
转Cry1Ac/1Ab基因棉花对异色瓢虫生长发育及其捕食功能的影响
2019-04-13 01:56朱香镇雒珺瑜李笠坤吴益东李春花崔金杰
生物安全学报 2019年1期
朱香镇, 雒珺瑜, 李笠坤,, 吴益东, 李春花, 崔金杰*
1中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南 安阳 455000;2南京农业大学植物保护学院/昆虫分子毒力学实验室,江苏 南京 210095
近20年来,随着转基因棉花在我国大规模的商业化种植,其转基因技术得到迅猛发展,越来越多的新型基因被导入棉花中,转基因棉花的种类也蜂拥出现(刘晨曦和吴孔明,2011),其中复合性状转基因棉花、转特殊基因棉花、转融合基因棉花等新型转基因棉花已成为商业化开发的一个重要趋势。新型转基因棉花和传统转单价基因棉花一样,大规模的种植可能会对生态环境产生潜在的影响,因此,在商业化种植前对新型转基因棉花的环境安全性评价显得尤为重要。
环境安全性评价主要包括靶标昆虫对转基因棉花产生的抗性、转基因作物的杂草化趋势、基因漂流、对有益昆虫和天敌生物以及生物多样性的影响等问题(钱迎倩等,2001; 吴孔明,2007; Hails,2000)。异色瓢虫Harmoniaaxyridis(Pallas)作为一种捕食性很强的天敌生物广泛存在于农田生态系统中(王甦等,2007),其捕食对象包括各种蚜虫、叶螨和介壳虫等同翅目昆虫以及鞘翅目、膜翅目、双翅目和鳞翅目昆虫的幼虫和蛹(Dixon,2000; Obrycki & Kring,1998)。捕食性天敌在维持田间生态平衡和复杂的生态链锁方面起着重要作用;而当转基因作物种植后,植物体内或组织上表达出异源蛋白或毒蛋白如花粉、花蜜、叶片等,被害虫取食后,影响害虫的生长发育和繁殖等,最终导致天敌生物猎物质量与数量改变(Harwoodetal.,2005; Zwahlen & Andow,2005)。通过三级营养关系评估转基因作物对天敌生物生长发育影响以及对农田害虫的捕食和控制作用已在转基因水稻、玉米、棉花等作物上成为研究热点(lvarez-Alfagemeetal.,2009; Hanetal.,2014; Romeisetal.,2004)。
丁瑞丰等(2016)以转Bt+CpTI基因抗虫棉花及其亲本为材料对棉花—棉蚜AphisgossypiiGlover —普通草蛉Chrysoperlacarnea(Stephens)三级营养关系进行研究,结果表明转基因棉花对普通草蛉捕食功能反应和搜寻效应没有显著的影响;施敏娟(2012)研究发现转Bt基因抗虫棉对中华草蛉ChrysoperlasinicaTjeder的生长发育和繁殖无不良影响,并且ELISA检验的结果显示,中华草蛉体内不含Bt毒蛋白,说明Bt毒蛋白不会经过棉蚜传至其天敌中华草蛉体内;对捕食性天敌草间小黑蛛Erigonidiumgraminicolum利用棉花—棉铃虫—草间小黑蛛三级营养关系的研究表明,转Cry1Ac+Cry2Ab基因棉花对草间小黑蛛体重及其体内外源蛋白含量的影响均不显著,并且对草间小黑蛛体内相关解毒酶和保护酶活性亦无显著影响(雒珺瑜等,2013);还有研究转Bt基因棉花对烟粉虱Bemisiatabaci(Gennadius)天敌昆虫龟纹瓢虫Propyleajaponica(Thunberg)影响时发现,龟纹瓢虫对烟粉虱的捕食功能符合HollingⅡ型反应,并且转Bt基因棉花上的烟粉虱对龟纹瓢虫的发育历期和存活率没有明显的影响(周福才等,2013)。
本研究以转Cry1Ac/1Ab基因棉花为实验对象,以其常规亲本棉为对照,研究饲喂不同棉花的初孵棉铃虫对天敌生物异色瓢虫生长发育的影响,以及不同龄期异色瓢虫对初孵棉铃虫的捕食功能,旨在探索我国转新型基因棉花的环境安全评价理论与方法,同时为我国转基因棉花对天敌生物安全性评价提供更多的参考依据。
1 材料与方法
1.1 供试棉花
转Cry1Ac/1Ab基因棉花品系(A26-5)和其亲本常规棉花品系(J14)由创世纪种业公司提供,种植在中国农业科学院棉花研究所东场实验田内,供试棉花叶片为直接在实验田块A26-5和J14采集的倒三叶。整个实验过程中田间未使用任何化学农药。
1.2 供试昆虫及饲养方法
异色瓢虫采集于非转基因棉田,在室内经人工饲养至产卵后,将瓢虫卵放在透明并带有透气孔的塑料盒内(直径9 cm,高10 cm),置于光照培养箱[温度(26±1) ℃,湿度 60%~70%,L/D=14/10]待孵化;棉铃虫由中国农业科学院棉花研究所提供,在室内利用人工饲料饲养多代且未使用任何农药。
1.3 饲喂转基因棉花的棉铃虫对异色瓢虫生长发育及体重的影响
在第二代棉铃虫发生期(棉花生长的蕾期),采集不同棉花品种的上部倒三叶,饲喂初孵棉铃虫幼虫后备用。选择体型大小一致的棉铃虫幼虫用于异色瓢虫捕食功能实验,转基因棉花对棉铃虫幼虫的生长发育具有抑制作用,因此,幼虫饲喂转基因棉花24 h,饲喂对照叶片12 h。初孵异色瓢虫幼虫每20头为一组,瓢虫幼虫在20 mm × 100 mm小试管中单头饲养,重复3次。瓢虫用初孵棉铃虫饲喂,每天更换棉铃虫初孵幼虫;小试管内事先放入清洁纸条和保湿的棉球,每天更换一次。观察其生长发育,记录蜕皮情况,用1/100000的天平对蜕皮后的瓢虫称重,直到所有供试瓢虫产卵结束为止。
1.4 饲喂转基因棉花的棉铃虫对异色瓢虫捕食功能反应的影响
异色瓢虫的捕食功能反应以取食不同转基因棉花的棉铃虫初孵幼虫为猎物,并在室内统一饥饿12 h后进行实验;在5个塑料盒中分别放置5、10、15、20、25头棉铃虫初孵幼虫,然后分别放入1头异色瓢虫1龄幼虫,每个密度设置5个塑料盒为1次重复,实验共重复3次;透明塑料盒的直径为9 cm,高为10 cm,置于光照培养箱中培养。对于异色瓢虫2、3和4龄幼虫的捕食功能反应模型,实验步骤同异色瓢虫1龄幼虫,只是设置的初孵棉铃虫的密度不同。对于异色瓢虫2龄幼虫,设置的5个棉铃虫初孵幼虫密度分别为10、20、30、40和50头;对于异色瓢虫3龄幼虫,设置的5个棉铃虫初孵幼虫密度分别为15、30、45、60和75头;对于异色瓢虫4龄幼虫,设置的5个棉铃虫初孵幼虫密度分别为30、45、60、75和90头。24 h后记录取食棉铃虫幼虫个数,并建立捕食功能反应模型——HollingⅡ型反应模型。功能反应圆盘方程为:1/Na=1/(aT)×1/N+Th/T。式中:Na为被捕食的猎物数量;N为猎物密度;a为瞬时攻击率;T为实验持续时间(1 d);Th为处置1头猎物时间。
1.5 数据分析方法
所有数据运用SPSS 12.0软件进行统计分析,采用单因素方差分析(One-way analysis,ANOVA)和Duncan′s 差异显著性分析。
2 结果与分析
2.1 转基因棉花对异色瓢虫生长发育及体重的影响
2.1.1 转基因棉花对异色瓢虫幼虫发育历期的影响 如表1所示,与常规亲本棉相比,取食饲喂转基因棉花叶片的初孵棉铃虫幼虫的异色瓢虫幼虫发育至2龄时间缩短0.33 d,差异不显著(P>0.05),但发育至3、4龄、蛹期时间均被延长,分别延长0.30、0.57、0.23 d,差异均不显著(P>0.05);异色瓢虫幼虫从1龄发育至化蛹期时间延长0.77 d,但差异不显著(P>0.05)。
2.1.2 转基因棉花对异色瓢虫幼虫体重的影响 如表2所示,与常规亲本棉相比,取食饲喂转基因棉花叶片的初孵棉铃虫的异色瓢虫幼虫,除1龄幼虫体重增加0.0773 mg外,其余各龄期幼虫体重均有所下降,但差异均不显著(P>0.05)。

表1 不同棉花品种对异色瓢虫幼虫发育历期的影响Table 1 Response of developmental duration of H.axyridis larvae to the varieties of cotton

表2 不同棉花品种对不同龄期异色瓢虫幼虫体重的影响Table 1 Body weight of H.axyridis larvae fed with the two cotton varieties
2.2 转基因棉花对异色瓢虫捕食功能反应的影响
异色瓢虫1、2、3、4龄幼虫对初孵棉铃虫捕食量均随棉铃虫密度的增加而增加,当棉铃虫密度达到一定限度后不再增加(图1)。各龄期异色瓢虫幼虫对饲喂不同棉花品种的初孵棉铃虫捕食功能均符合HollingⅡ型反应。通过捕食功能反应模型拟合实验数据(表3),不同龄期的异色瓢虫幼虫对初孵棉铃虫的功能反应相关系数均高于0.9284,达显著相关水平,说明实验得到的数据模型能反应不同龄期异色瓢虫在不同棉铃虫密度下的捕食规律。随着异色瓢虫龄期的增长,其对饲喂2种棉花的棉铃虫瞬间攻击率加快,即异色瓢虫1龄时对饲喂A26-5和J14棉花的棉铃虫攻击率分别为0.8818和0.8173,而至4龄时瞬间攻击率达到0.9521和0.9992;随着异色瓢虫龄期的增长,其对棉铃虫的处理时间逐渐缩短,即异色瓢虫取食饲喂A26-5和J14棉花的棉铃虫的处理时间从1龄的0.0161和0.0172 h减少到4龄的0.0026和0.0043 h,但异色瓢虫对饲喂2种棉花的棉铃虫的瞬间攻击率和处理时间差异均不显著(P>0.05)。
3 讨论
转基因作物的出现,一方面使农业生产成本大幅度降低并且有效地提升了品质和产量,另一方面却给生态环境安全带来不确定性和不稳定性。研究表明,转基因作物对生态环境安全的影响最为重要的是其对非靶标生物的影响(Conneretal.,2003; Paul,2002)。实验结果表明,与常规亲本棉相比,转Cry1Ac/1Ab基因棉对异色瓢虫生长发育历期无显著影响,取食饲喂转基因棉花叶片的初孵棉铃虫的异色瓢虫幼虫,除1龄幼虫体重增加外,其余各龄期幼虫体重均有所下降,但差异均不显著,与雒珺瑜等(2013)、Hanetal.(2014)、Zhuetal.(2006)的研究结果一致。本实验是捕食性天敌通过三级营养间接取食转基因作物,因此直接或间接取食转基因作物是否增加或减轻捕食性天敌的体重还需进一步研究(Sayyedetal.,2003)。
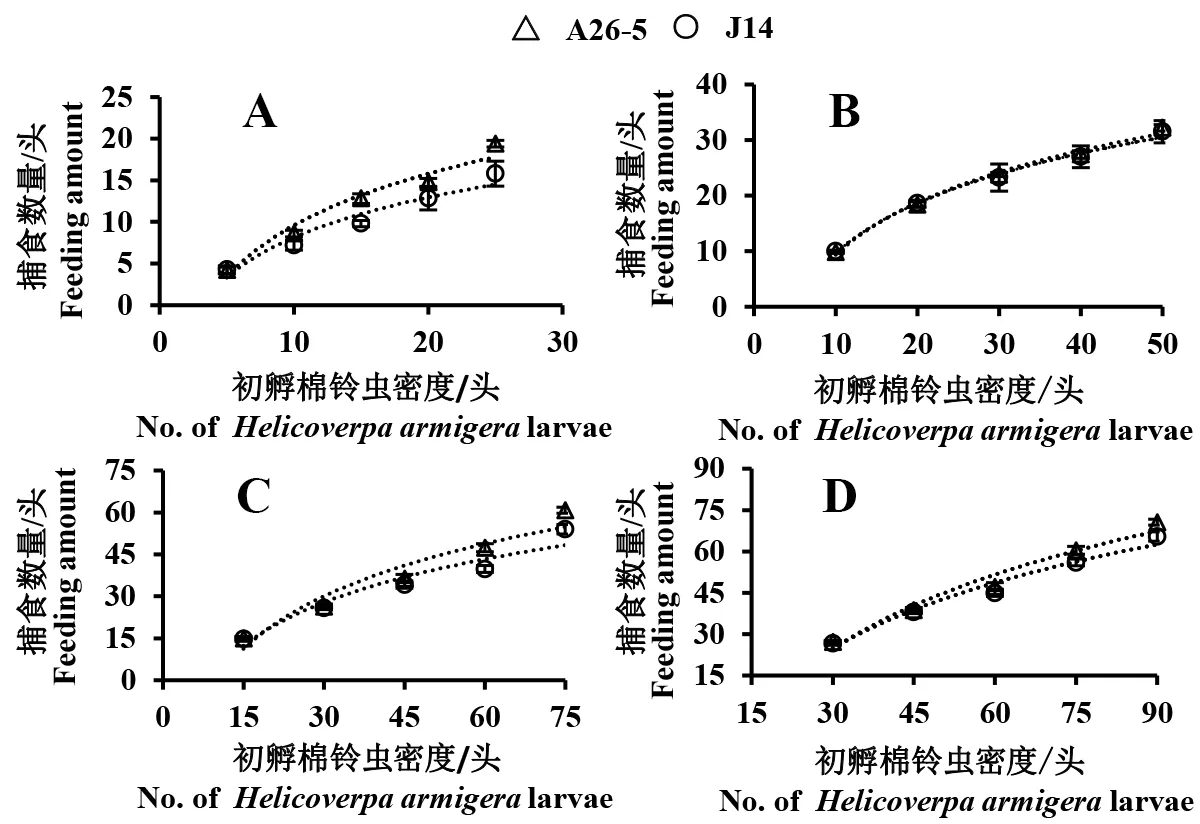
图1 不同龄期异色瓢虫对饲喂不同棉花初孵棉铃虫的捕食功能反应曲线Fig.1 The response curves of H.axyridis larvae preying on bollworms feeding in different cotton varieties A:异色瓢虫1龄幼虫;B:异色瓢虫2龄幼虫;C:异色瓢虫3龄幼虫;D:异色瓢虫4龄幼虫。 A: 1st instar H.axyridis larva; B: 2nd instar H.axyridis larva; C: 3rd instar H.axyridis larva; D: 4th instar H.axyridis larva.
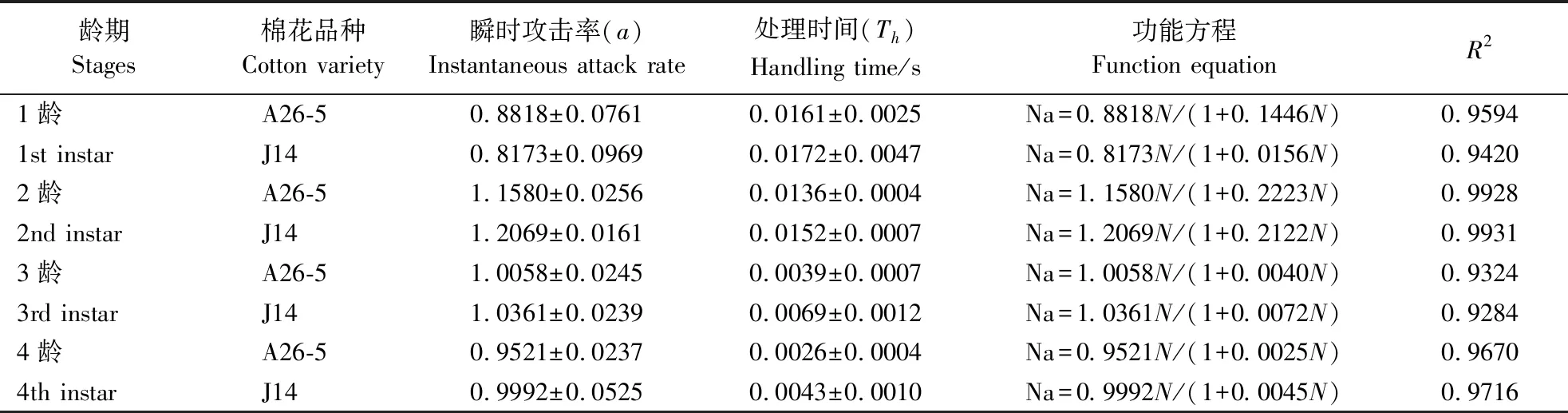
龄期Stages棉花品种Cotton variety瞬时攻击率(a)Instantaneous attack rate处理时间(Th)Handling time/s功能方程Function equationR21龄A26-50.8818±0.07610.0161±0.0025Na=0.8818N/(1+0.1446N)0.95941st instarJ140.8173±0.09690.0172±0.0047Na=0.8173N/(1+0.0156N)0.94202龄 A26-51.1580±0.02560.0136±0.0004Na=1.1580N/(1+0.2223N)0.99282nd instar J141.2069±0.01610.0152±0.0007Na=1.2069N/(1+0.2122N)0.99313龄 A26-51.0058±0.02450.0039±0.0007Na=1.0058N/(1+0.0040N)0.93243rd instar J141.0361±0.02390.0069±0.0012Na=1.0361N/(1+0.0072N)0.92844龄 A26-50.9521±0.02370.0026±0.0004Na=0.9521N/(1+0.0025N)0.96704th instar J140.9992±0.05250.0043±0.0010Na=0.9992N/(1+0.0045N)0.9716
摄取转基因产物的生物可能将未改变的转基因产物从植物传递给捕食者,而且生物能够通过自身的代谢潜在改变其生物化学活性并对捕食者产生某些不利的影响(Andow & Hilbeck,2004),其中包括对捕食性天敌捕食能力及功能反应的影响。功能反应(functional response)是捕食者对猎物的捕食效应,指环境中猎物密度的变化使猎物捕食率发生改变的一种反应,可用于鉴别捕食者对猎物的选择性以及生物防治中对敌害控制作用和转基因环境安全性评价的指标(Fernandez-arhex & Corley,2003; Murdochetal.,2003)。谢修庆等(2012)利用转Bt基因棉花、转Bt+CpTI基因棉花、常规棉为材料,研究了取食不同棉花品种的棉蚜对龟纹瓢虫1~4龄幼虫功能反应的影响,结果表明龟纹瓢虫对不同品种棉花上棉蚜的捕食量随猎物密度的增加而增大,均符合HollingⅡ圆盘方程,但对来自转基因棉田蚜虫的捕食量大多高于常规棉田,表明龟纹瓢虫对 Bt棉上的棉蚜并不存在排斥性。中华草蛉对取食转基因棉的甜菜夜蛾的捕食功能反应研究结果表明,与常规棉相比,中华草蛉对用转基因棉处理的甜菜夜蛾的捕食量和捕食效率均高于对照,但其捕食功能反应均符合HollingⅡ型(施敏娟等,2011)。
本研究表明,各龄期异色瓢虫幼虫对饲喂不同棉花初孵棉铃虫捕食功能反应均符合HollingⅡ圆盘方程,与常规棉相比,饲喂取食转Cry1Ac/1Ab基因棉花棉铃虫对异色瓢虫捕食功能无显著差异。但本研究是在室内条件下进行的,由于棉田自然环境因素复杂,害虫种类繁多,温度、湿度、光周期等多种因子在天敌捕食害虫过程起到某些干扰作用。因此,需进一步在棉田环境条件下研究异色瓢虫对棉铃虫的捕食能力;同时,转基因棉对其他天敌的影响还应进一步研究与完善。
猜你喜欢
上海蔬菜(2020年6期)2020-12-24
河北果树(2020年4期)2020-11-26
红蜻蜓·低年级(2020年9期)2020-09-26
农药科学与管理(2019年6期)2019-11-23
农民致富之友(2019年2期)2019-03-16
小学生作文(低年级适用)(2018年4期)2018-11-29
作物研究(2017年3期)2017-05-11
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
小朋友·快乐手工(2015年10期)2015-11-02
小学科学(2015年9期)2015-09-28
