。投喂4种饲料:对照组为100%基础饲料(A组),B、C、D组分别为90%基础饲料+10%生物饲料、80%基础饲料+20%生物饲料、70%基础饲料+30%生物饲料。养殖期间水温为23~28 ℃,溶解氧约为6.3 mg/l,pH值约为8.0,氨氮约为0.1 mg/l,亚硝酸盐约为0.1 mg/l。每天上下午各投喂一次,日投喂量为虾体重的5%。每天清理一次池底的残饵和排泄物,每3 d更换池水1/3,试验期为42 d。
1.4 生长性能的测定
饲养结束时计数各组活虾数,每组随机挑选300尾罗氏沼虾测量体长(精确到0.01 cm)和体重(精确到0.01 g),计算下列指标:
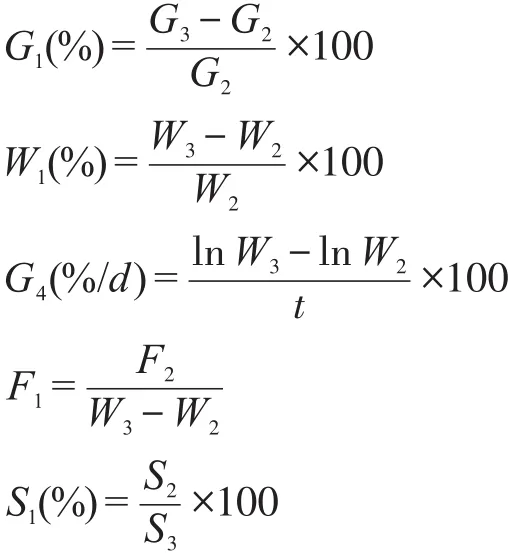
式中:G1——增长率(%);
G2——初体长(cm);
G3——末体长(cm);
W1——增重率(%);
W2——初体质量(g);
W3——末体质量(g);
G4——特定生长率(%/d);
t——饲养时间(d);
F1——饲料系数;
F2——投饵量(g);
S1——存活率(%);
S2——存活数(尾);
S3——总数(尾)。
1.5 肌肉营养指标测定
饲养结束后,将罗氏沼虾饥饿1 d。每组随机挑选500尾罗氏沼虾,在冰水浴中去除虾壳和内脏,取躯体肌肉样本,测定体成分。肌肉中水分含量按GB/T 6435—2014测定(直接干燥法),粗蛋白质含量按GB/T6432—94测定(仲裁法),粗脂肪含量按GB/T 6433—2006测定(索氏提取法),粗灰分含量按GB/T 6438—2007测定,肌肉氨基酸含量采用S-433D型高效液相色谱仪测定。
参考氨基酸评分标准模式[7]和全鸡蛋蛋白质的氨基酸模式[8],分别计算氨基酸评分(AAS)、化学评分(CS)和必需氨基酸指数(EAAI)。
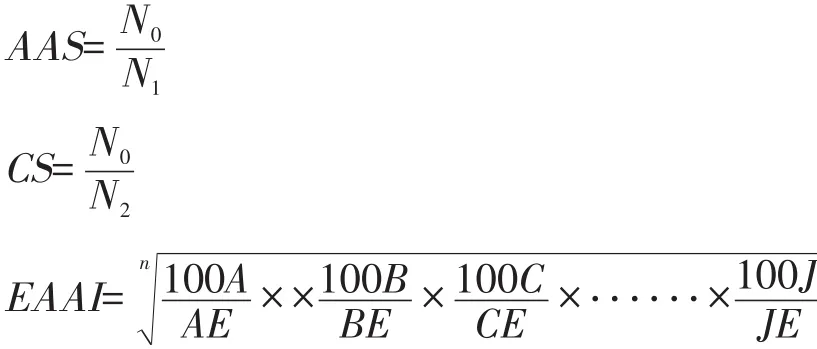
式中:N0——样品氨基酸含量(%);
N1——标准模式氨基酸含量(%);
N2——全鸡蛋蛋白质同类氨基酸含量(%);
n——必需氨基酸数量(个);
A、B、C…J——样品蛋白质的必需氨基酸含量(%);
AE、BE、CE…JE——全鸡蛋蛋白质同类的氨基酸含量(%)。
1.6 消化酶和免疫酶活性的测定
每组随机挑选500尾罗氏沼虾,取肝胰脏[9],依照试剂盒说明书的方法,冰水浴匀浆、离心后,采用试剂盒测定酶活性,测定仪器为紫外分光光度计。
1.7 数据处理
结果以“平均值±标准差(X±SD)”表示,原始数据采用Excel 2007软件进行统计,应用SPSS 22.0软件中“单因素(ANOVA)方差分析”,利用Duncan's法对数据进行多重比较,P>0.05为差异不显著,P<0.05为差异显著,P<0.01为差异极显著。
2 结果与分析
2.1 不同比例的生物饲料对罗氏沼虾生长性能的影响(见表3)
由表3可知,C组罗氏沼虾的增重率为296.34%,极显著高于A组(P<0.01),显著高于B组(P<0.05),增重率分别比A组、B组提高了7.42%和3.75%;D组罗氏沼虾的增重率为288.59%,显著高于A组(P<0.05),增重率提高了4.61%;B组罗氏沼虾的增重率与A组相比差异性不显著(P>0.05)。罗氏沼虾的增长率、特定生长率、饲料系数和存活率各组间的差异不显著(P>0.05)。
2.2 不同比例的生物饲料对罗氏沼虾肌肉营养指标的影响(见表4)

表3 不同比例的生物饲料对罗氏沼虾生长性能的影响

表4 不同比例的生物饲料对罗氏沼虾肌肉营养指标的影响(湿样,%)
由表4可知,不同比例的生物饲料对罗氏沼虾肌肉营养指标影响不显著(P>0.05)。其中C组罗氏沼虾的粗蛋白质含量最高,为19.00%,水分含量最低,只有78.50%;D组仅次之,粗蛋白质为18.96%、水分为78.53%;A组罗氏沼虾的粗蛋白质含量最低,只有18.44%,水分含量最高,达到了79.02%。四组罗氏沼虾的粗脂肪和粗灰分含量差异不大。
2.3 不同比例的生物饲料对罗氏沼虾肌肉氨基酸含量的影响
罗氏沼虾肌肉中氨基酸组成见表5,除色氨酸(Try)在水解过程中被破坏无法测定外,共含有17种氨基酸,包含9种必需氨基酸(EAA)和4种呈味氨基酸(DAA)。四组罗氏沼虾肌肉的总氨基酸(TAA)含量无显著性差异(P>0.05),其中C组总氨基酸、必需氨基酸、呈味氨基酸含量达到最高,分别是16.05%、7.02%、6.02%,D组次之,A组总氨基酸、必需氨基酸和呈味氨基酸含量最低。按照世界卫生组织/联合国粮农组织(WHO/FAO)对理想蛋白的定义,罗氏沼虾肌肉中EAA含量/TAA含量超过40%,EAA含量/NEAA含量超过60%,属于标准的蛋白质源。谷氨酸(Glu)含量占TAA含量的比例最高,Glu含量/TAA含量达到14%,赖氨酸(Lys)含量占EAA含量的比例最高,Lys含量/EAA含量接近22%。
罗氏沼虾肌肉氨基酸评分、化学评分和必需氨基酸指数评定结果见表6和表7。肌肉中必需氨基酸的氨基酸评分接近或超过1,化学评分接近或超过0.5,基本符合WHO/FAO理想蛋白的标准。各组罗氏沼虾肌肉氨基酸的必需氨基酸指数为67.43~70.80,赖氨酸的氨基酸评分和化学评分分别为1.43~1.48和1.10~1.14。蛋氨酸+胱氨酸(Met+Cys)的氨基酸评分和化学评分最低,分别只有0.71~0.76和0.40~0.43。
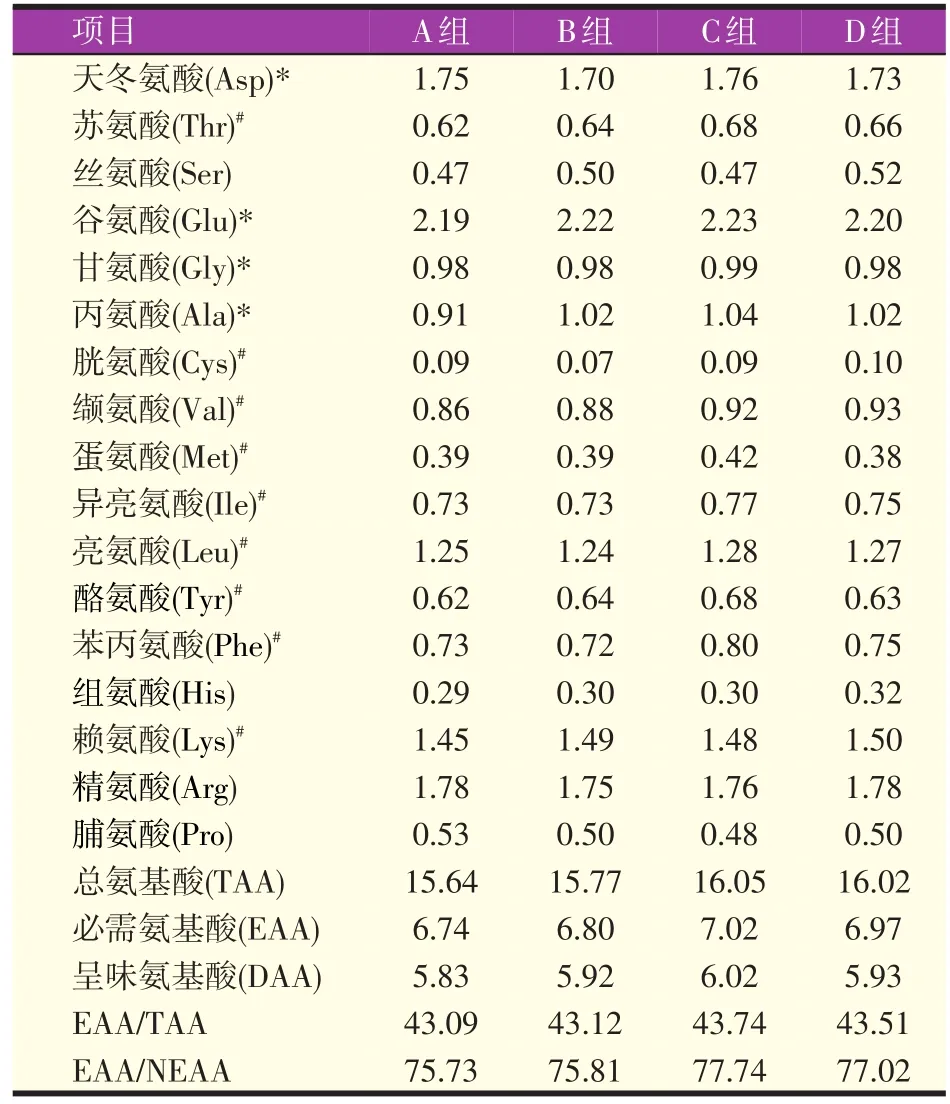
表5 不同比例的生物发酵饲料对罗氏沼虾肌肉氨基酸组成的影响(湿样,%)

表6 罗氏沼虾肌肉必需氨基酸分数与两个评分模式(mg/gN)

表7 罗氏沼虾肌肉必需氨基酸组成评分
2.4 不同比例的生物饲料对罗氏沼虾肝胰脏消化酶活性的影响(见表8)
由表8可知,B组罗氏沼虾胰蛋白酶的活性为130.81 U/mg prot.,极显著高于A组(P<0.01),C组罗氏沼虾胰蛋白酶的活性高于其他三组,达到了145.07 U/mg prot.,极显著高于A组(P<0.01),显著高于B组(P<0.05),D组罗氏沼虾胰蛋白酶的活性为135.30 U/mg prot.,极显著高于A组(P<0.01)。

表8 不同比例的生物饲料对罗氏沼虾肝胰脏消化酶活性的影响
C组罗氏沼虾淀粉酶的活性为2.12 U/mg prot.,极显著高于A组(P<0.01),显著高于B组(P<0.05);D组罗氏沼虾淀粉酶的活性为2.81 U/mg prot.,极显著高于其他三组(P<0.01)。纤维素酶的活性只有C组罗氏沼虾最好,为3.06 U/mg prot.,极显著高于A组(P<0.01),显著高于B组(P<0.05)。四组罗氏沼虾的脂肪酶活性差异不显著(P>0.05)。
2.5 不同比例的生物饲料对罗氏沼虾肝胰脏免疫酶活性的影响(见表9)
由表9可知,C组罗氏沼虾超氧化物歧化酶的活性为17.76 U/mg prot.,极显著高于A组(P<0.01),显著高于B组(P<0.05);D组罗氏沼虾超氧化物歧化酶的活性最高,达18.10 U/mg prot.,极显著高于A组和B组(P<0.01)。C组罗氏沼虾谷草转氨酶的活性为35.64 U/mg prot.,显著高于A组(P<0.05)。四组罗氏沼虾过氧化氢酶、碱性磷酸酶、酸性磷酸酶和谷丙转氨酶的活性差异不显著(P>0.05)。
3 讨论
3.1 不同比例的生物饲料对罗氏沼虾生长的影响
研究发现,发酵菌的发酵作用有效降解了饲料中抗营养因子,饲料中淀粉、粗蛋白质等大分子物质被水解成易被动物体吸收的各种小肽和氨基酸[10],发酵产物含有大量的营养物质[11]、外源性消化酶[12]和有益菌落[13],为机体合成生命所需物质提供充足的基础原料,补充消化道的消化酶和有益菌落,提高机体对饲料的消化能力,尤其是对罗氏沼虾难以消化的淀粉和纤维素进行分解,提高饲料利用率,降低成本,促进罗氏沼虾的生长发育。沈城等[14]试验发现,发酵豆粕代替15%~20%的鱼粉时幼蟹的体重指标有显著性的提高。饵料中添加适量发酵豆粕能促进岩鱼(Sebastes schlegeli)的生长发育[15],在饵料中添加植物乳酸菌发酵的大豆粉显著提高了大菱鲆(Scophthalmusmaximus L.)的生长[16]。
在本试验中,罗氏沼虾的增重率随生物饲料添加量的增加而增加,基础饲料中添加20%生物饲料时,罗氏沼虾增重率与对照组相比提高了7.42%;说明生物饲料与发酵豆粕一样能提高动物的增重率。有研究显示,随着发酵饲料添加比例的增大,动物的增重率和增长率开始下降[17]。发酵饲料具有易消化吸收、含多种促生长因子和一定诱食性等特点,但其经过微生物的消化吸收后,其产物营养价值不如常规的原料,添加量过多会导致饲料的营养水平低于动物的营养需求,影响动物的生长发育。本试验中添加30%生物饲料投喂后,罗氏沼虾增重率与对照组相比提高了4.61%,低于生物饲料添加比例为20%的罗氏沼虾的增重率。可见,在基础饲料中添加适量的生物饲料能有效提高罗氏沼虾增重率,而添加比例过多时动物的生长发育减缓。
在本试验中,投喂生物饲料对罗氏沼虾体长、特定生长率、饲料系数和存活率影响不明显,这可能与发酵产物对不同生长指标的促进作用不同有关,其作用机理还有待进一步研究。
3.2 不同比例的生物饲料对罗氏沼虾肌肉营养指标的影响
测量动物体肌肉营养成分能够准确判断动物对营养物质的需求和评估肌肉的营养价值,对生产具有指导意义。周兴华等[18]发现,添加不同比例的发酵豆粕对鲫鱼(Carassius auratus)肌肉营养成分没有改善效果,添加比例过多反而会降低肌肉粗蛋白质含量、提高肌肉水分;沈城等[14]研究表明,少量发酵豆粕代替鱼粉对中华绒螯蟹的肌肉粗蛋白质、粗脂肪和粗灰分无显著影响。王天神[5]通过饲养试验发现,生物发酵饲料对提高虾体肌肉氨基酸差异不显著。本试验结果显示,基础饲料中添加生物饲料对罗氏沼虾的肌肉营养成分的影响差异不显著,粗蛋白质含量随生物饲料添加比例的递增而先增加后降低。当添加20%生物饲料时,罗氏沼虾粗蛋白质含量最高为19.00%,其余试验组的罗氏沼虾粗蛋白质含量均高于对照组。投喂生物饲料的罗氏沼虾肌肉含水量比投喂基础饲料的罗氏沼虾低,而粗脂肪和粗灰分含量变化无明显规律。
罗氏沼虾肌肉中必需氨基酸分数基本符合WHO/FAO理想蛋白模型,必需氨基酸指数达到67.43~70.80,其中赖氨酸含量丰富,约占必需氨基酸总量的22%,恰好能补充人类谷类食品缺乏的赖氨酸,而罗氏沼虾肌肉中蛋氨酸和胱氨酸的氨基酸评分和化学评分分别只有0.71~0.76和0.40~0.43,达不到WHO/FAO理想蛋白模型,可见蛋氨酸和胱氨酸是罗氏沼虾的限制性氨基酸。
3.3 不同比例的生物饲料对罗氏沼虾肝胰脏消化酶活性的影响
罗氏沼虾的消化能力与体内酶活性有密切关系。发酵菌在发酵过程需要分泌多种消化酶,如胰蛋白酶、淀粉酶和纤维素酶等,保持很长时间的高活性。生物饲料中的消化酶能作为动物的外源性酶,进入动物肠道帮助动物消化吸收[13]。李程琼等[19]在基础饲料中添加21%的发酵豆粕饲养鳗鱼(Ell),发现其肠道消化酶活性有显著提高;彭翔等[20]利用发酵豆粕代替10%~20%的鱼粉进行饲喂能显著提高黑鲷(Acanthopagrus schlegelii)体内部分消化酶的活性;杨霞等[21]利用发酵棉籽粕饲喂中华绒蟹幼蟹也得出同样结论。这说明添加发酵饲料进行投喂能提高动物的消化酶活性。
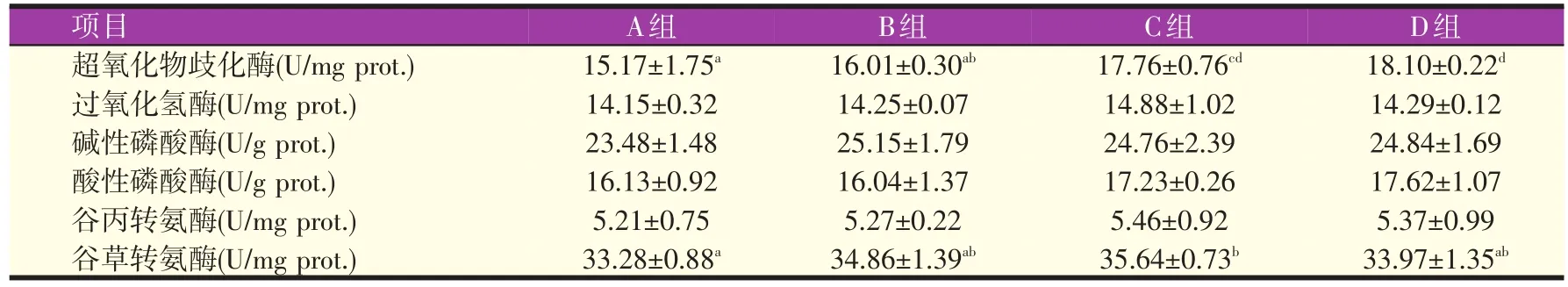
表9 不同比例的生物饲料对罗氏沼虾肝胰脏免疫酶活性的影响
在本试验中,罗氏沼虾胰蛋白酶和纤维素酶的活性随着生物饲料添加比例的递增而呈现不同程度的增强。当基础饲料中添加10%生物饲料时,罗氏沼虾胰蛋白酶和纤维素酶的活性明显提高。而添加比例到达20%时,罗氏沼虾胰蛋白酶和纤维素酶的活性分别比对照组提高了45.52%和18.60%。生物饲料与其他发酵饲料对消化酶活性的影响一致,能提高罗氏沼虾蛋白酶和纤维素酶的活性。随着生物饲料添加比例不断增加,消化酶活性呈下降趋势,吕云云等[17]、冯建等[22]、李云兰等[23]分别在圆斑星鲽(Verasper variegatus)、大黄鱼(Larimichthys crocea R.)和鲤鱼(Cyprinus carpio)试验中得到验证。生物饲料含有大量的外源性消化酶,在进入动物体内后和消化道的消化酶混合在一起对饲料进行消化吸收,增强胃肠道的排空率。当基础饲料中添加30%生物饲料时,胰蛋白酶和纤维素酶的活性分别为135.30 U/mg prot.和 2.82 U/mg prot.,有明显下降趋势。机体的消化道的消化速率加快,机体营养需求得到快速的补充,不再需要过多的消化酶参与消化,通过神经调节减少肝胰脏消化酶的合成[24],导致消化酶活性下降。而罗氏沼虾淀粉酶活性随着生物饲料比例递增而平稳上升,基础饲料中添加10%生物饲料对罗氏沼虾淀粉酶活性效果不显著,随着添加比例的增加,基础饲料中添加20%和30%生物饲料投喂后,罗氏沼虾淀粉酶活性出现显著差异,最高时比对照组淀粉酶活力提高了77.85%。本试验生物饲料的添加比例最高只添加到30%,未能探究出淀粉酶活力最高时生物饲料的最佳添加量,相关研究有待进一步完善。
有研究发现,鳙鱼(Aristichthys nobilis)肝胰脏、匙吻鲟(Polyodon spathula)肠道和肝脏消化酶与其体长体重呈显著的正相关[25]。本试验中,基础饲料中添加20%生物饲料显著提高了罗氏沼虾胰蛋白酶和纤维素酶活性,增长率比对照组提高了7.42%,说明生物饲料提高了罗氏沼虾胰蛋白酶和纤维素酶活性而提高其增长率,而添加30%生物饲料时,增长率只比对照组提高了4.61%,这与胰蛋白酶和纤维素酶活性较低有关。而鲢鱼和匙吻鲟消化道和肝胰脏的脂肪酶与其体长体重相关性不强,甚至呈负相关[25]。本试验中生物饲料对罗氏沼虾肝胰脏的脂肪酶活性有增强作用但没有显著性差异,这可能与生物种类不同有关。
3.4 不同比例的生物饲料对罗氏沼虾肝胰脏免疫酶活性的影响
抗氧化酶和代谢酶活性水平直接影响水产动物抗病力,两者相互协同构成一套生物体保护的机制[26]。发酵菌发酵能有效的消除饲料中的抗营养因子,减少硫化氢、二氧化碳等有害气体在肠道消化过程的产生,避免机体受到刺激产生过敏反应[27]。生物饲料发酵后包含多种益生菌,能在动物体肠道中创造酸性环境,抑制病原菌的生长发育,同时能产生多种氨基酸和小肽等物质,增强机体免疫系统[13]。夏青等[28]利用生物饲料饲喂凡纳滨对虾(Litopenaeus vannamei),虾体内超氧化物歧化酶活性显著提高。徐奕晴等[6]在基础饲料中添加15%发酵料投喂中华绒螯蟹,发现能显著提高幼蟹的抗氧化酶活性。而孙宏等[29]利用发酵棉籽粕代替鱼粉饲喂黑鲷后谷草转氨酶活性有所提高,但差异不显著。本试验中,随着生物饲料添加比例的增加,超氧化物歧化酶和谷草转氨酶活性也有不同程度的增强。当基础饲料中添加20%生物饲料时,罗氏沼虾超氧化物歧化酶和谷草转氨酶的活性分别比对照组提高了17.07%和7.09%,说明饲喂生物饲料后有益菌在肠道占领主要地位,提高了动物体免疫水平,与此同时谷草转氨酶活性增强,罗氏沼虾体内蛋白质合成能力增加,分解降低,有助于氮在体内沉积,提高动物肌肉粗蛋白质含量,达到了19.00%,但相对于其他组差异不显著,这可能与其他因素有关。当基础饲料中添加30%生物饲料时,谷草转氨酶的活力出现下降趋势,与对照组相比差异不显著,肌肉粗蛋白质含量也下降,这可能是因为生物饲料的添加比例过高,饲料营养水平不能满足机体的需求,导致蛋白沉淀下降。而罗氏沼虾超氧化物歧化酶活性却提升到18.10 U/mg prot.,比对照提高了19.31%,说明生物饲料的添加比例越高,微生物数量越多,抗活性氧能力越高,进一步增强动物机体免疫能力。本试验生物饲料的添加比例最高只添加到30%,未能探究出超氧化物歧化酶活性最高时生物饲料的最佳添加量,相关研究有待进一步完善。
4 结论
①基础饲料中添加10%~30%生物饲料能明显提高罗氏沼虾肝胰脏胰蛋白酶的活性。
②基础饲料中添加20%~30%生物饲料能显著提高罗氏沼虾增重率、肝胰脏淀粉酶、纤维素酶和超氧化物歧化酶的活性。
③基础饲料中添加20%生物饲料能显著提高罗氏沼虾肝胰脏谷草转氨酶的活性。
猜你喜欢
当代水产(2022年7期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2021年9期)2021-12-02
当代水产(2021年8期)2021-11-04
当代水产(2021年8期)2021-11-04
医学前沿(2021年18期)2021-04-14
海洋与渔业(2017年11期)2018-01-26
中国民族民间医药·下半月(2014年4期)2014-09-26
