
长江湖口段春夏季仔稚鱼群落结构研究
2019-01-07 11:17杨习文李新丰方弟安徐东坡
水生生物学报 2019年1期
刘 熠 杨习文 任 鹏 李新丰 方弟安, 徐东坡,
(1. 上海海洋大学水产科学国家级实验教学示范中心, 上海 201306; 2. 中国水产科学研究院淡水渔业研究中心, 农业农村部长江下游渔业资源环境科学观测实验站, 无锡 214081)
鱼类早期生活史阶段是鱼类生存繁衍过程中的决定性环节之一, 这个阶段的鱼类存活率能直接影响鱼类群体的数量水平及年龄组成结构[1]。仔稚鱼阶段是鱼类早期生活史阶段的重要组成部分, 对仔稚鱼群聚特征的研究能了解仔稚鱼种群数量在自然环境下的动态变化过程, 评估可开发利用鱼类的种群数量并探索出鱼类在仔稚鱼阶段更适宜存活的环境条件, 进而在渔业政策中制定合理策略对渔业资源进行开发利用和保护[2]。长江流域面积广泛, 水系支流众多, 拥有丰富的鱼类资源[3], 沿长江干流分布着各种鱼类的产卵场和育幼场。为深入了解长江鱼类资源状况, 诸多学者对长江鱼类早期资源做了大量的研究, 最早可追溯到1959年王昌燮等[4]对长江中游江段鱼苗进行了种类鉴定; 1982年长江四大家鱼产卵场调查队等[5]对葛洲坝水利枢纽工程截流后长江四大家鱼产卵场情况进行了调查;1984年余志堂等[6]对铜鱼和圆口铜鱼的早期发育进行了研究; 1988年易伯鲁等[7]对长江干流四大家鱼产卵场的分布、规模和自然条件进行了深入调查;2005年乔晔[8]对长江各种鱼类早期形态发育与种类鉴别做了详细的描述。近年来, 唐锡良等[9]、熊飞等[10]分别调查了长江上游江津江段和宜宾江段的鱼类早期资源现状; 黎明政等[11]、李世建等[12]分别分析了长江中游武穴江段与监利江段的仔稚鱼时空分布; Ren等[13]调查研究了长江下游安庆、南京、靖江3个江段鱼类早期资源的分布特征及周年动态; 钟俊生等[14]研究了长江口沿岸碎波带仔稚鱼种类组成及其季节性变化。这些研究为揭示长江流域鱼类早期资源的基本面貌以及保护长江鱼类产卵场、育幼场提供了现实依据。然而, 基于湖口江段早期资源的研究仅见零星报道。
江湖连通系统对四大家鱼的繁殖和育幼过程可能发挥着重要作用, 使长江下游靠近鄱阳湖的江段四大家鱼仔鱼丰度越高[13,15]。湖口江段使长江与鄱阳湖连通[16], 是鄱阳湖与长江下游进行生态联系的首要江段, 其水域生境复杂, 为多种江湖洄游型鱼类提供了关键洄游通道[17]。对湖口江段渔业资源的研究有助于维护江湖复合生态系统的连通性和完整性[18], 近年关于湖口水域的鱼类群落特征、鱼类资源现状以及水文情势已有研究[19—21], 鄱阳湖通江水道部分鱼类的生长特性和四大家鱼幼鱼的研究也见报道[22—24], 但对长江湖口江段仔稚鱼群落特征的研究确未见报道。随着湖口县经济地带不断延伸, 江段沿岸工厂建设分布密集, 水质污染加剧, 渔业捕捞力度也不断加大, 过大的水域生境压力致使渔业资源明显下降[25]。为此, 本研究采用鱼类早期资源监测的科学方法, 首次在长江中下游鱼类繁殖期内对湖口江段系统开展鱼类仔稚鱼资源的调查研究, 分析湖口江段的仔稚鱼群落特征,并结合水文环境, 解析该江段仔稚鱼与环境因子的关系, 为长江中下游鱼类资源的保护和生态调度策略提供参考。
1 材料与方法
1.1 调查时间与地点
2016年5月18日至8月15日于长江湖口江段开展早期资源调查, 共计调查88d。采样点(E:116°16′46.04″, N: 29°47′28.84″)设在长江与鄱阳湖的交汇口往下7 km处(图 1)。
1.2 调查方法和样品的保存与鉴定
早期资源采样使用主动网具和定置网具。主动网具为0.5 mm网目的圆锥状浮游生物网, 网长2 m, 网口面积为0.50 m2, 锥形网尾部连有一个圆柱型的鱼苗收集装置, 网口固定一个螺旋桨式的流速仪用于测量网口流速。定置网具为弶网, 后面连接着一个可拆卸的网箱用于收集鱼苗[13]。
在调查期间, 定量采集时间为每日8:00至12:00, 定性采集时间为每日16:00至次日8:00。分别于湖口江段南岸、江心、北岸设置3个采样点(图 1), 采样同时记录每次拖网时间, 每个采样点的水温、透明度、流速仪转数等数据。所采集仔稚鱼样品用试剂保存带回实验室分析鉴定, 主动网具采集样品使用甲醛浓度为5%的福尔马林溶液保存,定置网具采集样品的10%使用冰水保存, 其余部分使用甲醛浓度为5%的福尔马林溶液保存, 所有样品鉴定完成后使用75%的中性酒精保存。仔稚鱼种类的鉴定按照曹文宣等[1]编写的《长江鱼类早期资源》进行, 对于难以鉴定确认的物种使用分子方法鉴定。
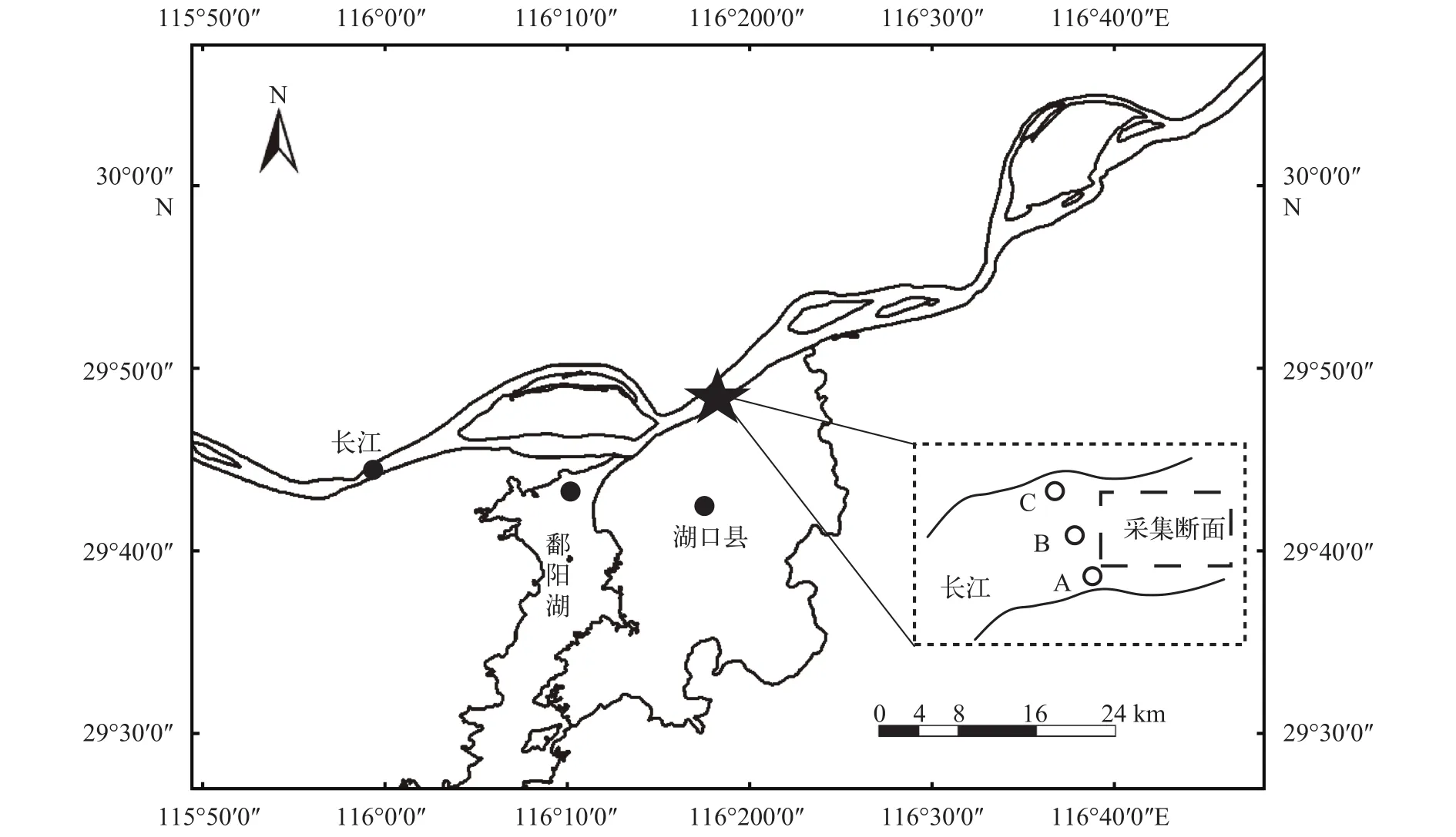
图 1 湖口江段鱼类早期资源采样点设置Fig. 1 The location of sampling sites in Hukou section on fisheries resources of early life history stages
使用定置网具采集的冰水样品转酒精后作为分子鉴定的样本, 具体步骤是: 形态学与分子生物学相结合, 5%的福尔马林溶液保存的样品中难以鉴定的物种在冰水保存的样品中利用形态学方法进行对比, 找出形态相同的样本, 冰水转至75%的中性酒精保存后进行分子鉴定。分子生物学的鉴定方法即提取样本的基因组DNA, 基于COⅠ (Cytochrome C oxidase Subunit Ⅰ, 线粒体细胞色素C氧化酶亚基Ⅰ)基因条形码, 采用PCR扩增目的条带并测序, 将所测结果同已经用成鱼建立的长江鱼类条形码数据库进行比对得到种类鉴定的结果[26]。
1.3 水文资料收集和数据处理
从长江水文网(www.cjh.com.cn)获得湖口江段的水位信息。仔稚鱼密度计算方法参考易伯鲁等[7]和曹文宣等[1]的计算方法, 依据所采集仔稚鱼的数量、采集时间、流速仪流量差和主动网具网口面积按下列公式处理:
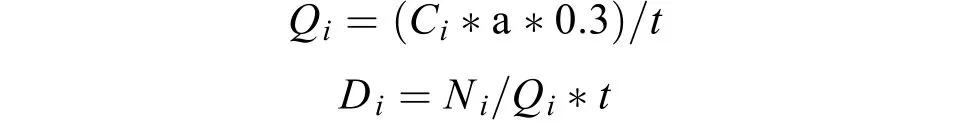
式中,Qi为第i次采集主动网具网口的过水流量(m3/s),Ci为第i次采集流速仪的流量差,a为主动网具网口面积(m2), 0.3为流速仪的常数,t为每次采集仔稚鱼的时间(s),Di为第i次采集仔稚鱼的密度(ind./m3),Ni为第i次采集仔稚鱼数量(ind.)。
依据主动网具采集的仔稚鱼的数量和出现的频率计算各种仔稚鱼的相对重要性指数(Index of Relative Importance,IRI):
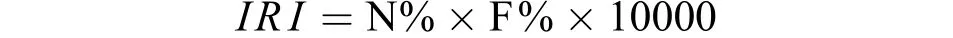
式中N%代表某种仔稚鱼数量占其群聚总数量的比例, F%代表采样期间某种仔稚鱼出现的频率, 依据IRI将仔稚鱼分成: 优势种(IRI≥100), 常见种(IRI≥10), 少见种(IRI<10)[27]。
1.4 统计分析
使用PRIMER 5.0软件分析仔稚鱼群落的时间动态。将调查期间不同时期作为样本, 仔稚鱼种类为变量, 建立1个仔稚鱼密度的原始矩阵, 再根据Bray-Curtis相似性系数, 构建仔稚鱼群落的相似性三角矩阵。根据不同时期的相似性指数, 使用组平均连接法进行等级聚类分析(Cluster analysis)和非参数多维尺度分析(non-Metric Multidimensional scaling, MDS), 并通过ANOSIM对不同群落矩阵间差异显著性进行检验[28—31]。检验 MDS分析结果的优劣用胁强系数(stress)来衡量, 通常认为当 stress<0.2时, 其图形有一定的解释意义; 当stress<0.1时,可以认为是一个好的排序; 当stress<0.05时, 说明具有很好的代表性[32]。
用Canoco 4.5软件对仔稚鱼密度和水环境因子的相关关系进行冗余分析, 按照仔稚鱼物种在各采样点出现的频度>12.5%, 且至少在1个采样点的相对密度≥1%的条件下进行物种筛选[33]。由于原始数据较为离散, 对仔稚鱼密度及水环境因子均进行log (X+1)处理, 以满足所分析数据呈正态分布[34]。数据分析使用SPSS 19.0软件处理, 相关性分析采用Pearson相关性检验。
2 结果
2.1 种类分析
种类组成对湖口江段采集的仔稚鱼分析其种类组成, 共鉴定隶属于8目12科39个仔稚鱼类群, 其中37个类群鉴定到种, 1个类群鉴定到亚科,1个类群鉴定到科。在鉴定到种的37个类群中, 包括鲤科19种, 鳅科4种, 鮨科、塘鱧鱼科、鲿科、银鱼科各两种, 虾虎鱼科、鱵科、鳀科、斗鱼科、月鱧科以及青鳉科各一种(表 1)。
仔稚鱼优势种和常见种对定量采集的仔稚鱼进行IRI分析发现优势种有4种, 常见种6种, 其余的都为少见种(表 2)。按相对重要性指数高低,优势种依次为贝氏䱗(Hemiculter bleekeri) (6831.33)、虾虎鱼科(Gobiidae) (1583.12)、鳊(Parabramis pekinensis) (410.49)、飘鱼(Pseudolaubuca sinensis) (287.73),常见种依次为银鲴(Xenocypris argentea) (96.60)、刀鲚(Coilia nasus) (92.30)、翘嘴鲌(Culter alburnus)(62.22)、间下鱵(Hyporhamphus intermedius) (27.34)、四大家鱼(20.32)、鳜(Siniperca chuatsi) (13.41)。其中贝氏餐出现频率和第二优势种虾虎鱼科物种持平, 但其仔稚鱼数量占群聚总数量的比例高于虾虎鱼科, 约占群聚总数的68.31%, 四种优势种一共占群聚总数量的97.3% (图 2)。
2.2 仔稚鱼群落结构的时空分布
仔稚鱼种类数的时间变化调查期间仔稚鱼种类数最高在7月4日达到16种, 7月4日之前仔稚鱼种类数呈上升趋势, 随后又回到之前水平(图 3)。
仔稚鱼密度时间变化及断面空间对比采样调查期间, 湖口江段仔稚鱼每日的平均密度呈波段性变化, 从5月18日至7月1日仔稚鱼平均密度整体趋势为在波动中逐渐上升, 在7月1日达到整个调查期间仔稚鱼平均密度的峰值, 峰值为1230.77 ind./100 m3; 7月1日后直至8月15日仔稚鱼平均密度整体呈波动性下降的趋势。在仔稚鱼平均密度上升的过程中除去最高峰一共出现5次小峰值, 日期分别是6月1日、10日、17日、25日和28日, 其仔稚鱼平均密度分别为797.95、775.86、809.07、1080.68和1230.77 ind./100 m3, 6月25日至7月1日湖口江段仔稚鱼平均密度在较高的水平上持续了1周; 在下降过程中出现2个小峰值, 日期为7月6日和14日, 仔稚鱼平均密度分别为475.17和400.91 ind./100 m3(图 4)。由图 4可见, 断面上3个采样点仔稚鱼密度变化趋势较为一致, 其中南北两岸采样点仔稚鱼的密度高于断面中心的采样点, 南岸、北岸及中心采
样点最高仔稚鱼密度分别为3484.44、1342.26和391.09 ind./100 m3, 中心采样点仔稚鱼密度最高峰出现时间早于南北两岸。

表 1 2016年5—8月湖口江段鱼类仔稚鱼种类组成Tab. 1 The composition of larval and juvenile fish species in Hukou section from May to August in 2016
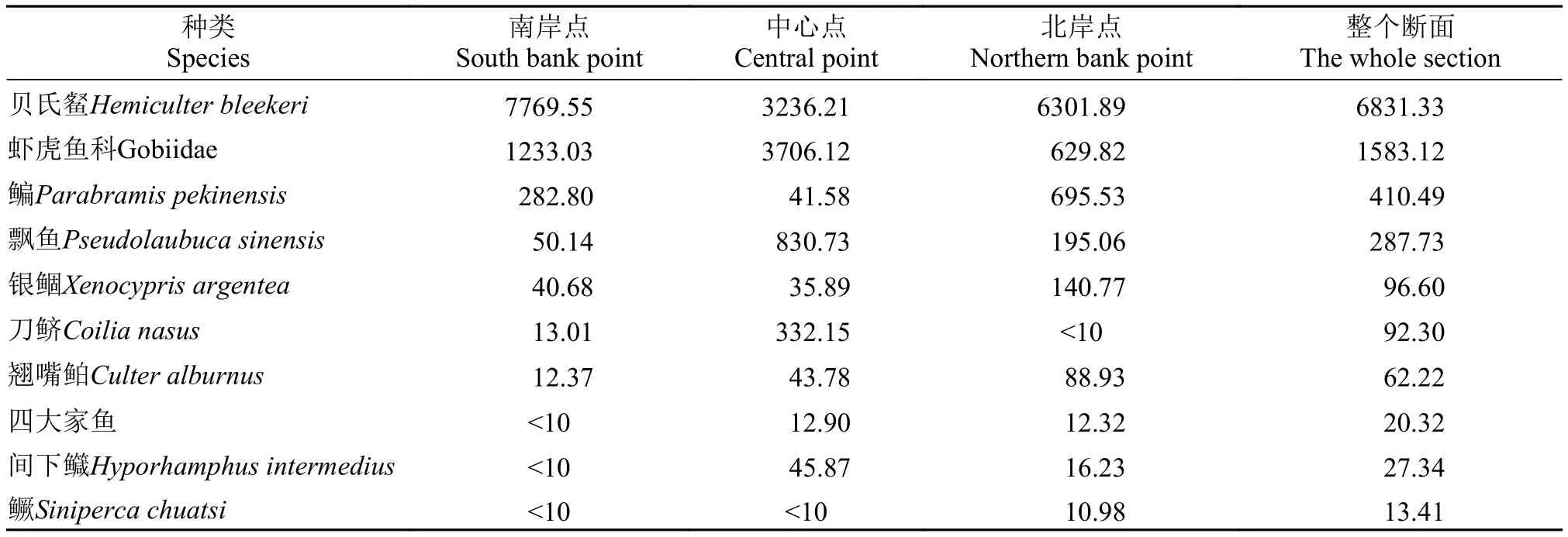
表 2 仔稚鱼的相对重要性指数Tab. 2 The index of relative importance of larval and juvenile fish
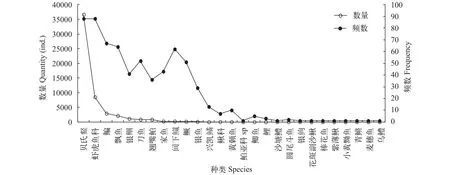
图 2 2016年5—8月湖口江段仔稚鱼群聚数量和出现频数排序Fig. 2 Rank abundance and occurrence of the larvae and juvenile species in Hukou section from May to August in 2016

图 3 2016年5—8月湖口江段仔稚鱼种类数量的逐日变化Fig. 3 The daily quantity of larval and juvenile fish species in Hukou section from May to August in 2016
仔稚鱼时间分布及聚类分析不同时期仔稚鱼种类密度显示, 各优势种与常见种在湖口江段出现时间不尽相同。如图 5所示, 贝氏䱗出现在5月下旬至8月上旬, 6月下旬达到峰值(39.87 ind./100 m3);虾虎鱼科物种出现在5月下旬至8月上旬, 密度持续下降, 5月下旬达到峰值(7.55 ind./100 m3); 刀鲚出现在6月上旬至8月上旬, 6月下旬达到峰值(0.78 ind./100 m3); 飘鱼5月下旬至8月上旬, 8月上旬达到峰值(1.28 ind./100 m3); 银鲴出现在5月下旬至7月上旬,密度持续下降, 5月下旬达到峰值(1.32 ind./100 m3);鳊出现在6月上旬至8月上旬, 7月上旬达到峰值(3.50 ind./100 m3); 翘嘴鲌出现在6月下旬至8月上旬, 6月下旬达到峰值(1.82 ind./100 m3); 四大家鱼出现在6月上旬至8月上旬, 7月上旬达到峰值(0.35 ind./100 m3); 间下鱵出现在5月下旬至7月下旬, 6月上旬达到峰值(0.14 ind./100 m3); 鳜出现在5月下旬至8月上旬, 7月上旬达到峰值(0.15 ind./100 m3)。
由各时期聚类分析树状图可以看出(图 6), 以相似度65为界限可将6个时期分为两组: 5月下旬至7月上旬聚为一组, 7月下旬至8月上旬聚为一组。其中, 5月下旬至7月上旬四个时期以相似度70为界可分两小组: 5月下旬至6月上旬相似度较高, 聚为1组; 6月下旬至7月上旬相似度较高, 聚为一组。
基于各时期间 Bray-Curtis 相似性指数的6个时期仔稚鱼群落MDS平面图的胁强系数为0.01 (<0.05), 能很好地解释各时期间的聚类结果, MDS图具有很好的代表性。相似性分析(Analysis of similarity, ANOSIM)对组间差异显著性进行检验, 结果显示R=0.607,P(level%)=6.7%, 说明两组间差异性不显著(图 7)。
2.3 仔稚鱼与水环境因子的关系
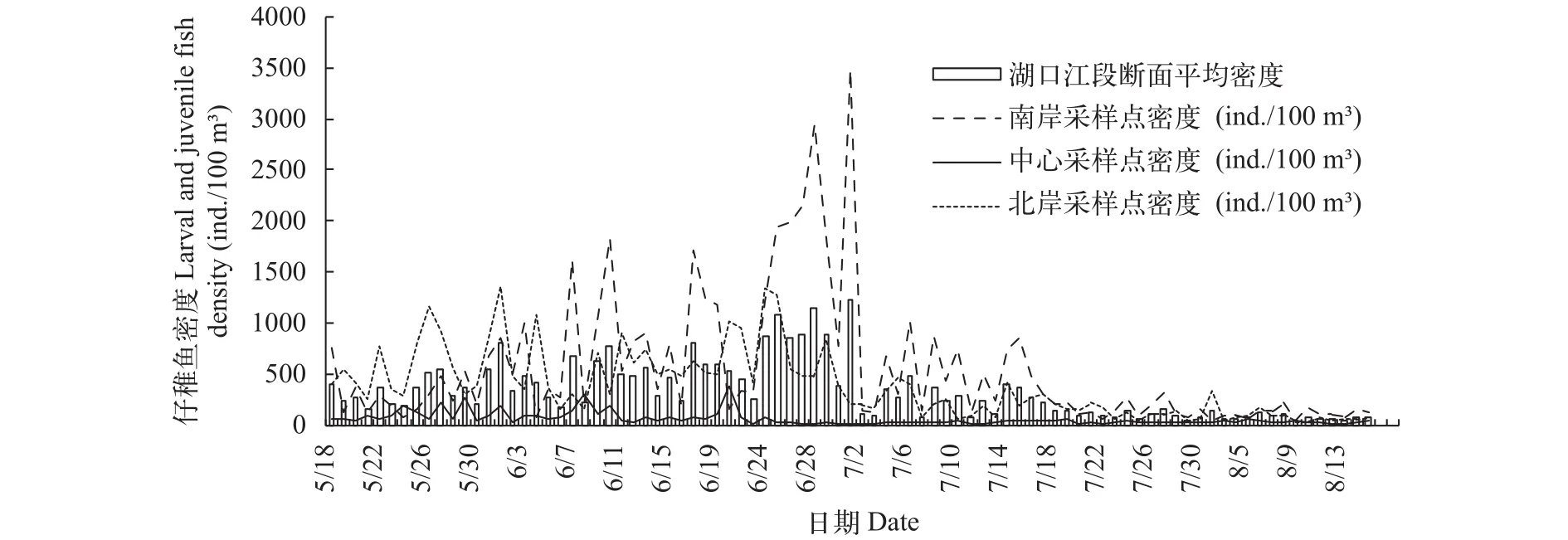
图 4 2016年5—8月湖口江段仔稚鱼密度变化与断面空间对比Fig. 4 The comparison of average density and sectional space of larval and juvenile fish in Hukou section from May to August in 2016

图 5 仔稚鱼优势种与常见种在各时期出现情况Fig. 5 The dominant and common species of larval and juvenile fish at various times
仔稚鱼密度变化与环境因子的关系湖口江段的水位在5月18日至6月28日一直保持在相对平稳的状态, 在此期间最高水位仅达18.75 m, 最低水位为17.34 m, 上下波动不超过1.5 m, 此时仔稚鱼密度逐渐增加; 6月29日起, 湖口江段水位逐渐上涨, 仔稚鱼密度在水位高涨前达到峰值; 水位在7月9日涨至最高点, 持续3d后逐渐下降, 在水位涨至最高点前后仔稚鱼密度出现了3个小峰值; 调查末期水位才恢复到之前正常水平, 此时仔稚鱼密度也下降到较低水平。对水位与仔稚鱼密度的相关性分析表明, 两者呈显著相关(图 8a,P<0.01)。调查期间平均水温为25.8℃, 水温变化范围为21.5—30.2℃, 变化幅度达到8.7℃(图 8b); 平均透明度为42.03 cm, 变化范围为22.6—55.7 cm, 仔稚鱼密度达到峰值时, 透明度为45 cm (图 8c); 平均水流量为0.44 m3/s, 变化范围为0.28—0.56 m3/s (图 8d)。
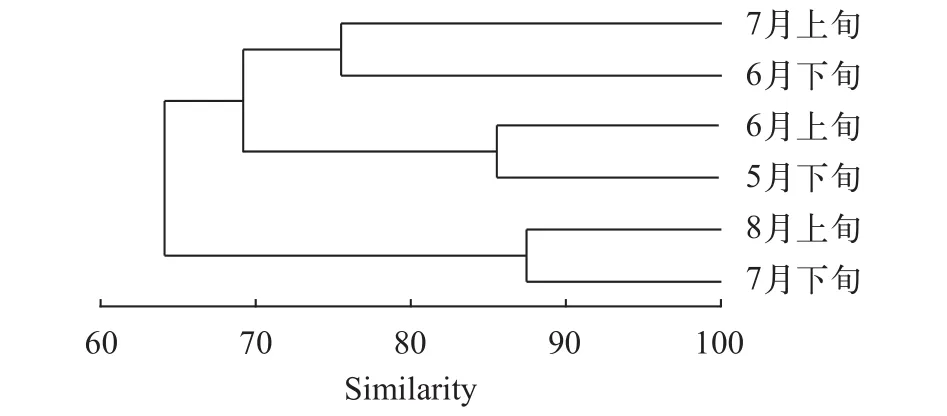
图 6 湖口江段各时期仔稚鱼群落聚类分析图Fig. 6 Cluster dendrogram of larval and juvenile fish community from 6 periods in Hukou section
仔稚鱼与水环境因子相关关系的冗余分析
对经过筛选的10种仔稚鱼及3种环境因子进行DCA分析。分析结果表明, 湖口江段的第一轴梯度长度(Lengths of gradient)小于3.0, 适合进行RDA分析。由表 3可知, 3种环境因子共能反应5.7%的物种信息, 轴1和轴2的特征值分别为0.055和0.002; 环境因子轴与物种排序轴的相关系数为0.258和0.149; 轴1和轴2解释了5.7%的物种信息和99.5%物种-环境关系信息。经蒙特卡洛(Monte Carlo) 检验,水温、水流量和透明度3种环境因子对仔稚鱼群聚均有显著的影响(P<0.05)。
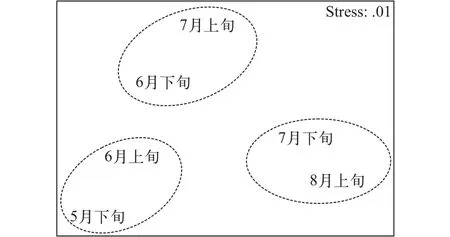
图 7 湖口江段各时期仔稚鱼群落度排序分析图Fig. 7 MDS ordination of larval and juvenile fish community from 6 periods in Hukou section
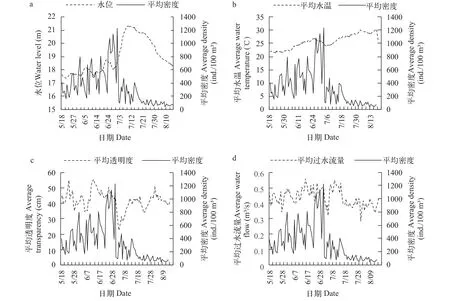
图 8 2016年湖口江段仔稚鱼密度时间变化与环境因子的关系Fig. 8 The relationship between the average density of larval and juvenile fish and environmental factors in Hukou section in 2016
RDA排序图反应出10种仔稚鱼对各环境条件的不同适应特点(图 9)。在由轴1和轴2构成的平面上, 与轴1相关性较高的是水温(T), 与轴2相关性较高的是水流量(FLow)和透明度(SD)。物种与环境因子的关系显示, 贝氏䱗(SP1)、间下鱵(SP9)与水流量和透明度呈正相关, 而与水温呈负相关; 虾虎鱼科(SP2)、银鲴(SP5)与水温呈正相关; 鳊(SP3)、翘嘴鲌(SP7)、四大家鱼(SP8)、鳜(SP10)与水流量和透明度呈负相关; 飘鱼(SP4)、刀鲚(SP6)与水温、水流量和透明度均呈正相关。
3 讨论
3.1 湖口江段仔稚鱼物种组成
湖口江段仔稚鱼种类较为丰富, 本研究共鉴定了来自7目12科39个类群, 优势种有贝氏䱗、虾虎鱼科、鳊、飘鱼和银鲴, 它们在仔稚鱼密度上较其他鱼类有更大的占有比例, 常见物种有银鲴、刀鲚、翘嘴鲌、四大家鱼、间下鱵和鳜等。陈文静等[19]在2007年至2009年对湖口江段的成鱼逐月进行了2—5次调查, 共监测到了隶属于6目7科的26个种类, 渔获物组成以黄颡鱼、粗唇 、刀鲚、鳊等为主, 其中长江定居型鱼类占比最高, 其次是江湖洄游性及河海洄游性鱼类。本次调查监测的仔稚鱼物种组成较往年成鱼相比多13种, 群落组成结构也与成鱼较为不同, 仔稚鱼群落中江湖洄游性鱼类占比较高, 但仔稚鱼和成鱼的群落组成均以中小型鱼类为主, 这可能由鱼类的繁殖策略决定的[35,36]。成鱼具备良好的主动游泳能力, 可自主选择栖息地,而鱼类仔稚鱼时期处于被动漂流阶段, 鱼苗依靠自身重力和水流产生的上举分力处于动态平衡中, 其群落组成受河流形态、流量的影响, 从而显示出与成鱼群落不同的群落特征, 不同河流和江段中的仔稚鱼群落特征差别则更为明显[12,37,38]。
3.2 仔稚鱼的群落结构
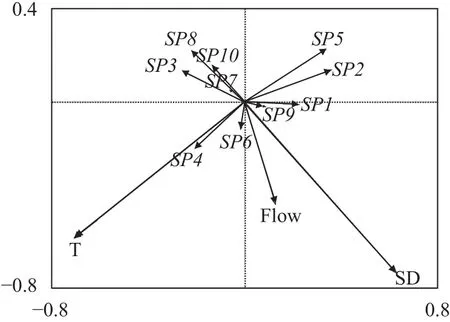
图 9 湖口江段鱼类仔稚鱼物种-环境关系的RDA二维排序图Fig. 9 RDA biplot of larval and juvenile fish species and environmental variables in Hukou section
鱼类早期生活史的时间动态变化直接反映了其具体的繁殖时间[39], 湖口江段仔稚鱼密度变化和时间尺度上的分析表明, 仔稚鱼主要集中在5月下旬至7月上旬大量出现, 6月下旬总密度达到峰值,这可能是由于此时段是长江下游鱼类繁殖盛期所致[40]。在时间尺度上影响仔稚鱼群落结构有鱼类生活史策略(内源)和外界环境因子(外源)两大因素。内源因素方面如贝氏䱗在整个调查期间都占据着绝对优势种地位, 其繁殖季节长、性成熟早,具有快速的世代更替率, 是典型的机会策略物种,用连续产卵的模式保证对其种群的补充, 使其在繁殖期内仔稚鱼阶段一直占据着优势种群地位; 优势种中飘鱼也具有同样的生活史策略[35,41,42]。王红丽等[43]对三峡库区江段早期资源的研究中也调查到6月份翘嘴鲌与鳜等种类开始增加, 调查结果与本研究一致, 其中鳜在七月上旬达到峰值。鳜属肉食性凶猛鱼类, 其仔鱼阶段就开始以其他种类仔鱼为食, 有研究指出, 丰富的饵料会促进鳜鱼的繁殖,湖口江段仔稚鱼密度增加对鳜种群的增长也起到一定促进作用[44]。鄱阳湖属于长江泛滥平原湖泊之一, 湖口江段与其相通, 有研究表明泛滥平原的连通度是影响其物种多样性的一个重要因素, 江湖洄游型鱼类随长江水位上涨而进入长江进行繁殖活动, 胡茂林等在2007—2008年对长江四大家鱼幼鱼入湖格局的研究中发现湖口江段水位与四大家鱼幼鱼入湖数量呈正相关关系, 结合此次调查随水位和水流量的升高, 仔稚鱼的种类数量和密度都随之达到峰值, 说明水位的变化对湖口江段仔稚鱼群落多样性有着一定影响, 这也在一定程度上反映了湖口江段江湖连通的重要性[18,45—47]。
长期以来, 鄱阳湖与长江的交汇处由于鄱阳湖自西而来的清澈湖水与长江水汇合, 形成了一条清浊分明的水界限[48]。江段中心可能由于江湖水的汇合产生巨大的冲击, 中心地带的流速加快, 泥沙增多, 使得其仔稚鱼密度远低于两岸的密度, 造成空间上的差异[49]。由于江湖交汇处具有独特的空间异质性, 需有针对性的对不同鱼类的产卵场、育幼场以及影响鱼类繁殖的关键环境因子制定出不同的保护管理政策, 使其能有效为补充群体做出贡献。因此为了对长江下游以及鄱阳湖的鱼类多样性进行保护, 建议将湖口江段这一块连接长江与鄱阳湖的关键通道作为重点水生生物资源的保护区域。
3.3 环境因子与湖口江段仔稚鱼密度的关系
RDA分析结果表明, 水温、流量和透明度均为影响仔稚鱼群落结构的重要环境因子。仔稚鱼的密度变化及群落组成不是受单个环境因素的影响,而是由各种环境因子共同作用, 其主要机制是外界环境因子通过刺激亲鱼内部生理机制使之变化产生一系列产卵繁殖动作造成的[50]。研究表明, 水温是影响鱼类生理、生化及其生活史的主要环境因子, 本次调查结果发现湖口江段各种鱼类产卵最适合温度在21.5—27℃, 这与曹文宣等[1]对长江下游各种鱼产卵的温度调查结果基本一致[51]。四大家鱼、翘嘴鲌、鳜等与透明度有着明显的负相关关系, 但仔稚鱼总密度随着透明度的下降有一个急剧下滑的过程。透明度是反应水体污染程度大小的一个综合表征, 过高或过低的透明度均会对鱼类摄食及繁育产生一定的影响[52]。邬国锋等[48]在2005年和2006年借助遥感技术发现长江与鄱阳湖交汇处出现长江水清而鄱阳湖水浑浊的清浊倒置现象,这种现象导致的透明度变化, 可能对鱼类的产卵活动产生极大的影响。刀鲚等与水温、透明度和流量均呈正相关关系, 徐钢春等[53]对刀鲚人工繁殖的研究过程中也显示, 使用水流刺激是池塘内成功培育性腺成熟刀鲚的关键因素, 说明刀鲚的繁殖对环境的要求较高, 水流量对其生殖过程至关重要。除去非生物因素, 生物因素也对仔稚鱼群落结构影响重大, 如浮游动植物密度大小和群落结构对仔稚鱼的生长与存活有着直接影响, 鉴于此, 后续将加入生物因素来深入研究其变化与湖口江段仔稚鱼群落结构的关系[54]。
3.4 对长江湖口段渔业资源保护的建议
与湖泊直接相连的江段是形成江湖复合生态系统不可或缺的环境单元结构[55], 长江湖口段鱼类群落结构的变动对鄱阳湖及长江中下游的渔业资源状况都有广泛影响。由于此次调查开展在禁捕期内, 本研究没有结合当时的成鱼资源状况进行系统分析, 后期研究将加入对湖口江段成鱼资源的调查, 完善湖口江段整体的鱼类群落结构, 分析仔稚鱼资源变动对成鱼资源状况的影响。为了更有效的发挥江湖复合生态系统的作用, 利用江湖复合生态系统来恢复和保护长江中下游渔业资源, 基于以前学者及本文研究结果建议: (1)在江湖连通江段全面禁渔。只有在湖口江段实现全面禁渔才能保证江湖连通的自然性, 使长江中下游鱼类种群顺利进入鄱阳湖生存繁殖。(2)基于成鱼调查数据, 结合历史资料制定湖口江段增殖放流方案, 为长江下游补充鱼类繁殖群体, 逐渐恢复原有鱼类群落结构。(3)逐步撤离江段周边具有污染性质的工厂。鱼类仔稚鱼的活动能力不强, 沿岸的工厂排放至水中的污染造成仔稚鱼个体的死亡, 缩小了其生存空间,直接影响了仔稚鱼的群落结构。
猜你喜欢
水产科学(2022年1期)2022-01-26
渔业科学进展(2021年6期)2022-01-04
浙江海洋大学学报(自然科学版)(2021年1期)2021-07-06
中国水利(2020年23期)2020-12-24
北方音乐(2020年17期)2020-12-06
科学导报(2020年27期)2020-05-13
浙江林业(2016年5期)2016-06-14
资源节约与环保(2016年12期)2016-02-15
应用海洋学学报(2015年3期)2015-11-22
