
哮喘大鼠肺组织5-脂氧合酶mRNA,15-脂氧合酶mRNA表达失衡
2019-08-15 07:07陈泗虎革丽莎王小明李昌崇
医学研究杂志 2019年8期
陈泗虎 革丽莎 王小明 李昌崇
5-脂氧合酶(5-lipoxygenase,5-LO)和15-脂氧合酶(15-lipoxygenase,15-LO)是花生四烯酸代谢中关键的脂质过氧化物酶。一方面,5-LO催化花生四烯酸生成半胱氨酰白三烯,引起气道平滑肌收缩、气道黏液分泌增加、黏膜水肿、炎性细胞聚集及促进慢性哮喘患者的气道重塑[1]。另一方面,15-LO和5-LO通过跨细胞作用共同参与脂氧素(lipoxins,LXs,包括LXA4、LXB4及其衍生物)的合成,在体内起广泛的抗炎促消退作用[2,3]。目前认为,脂氧素的合成不足是哮喘气道炎症持续存在的重要原因[4,5];因而,推测5-LO,15-LO表达失衡是导致脂氧素合成障碍,以及哮喘不同阶段病理生理改变的重要因素。本研究通过观察5-LO,15-LO在幼年哮喘大鼠肺组织中的表达情况,以及与IgE、LXA4之间的关系,探讨它们在哮喘病理生理改变中的意义。
材料与方法
1.材料:(1)实验动物:健康SD(Sprague-Dawley)雄性大鼠,二级,4~6周龄,共21只,体重(105±20)g,温州医学院实验动物中心提供。(2)主要药品与试剂:鸡卵清白蛋白(OVA)、琼脂糖(美国Sigma公司),氢氧化铝[将NaOH和AlCl3按摩尔比3∶1混合,产生Al(OH)3沉淀,用三蒸水充分洗涤,适量生理盐水混合后高压灭菌],地塞米松磷酸钠注射液(浙江仙居制药1ml∶5mg,批号09072911)。大鼠LXA4ELISA试剂盒(上海西唐生物科技有限公司);大鼠卵白蛋白特异性IgE ELISA试剂盒(美国R&D公司),Trizol试剂(美国Invitrogen公司);反转录酶、PCR试剂(天根生化科技有限公司);PCR引物:由上海生工生物工程有限公司合成。(3)实验仪器:高速冷冻离心机 (德国 Hettich 公司),UV-2800型紫外可见分光光度计(美国UNICO公司),Bio-Tek 酶标仪(ELX808IU,美国),PCR仪(德国Biometra公司),凝胶成像系统(Biosens SC645 ,日本)。
2.动物模型制备与分组:随机将21只SD大鼠分为3组:正常对照组(n=7)、哮喘模型组(n=7)、地塞米松组(DXM组,n=7)。哮喘模型制备参照文献[6]:分别于第1天和第8天腹腔注射OVA/Al(OH)3混合液1.5ml[内含OVA 1mg,Al(OH)3100mg],第15天始连续3天定时定点给予1%OVA雾化吸入,每次30min。DXM组于每次雾化激发前1h,予地塞米松磷酸钠注射液0.5mg/kg腹腔注射。正常对照组和哮喘组均以等量0.9%氯化钠注射液替代。
3.标本留取:末次激发后12h内,10%水合氯醛4ml/kg腹腔注射麻醉。腹主动脉取血7ml,4℃凝固,2000r/min离心10min,取上层血清分装,-70℃保存,用于ELISA检测。取右肺中叶肺组织约50mg,液氮速冻后-70℃保存,行RT-PCR检测。
4.ELISA法检测血清LXA4、OVA特异性IgE含量:操作严格按试剂盒说明进行。最后使用Bio-Tek酶标仪测量450nm波长吸光度。
5.RT-PCR检测肺组织5-LO mRNA、15-LO mRNA水平:Genebank上检索大鼠β-actin和5-LO cDNA,15-LO cDNA序列,借助Primer Premier 5.0生物软件设计它们各自特异的引物序列,委托上海生工生物技术有限公司合成。内参照β-actin 引物:上游引物为5′-GCCAACCGTGAAAAGATG-3′,下游引物为5′-CCAGGACAGAGCCACCAAT-3′,PCR产物701 bp。5-LO 引物:上游引物为5′-GCGTTCAAGTGACTTCCATATC-3′,下游引物为5′-GGGATGTCCTCGGTGTTGT-3′,PCR产物336bp。15-LO 引物:上游引物为5′-GGCTGAGGTCTTTGCTG-3′,下游引物为5′-CGCTGTTGGGAATCCTGTCT-3′,PCR 产物长度为 363 bp。采用Trizol一步法抽提肺组织总RNA,紫外分光光度计上测RNA 的A260和A280值,计算RNA浓度,评价其纯度(A260/A280比值为1.8~2.0表明所提RNA有较好的纯度)。取1g/L总RNA 2μl进行反转录。以5μl反转录产物为模板,加入特异性引物和PCR反应体系,进行聚合酶链反应。15-LO反应参数:94℃变性60s,退火30s,72℃延伸30s,退火温度58.6℃,最适循环数36。5-LO反应参数:94℃ 变性 60s,退火30s,72℃延伸30s,退火温度61.0℃,最适循环数36。β-actin退火温度分别为58.6℃和61.0℃。最后,取PCR 产物各5μl,15g/L琼脂糖凝胶电泳分离鉴定,凝胶成像分析系统扫描后使用Gel-pro32软件进行图象分析,计算目标基因 RT-PCR产物荧光强度与β-actin产物荧光强度比值,作为目标基因 mRNA 的相对表达量。
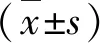
结 果
1.光镜下肺组织病理改变:哮喘组支气管、肺组织血管周围、肺间质及肺泡腔内可见大量炎性反应细胞,以嗜酸性粒细胞和肺泡巨噬细胞为主;支气管黏膜下水肿,黏膜皱褶增多,部分黏膜上皮细胞脱落,可见气道黏液栓。DXM组支气管肺组织炎性反应细胞浸润明显减轻,正常对照组未见异常表现。
2.各组大鼠肺组织5-LO mRNA和15-LO mRNA水平:与正常对照组比较,哮喘组大鼠肺组织5-LO mRNA水平显著上调,15-LO mRNA水平下降; 5-LO mRNA/15-LO mRNA比值升高,差异有统计学意义(P均<0.01)。地塞米松组较哮喘组,5-LO mRNA水平下降(P<0.01),15-LO mRNA水平上升(P<0.05),5-LO mRNA/15-LO mRNA比值下降(P<0.01)。5-LO mRNA、15-LO mRNA水平及5-LO mRNA/15-LO mRNA比值在地塞米松组与正常对照组比较,差异无统计学意义,详见图1、图2及表1。

图1 各组大鼠肺组织5-LO mRNA表达
A.正常对照组;B.哮喘组;C.DXM组

图2 各组大鼠肺组织5-LO mRNA表达
A.正常对照组;B.哮喘组;C.DXM组

表1 各组大鼠肺组织15-LO mRNA和5-LO mRNA水平
与正常对照组比较,*P<0.01;与哮喘组比较,#P<0.05,##P<0.01
3.血浆LXA4和OVA特异性IgE蛋白水平:哮喘组血清LXA4水平较正常对照组和地塞米松组升高,但差异无统计学意义(P>0.05)。 血清OVA-IgE含量在哮喘组明显升高,地塞米松干预后下降,差异有统计学意义(P均<0.01)。详见表2。
4.IgE相关性分析:各组大鼠血浆IgE与肺组织5-LO mRNA(r=0.879 ,P<0.01,图3),以及15-LO mRNA(r=-0.604 ,P<0.05,图4)均呈相关关系。当排除15-LO mRNA影响后,5-LO mRNA与IgE呈显著相关(r=0.814,P<0.01); 而当排除5-LO mRNA影响后,15-LO mRNA与IgE无相关关系(r=-0.248,P>0.05)。各组大鼠血浆IgE水平与血浆LXA4水平之间无显著相关性(r=0.267,P=0.243)。
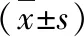
表2 各组大鼠血浆中LXA4和OVA特异性IgE蛋白水平
与正常对照组比较,*P<0.01;与哮喘组比较,#P<0.01

图3 大鼠血浆IgE与肺组织5-LO mRNA相关性

图4 大鼠血浆IgE与肺组织15-LO mRNA相关性
5.脂氧合酶与LXA4水平之间的关系:当5-LO mRNA/15-LO mRNA比值<1时,LXA4水平与5-LO mRNA/15-LO mRNA呈显著正相关(r=0.619,P<0.05,图5),当5-LO mRNA/15-LO mRNA比值>1时,LXA4水平与5-LO mRNA/15-LO mRNA呈显著正负相关(r=-0.879,P<0.01,图6)。

图5 LXA4与5-LO mRNA/15-LO mRNA相关性

图6 LXA4与5-LO mRNA/15-LO mRNA相关性
讨 论
支气管哮喘是由多种细胞,包括炎性细胞(嗜酸性粒细胞、肥大细胞、T淋巴细胞、中性粒细胞等)、气道结构细胞(气道平滑肌细胞和上皮细胞等)和细胞组分参与的气道慢性炎性疾病[7]。其发病机制复杂,涉及细胞因子、靶细胞、炎性介质等各个方面,但总的来说,是机体内环境稳态被打破而难以恢复的过程。近年来,花生四烯酸代谢产物半胱氨酰白三烯和脂氧素生成失衡受到关注,但对其中的机制研究甚少[8]。脂氧素是人体内源性产生的抗炎促消退物质,能拮抗白三烯引起的炎性浸润和气道收缩,被誉为白三烯的天然拮抗剂[9]。目前研究显示,半胱氨酰白三烯主要通过5-LO代谢途径生成;而脂氧素需要通过15-LO和5-LO的共同作用产生[2,3]。因而推测哮喘患者体内存在5-LO和15-LO的表达失衡。Wu等[10]通过对正常以及轻、中、重度哮喘患者的研究发现,随着哮喘程度的不断加重,患者白细胞15-LO mRNA的表达和血清LXA4水平逐渐下降;5-LO mRNA及血清LTB4、LTC4、IL-5、IL-8其表达逐渐升高,体现了哮喘患者中这种失衡的存在。但是,两者在哮喘动物模型中的表达情况还不清楚,15-LO和5-LO表达失衡对LXA4合成的影响研究极少。
本研究采用PCR法检测哮喘大鼠肺组织5-LO,15-LO表达水平,结果表明,与正常对照组比较,哮喘组大鼠肺组织5-LO mRNA水平升高,15-LO mRNA水平下降; 5-LO mRNA/15-LO mRNA比值升高,差异有统计学意义(P均<0.01);证实了在大鼠哮喘模型中也存在5-LO,15-LO表达失衡。糖皮质激素是哮喘治疗最有效的抗炎药物,用地塞米松进行模型干预后,出现5-LO,15-LO回复性调节,表明地塞米松的抗炎作用部分通过调节5-LO,15-LO表达平衡而实现。
过敏性哮喘是由IgE介导的Ⅰ型变态反应。本研究中哮喘大鼠组血清IgE明显升高,与5-LO呈显著正相关,这符合哮喘病理生理改变。15-LO在一定程度上对IgE具有负性调节作用,并受到5-LO的影响,体现在两者功能上的对立以及在代谢中的相互影响,因而其机制具有相当的复杂性,有待深入研究。无论如何,15-LO的抗炎作用是明确的。2008年开展的一项大规模基因调查研究就表明编码15-LO的基因ALOX15可能在过敏性哮喘中起保护作用[11]。本研究中地塞米松对15-LO的正面调控也进一步支持这种观点。为进一步探讨15-LO在过敏性哮喘中的保护作用是否通过其代谢产物LXA4的抗炎作用实现,本研究采用ELISA法检测了各组大鼠血清LXA4蛋白水平,结果发现,哮喘组大鼠LXA4水平较正常对照组升高,但差异无统计学意义;血清IgE水平与LXA4亦无明显相关性。提示15-LO的抗过敏作用可能独立于LXA4。
目前已经阐明,LXA4主要通过跨细胞途径合成,其中重要的一条是15-LO与5-LO合成途径[3]:上皮细胞、单核细胞或嗜酸性粒细胞内的15-LO在炎性细胞因子如IL-4、IL-13等的作用下上调,催化花生四烯酸的第15位C原子插入氧分子生成15S-HETE,然后被PMN吸纳,作为5-LO的底物生成 LXs。为了进一步探讨哮喘中5-LO与15-LO表达异常对LXA4合成的影响,本研究对LXA4和5-LO mRNA/15-LO mRNA 进行了相关性分析,结果发现,在一定范围内, 5-LO mRNA/15-LO mRNA的增大有助于LXA4的合成;超出这个范围后,LXA4的合成能力降低,并与5-LO mRNA/15-LO mRNA呈负相关。所以,5-LO、15-LO的表达失衡仍是LXA4合成不足的重要原因。
与Profita等[12]对哮喘患者的研究结论不同的是,在本研究中LXA4水平在哮喘组和正常组大鼠之间差异无统计学意义,这可能与研究对象在不同病理生理阶段5-LO、15-LO表达状态有关,纵向多时间点研究哮喘大鼠LXA4的表达,将有助进一步证实这种观点。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
江西医药(2018年8期)2018-10-24
中学生数理化·教与学(2017年4期)2017-04-22
罕少疾病杂志(2017年2期)2017-02-23
西南军医(2016年6期)2016-01-23
中国医学影像学杂志(2015年9期)2015-12-15
医学研究杂志(2015年12期)2015-06-10
中国民族民间医药·下半月(2014年2期)2014-09-26
中国中医药现代远程教育(2014年13期)2014-03-01
