
黄顶菊对入侵地群落动态及植物生长生理特征的影响
2019-08-20 10:58祁小旭王红岩张思宇皇甫超河杨殿林
生态学报 2019年12期
祁小旭,王红岩,林 峰,张思宇,王 慧,皇甫超河,*,杨殿林
1 沈阳农业大学植物保护学院, 沈阳 110866 2 农业部环境保护科研监测所, 农业农村部产地环境污染防控重点实验室/天津市农业环境与农产品安全重点实验室, 天津 300191 3 天津农学院农学与资源环境学院, 天津 300384
生物入侵严重威胁入侵区域的生物多样性,是21世纪五大全球性环境问题之一,是影响全球生物多样性的第二大因素[1- 3]。随着全球化进程的日益加剧,越来越多的外来物种被有意无意地带入新的生境或区域,增加了外来物种对入侵地的入侵风险[4]。外来植物入侵是生物入侵的重要组成部分,最近的研究表明,外来入侵植物主要通过改变入侵地土壤物理化学及生物学特性、土壤养分循环、群落多样性等从而在入侵地迅速地繁殖,对入侵区域的生态环境、社会经济和人类健康造成了严重威胁[5]。例如:银荆(Acaciadealbata)在被入侵地生态系统中造成了不同程度的损害,包括严重减少当地的物种多样性[6- 7],改变土著植物的分解过程[8],改变土壤生物化学成分[9],土壤微生物群落或土壤微生物功能的变化[10]等。
黄顶菊(Flaveriabidentis)属菊科(Asteraceae)万寿菊族(Tageteae)黄菊属(Flaveria),一年生草本植物,原产于南美洲[11]。2001年黄顶菊在我国河北省衡水湖地区和天津市首次被发现以来,目前已扩散至河北、天津、河南、山东等省市,以河北省发生面积最大,广泛分布于该省的70多个县市区,并且有不断扩散的趋势[12],给当地农林业生产以及环境带来巨大危害[13]。目前研究认为,黄顶菊有较强的环境适应能力,耐盐碱和干旱等胁迫条件。其生态幅广泛,根部能分泌化感物质抑制其他植物生长,获取竞争优势,对入侵地的生态环境造成威胁,尤其是其排它性的生长特性对及生物多样性带来严重影响,危及生物生态系统安全[14],被称为 “生态杀手”[15]。
黄顶菊对土壤生态系统影响研究表明,黄顶菊改变了土壤的养分循环[16],黄顶菊的入侵增加了土壤有机质和全氮的含量,降低了pH和硝态氮的含量[17],其影响机制主要是改变了微生物群落结构、功能和相关酶活性等[18]。黄顶菊入侵土壤氨氧化细菌多样性下降,氨氧化古菌[19]、固氮菌、磷细菌和钾细菌多样性及丰富度升高[20]。尽管如此,目前对黄顶菊研究多侧重其对入侵地土壤生态系统的影响,对其入侵对本土植物群落结构的影响和种间竞争机制研究鲜有报道,尤其是对入侵植物与本土群落动态监测研究报道更为缺乏。通常野外观测和采样研究结果受微环境影响较大,本研究采用同质园试验模拟野外生长条件,对黄顶菊入侵和未入侵群落开展整个生育期动态监测,以期探求黄顶菊对入侵地整体植物群落和土著植物的生理生长特征的影响规律,研究拟验证以下假设:(1)黄顶菊入侵降低入侵地群落物种多样性水平;(2)黄顶菊入侵主要影响本土植物的生长生理特性,且不同本土植物对黄顶菊入侵的耐受性存在物种差异。该研究能够丰富黄顶菊的入侵机制,为入侵植物生态控制提供一定的理论依据。
1 材料与方法
1.1 试验设计和试验用土采集
本研究为同质园试验,试验用土采集于天津市静海团泊北堤(38°54′04″N, 117°08′46″E,海拔2.4 m),处于暖温带大陆性季风气候区,年均气温11.8℃,年均降水量580 mm,集中于6—8月,无霜期193 d,土壤为碱性潮土。入侵地伴生植物为羽叶鬼针草(BidensmaximowiczianaOett)、狗尾草(Setariaviridis(L.) Beauv)等;本地植物群落一般由反枝苋(AmaranthusretroflexusL.)、牵牛(Pharbitisnil(Linn.) Choisy)、灰绿藜(ChenopodiumglaucumL.)、地肤(Kochiascoparia(L.) Schrad)、芦苇(Phragmitesaustralis(Cav.) Trin. ex Steu)和猪毛菜(SalsolacollinaPall)等组成[19]。
于2016年12月采集非入侵地和入侵地的土壤(入侵样地入侵时间大于15年;非入侵地无黄顶菊入侵)。入侵地和非入侵地中间距离10 m或更少,确保两种土壤主要差异为是否有黄顶菊生长,取样点距离植物群落边缘至少有3 m。非入侵地和入侵地分别设置3个15 m的南北向平行样线,作为3个取样区,每样线间隔3 m,每个样线上设置5个1 m × 1 m取样样方,因此,本研究中总共划定6个取样区,30个取样样方。在每个样方中用铁锹取出土壤,取样深度15 cm,识别并除去植物组织,收集土壤,每个土壤样品中包含土壤和细根,入侵地和非入侵地每个取样区内土样单独混匀,因此每个取样区内的土壤样品可作为一个试验重复,共3次重复。每个取样区内的土壤样品分别装入5个种植箱(22 kg/箱)(规格50 cm × 30 cm × 15 cm,北京丰瑞佳业有限公司),入侵地和非入侵地分别装15个种植箱,总共装30个种植箱(2种土壤类型 × 5次监测 × 3次重复),并编号以区分重复(N1.1,N1.2,N1.3,N1.4,N1.5;N2.1,N2.2,N2.3,N2.4,N2.5;N3.1,N3.2,N3.3,N3.4,N3.5;F1.1,F1.2,F1.3,F1.4,F1.5;F2.1,F2.2,F2.3,F2.4,F2.5;F3.1,F3.2,F3.3,F3.4,F3.5,其中N为非入侵土壤,F为入侵土壤),将这30个种植箱放置在农业部环境保护科研监测所实验网室(天津,39°05′49″N,117°08′46″E,海拔4 m)过冬(模拟野外条件)直至试验开始。供试入侵地和非入侵地的本底土壤基本理化见表1。

表1 本底土壤理化性质(平均值±标准误)
同一列中,相同字母表示差异不显著(P> 0.05),不同字母表示差异显著(P< 0.05)
在2017年5月3日起,每天向箱子内加入自来水,保持土壤持水量在70%水平,促使土壤中种子库自然萌发。此时亦为野外黄顶菊出苗时期。待种子库萌发出苗后水分保持在田间持水量的40%左右。在试验期间不进行间苗处理,且种植箱每3天在网室内随机放置和旋转,避免微环境变化对植物生长的影响。
1.2 植物群落的清查
从2017年6月10日起,分别于2017- 6- 10、2017- 6- 20、2017- 6- 30、2017- 7- 10、2017- 7- 20进行不同群落种植箱植物个体计数,每次计数在每个重复内的5个种植箱内随机选取一个种植箱,每次计数在不同土壤中选择3个种植箱,例如:N1.2,N2.1,N3.5;F1.4,F2.2,F3.1。由于种植箱内植物密度很大(每个种植箱超过100株),因此每个种植箱被分成4个25 cm × 15 cm的小样方,作为植物群落清查的工作单元。随机选取一个小样方记录物种数据(即物种名称和株数)。收集的植物种类根据它们的性状(本地种或入侵种)、生活型(禾草类或非禾草类)进行分类。基于17个覆盖类别(百分比: 0.01%, 0.2%, 0.5%, 1%, 3%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 98%和100%)估计每物种覆盖度。物种的命名和本地种/入侵种的定义遵循USDA Plants (USDA NRCS 2017)。我们通过测量每个小样方中的“平均”植被高度来估算植被的相对高度,然后求每个样方中的平均相对高度。
重要值(important value)是衡量一个群落中物种的生长和相对优势度的综合量化指标,并根据相对盖度、相对高度和相对丰度进行计算[21-22],公式如下:
重要值=(相对盖度+相对高度+相对丰度)/3
式中:
相对盖度=某一物种的盖度/所有物种的盖度总和
相对高度=某一物种的高度/所有物种高度总和
相对丰度=某一物种的个体数/所有物种的个体数总和
α多样性指数是用来描述物种丰富度和均匀度的指标,Patrick 丰富度指数指示植物群落的总丰富度,Simpson多样性指数、Shannon-Wiener 多样性指数表征植物群落内部的多样性变化,Simpson多样性指数对于群落均匀度的敏感性高于 Shannon-Wiener 多样性指数,而Shannon-Wiener 多样性指数对于不同物种丰富度的敏感性高于Simpson多样性指数,Pielou 均匀度指数指示植物群落的总均匀度,物种丰富度越高,各种个体分配越均匀,Patrick 丰富度指数、Simpson 多样性指数、Shannon-Wiener 多样性指数、Pielou 均匀度指数越高,指示群落多样性越好[23- 25]。各参数计算公式如下:
Patrick 丰富度指数:
R=S
Simpson 多样性指数:λ= 1 - ∑Pi2
Shannon-Wiener 多样性指数:H= -∑PilnPi
Pielou 均匀度指数:E= (-∑PilnPi)/lnS
式中,S是一个种植小区内的物种数,Pi是某一个物种的相对丰度。
同时,用Li-3100C叶面积仪(Biosciences, Lincoln, Nebraska, USA)测定新鲜叶片叶面积。清查后按种植箱和物种收获植物地上生物量之后把每个植株茎叶分开放入烘箱内烘干(48 h(叶)、72 h(茎),70℃),称取干重,计算比叶面积(SLA;叶面积/干叶质量;cm2/g),在每个种植箱内选取代表性的单株,每个种植箱测定10个叶片,求其平均值,3次重复。
1.3 植物生长动态的监测-本地主要物种
选取在本地和入侵地共同存在的4种优势本土物种(狗尾草(Setariaviridis)、羽叶鬼针草(Bidensmaximowicziana)、灰绿藜(Chenopodiumglaucum)、地肤(Kochiascoparia)做代表性本地共生物种的研究,特别地,开展黄顶菊与近缘植物(如羽叶鬼针草)比较研究,利于揭示入侵种独特的环境适应性和入侵策略。出苗约12周后,在2017年7月12—13日,植物营养生长的高峰期时,每个处理中随机选取代表性单株进行气体交换参数的测定。为了尽量减少边界效应,植物生长和生理特征的测定都是尽可能用中心区域植物进行测定(距离箱缘5 cm,中心面积为0.062 m2)。气体交换参数原位测定用LI- 6400XT便携式光合仪(Li-Cor Inc, Lincoln, USA)。净光合速率(Pn)在08:30—11:30测定,参比室CO2浓度设定在400 μmol/mol。饱和光子通量密度设定为1200 μmol/m2s1。叶室温度25℃,湿度60%,CO2浓度380 μmol/mol。同时得到蒸腾速率(Tr)、气孔导度(Cd)和细胞间CO2浓度(Ci)等数据,在每个种植箱内选取代表性的植株,每个种植箱测定10个叶片,求其平均值,3次重复。
为了研究黄顶菊入侵对伴生土著物种根系形态学的影响,对不同处理的种植箱内的植株测定了比根长(SRL)和比根面积(SRA)。用于生理指标测定植株收获后随机抽取4个大约10 cm长的细根(直径<2 mm),用自来水冲洗掉细根表面的土壤。所有的细根样品在400 dpi的根系扫描仪上扫描(model J181A,Epson Expression 10000XL flatbed scanner, Regent Instruments, Quebec City, Quebec, Canada),用Winrhizo软件(v. 2009, Regent Instrument, Quebec, Canada)确定根长(L,cm)和根表面积(SA, cm2)。随后,烘干(70℃, 48 h)细根并称量细根的生物量(DM, g)。在每个种植箱内选取3株有代表性的单株,求其平均值,3次重复。SRL和 SRA计算公式如下:
SRL(cm/g)=L/DM
SRA(cm2/g)=SA/DM
1.4 土壤理化性质的测定
于2017年7月20日群落清查之后,用直径5 cm的土钻采集土壤,每个种植箱按照“五点取样法”采集5钻土壤样品,每个种植箱内单独混匀,取3个种植箱,采样深度15 cm,土壤采集后测定理化性质。土壤有机质测定采用重铬酸钾-浓硫酸外加热氧化法;土壤全氮采用浓硫酸消煮法,之后采用流动分析仪(AA3,Bran+Luebbe Crop,德国)测定;土壤硝态氮、土壤铵态氮采用氯化钙浸提法,之后采用流动分析仪测定,土壤全磷采用钼锑抗比色法;土壤pH采用玻璃电极法(MP511 pH计)按土水比 1∶2.5 测定。
1.5 数据分析
试验数据用SPSS 21.0 软件处理,对Patrick丰富度指数,Simpson 多样性指数、Shannon-Wiener 多样性指数、Pielou均匀度指数、株高、株数、生物量、重要值等先进行方差齐性检验,然后采用重复测量方差分析(repeated-measure ANOVA)样地类型(入侵地和非入侵地)和取样时间(5次:2017- 6- 10、2017- 6- 20、2017- 6- 30、2017- 7- 10、2017- 7- 20)及其交互作用对群落动态的影响。对入侵地和非入侵地处理间的群落动态(Patrick丰富度指数、Simpson 多样性指数、Shannon-Wiener 多样性指数、Pielou均匀度指数)、生长指标(株高、株数、生物量、重要值)、生理指标(净光合速率、气孔导度、蒸腾速率、胞间CO2浓度)及根系形态学指标(比叶面积(SLA)、比根长(SRL)、比根面积(SRA))等差异显著性分析采用独立样本t检验,差异显著性水平设为α=0.05。结果图表用SigmaPlot 13.0、WPS Excel 2016绘制,数据为平均值±标准误。
2 结果与分析
2.1 土壤理化因子
黄顶菊对入侵地土壤理化因子产生了显著的影响(表2)。入侵地土壤中有机质含量和全氮含量显著高于非入侵地即本地土壤,分别高34.47%和114.29%,说明黄顶菊的入侵增加了入侵地土壤的养分水平;而硝态氮、铵态氮、全磷、pH的呈下降趋势。

表2 黄顶菊入侵对土壤理化因子的影响(平均值±标准误)
同一列中,相同字母表示差异不显著(P> 0.05),不同字母表示差异显著(P< 0.05)
2.2 本地植物群落多样性
除Patrick丰富度指数外,Simpson 多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数在不同处理下随时间变化显著(表3,P<0.001),显示黄顶菊对本地植物群落的多样性指标影响表现出季节差异。

表3 黄顶菊入侵对植物群落影响的重复性测量方差分析
黄顶菊入侵对本地植物群落生长的影响随时间的变化如图1所示:Patrick丰富度指数、Simpson 多样性指数、Shannon-Wiener 多样性指数和Pielou均匀度指数本地土壤均高于入侵土壤,且随时间的变化差异逐渐减小,Simpson多样性指数、Shannon-Wiener 多样性指数在生育期内均有显著性差异(P<0.05),本地土壤中生长的植物群落4种指数在生育期内没有显著变化,但是在入侵地生长中4种指数在生育期内逐渐升高,在生育后期差异逐渐减小,两种土壤中生长的Patrick丰富度指数、Pielou均匀度指数在生育后期没有显著差异(P>0.05)。说明黄顶菊入侵主要影响入侵地群落多样性,且有显著的时间效应,随生育期推进影响逐渐减小。
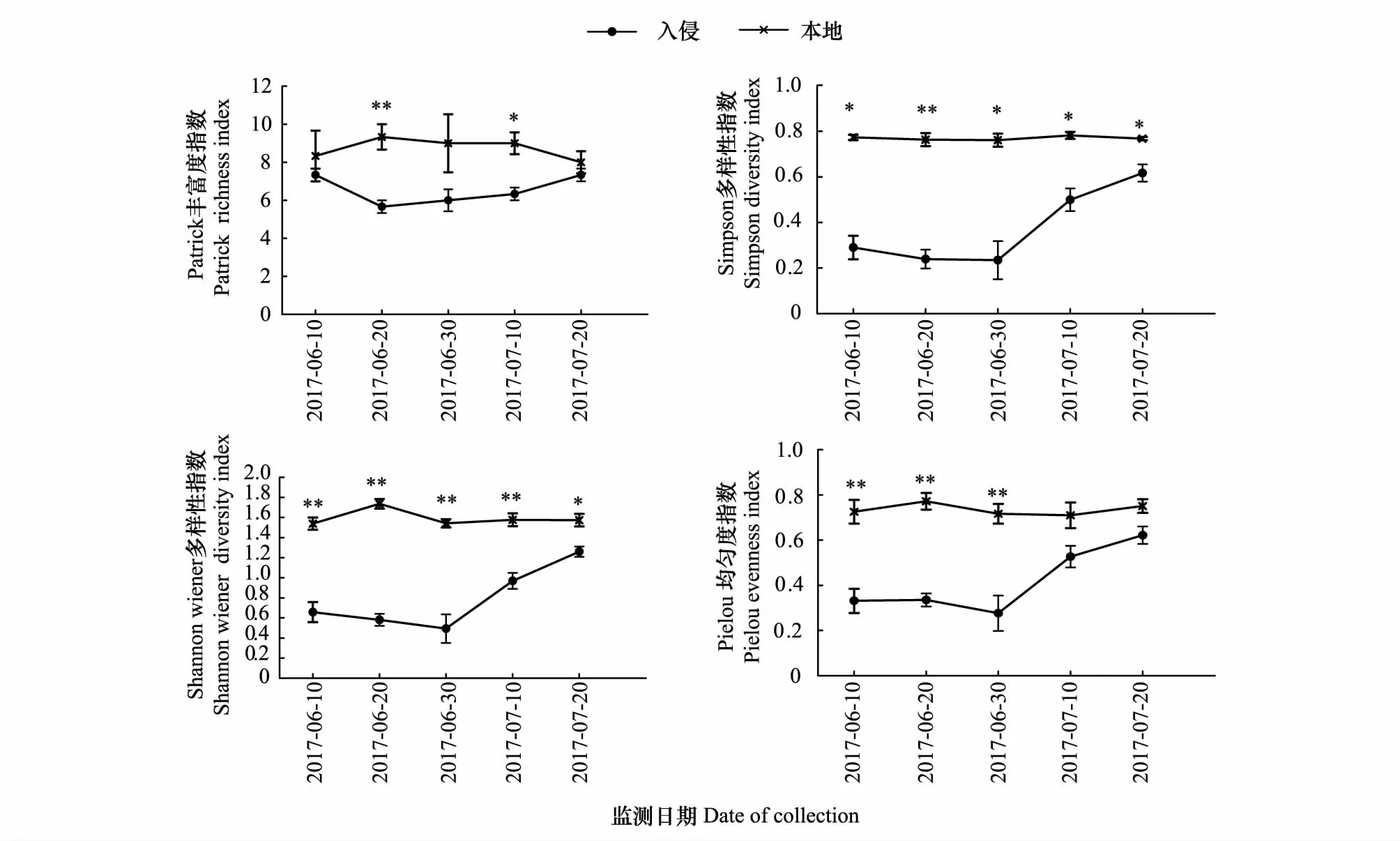
图1 黄顶菊入侵对植物群落的影响Fig.1 The effect of Flaveria bidentis invasion on plant community variables数值为平均值±标准误,其中标*为不同处理达到显著性差异(P<0.05),标**为不同处理达到极显著性差异(P<0.01)
2.3 本地主要物种的生长指标
在不同处理下:狗尾草、羽叶鬼针草、灰绿藜、地肤在株高、株数、生物量等指标在不同处理下随时间变化显著(表4,P<0.001),显示黄顶菊对本地植物的株高、株数、生物量指标影响表现出季节差异;在重要值方面,除地肤外,狗尾草、羽叶鬼针草、灰绿藜在不同处理下随时间变化显著(P<0.01),黄顶菊入侵对地肤重要值的影响没有季节差异。
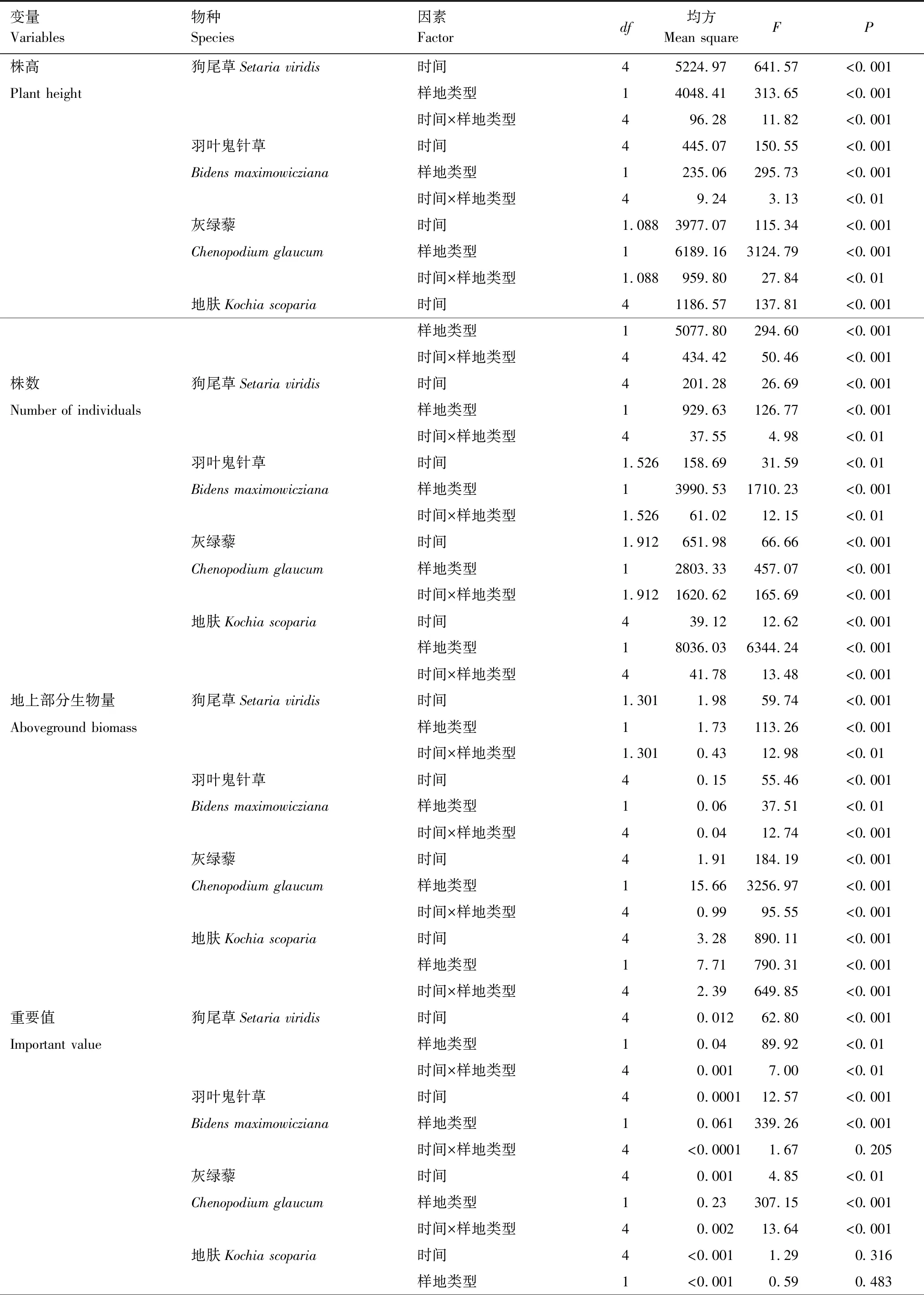
表4 黄顶菊入侵对本地主要物种生长影响的重复性测量方差分析
黄顶菊入侵对4种本地植物株高的影响随时间的变化如图2所示,由图可以看出:4种植物本地生长的株高均显著高于入侵地(P<0.05)。狗尾草株高的差异在五个时期均达到极显著差异(P<0.01),且随时间的变化差异有先增加后减小的趋势;羽叶鬼针草、灰绿藜、地肤三个物种在入侵地和非入侵地的差异随生育期的推进有不断增加的趋势。
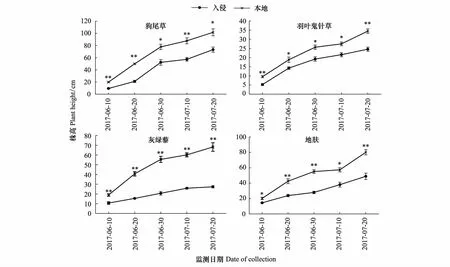
图2 黄顶菊入侵对本地物种株高的影响Fig.2 The effect of Flaveria bidentis invasion on plant height of the native species数值为平均值±标准误,其中标*为不同处理达到显著性差异(P<0.05),标**为不同处理达到极显著性差异(P<0.01)
黄顶菊入侵对4种本地植物株数的影响随时间的变化如图3所示:狗尾草和地肤在入侵地的株数在五个时期都比在本地的株数高,且达到显著性差异(P<0.05),相反,羽叶鬼针草和灰绿藜在本地土壤生长的株数均显著大于在入侵地株数(P<0.05)。
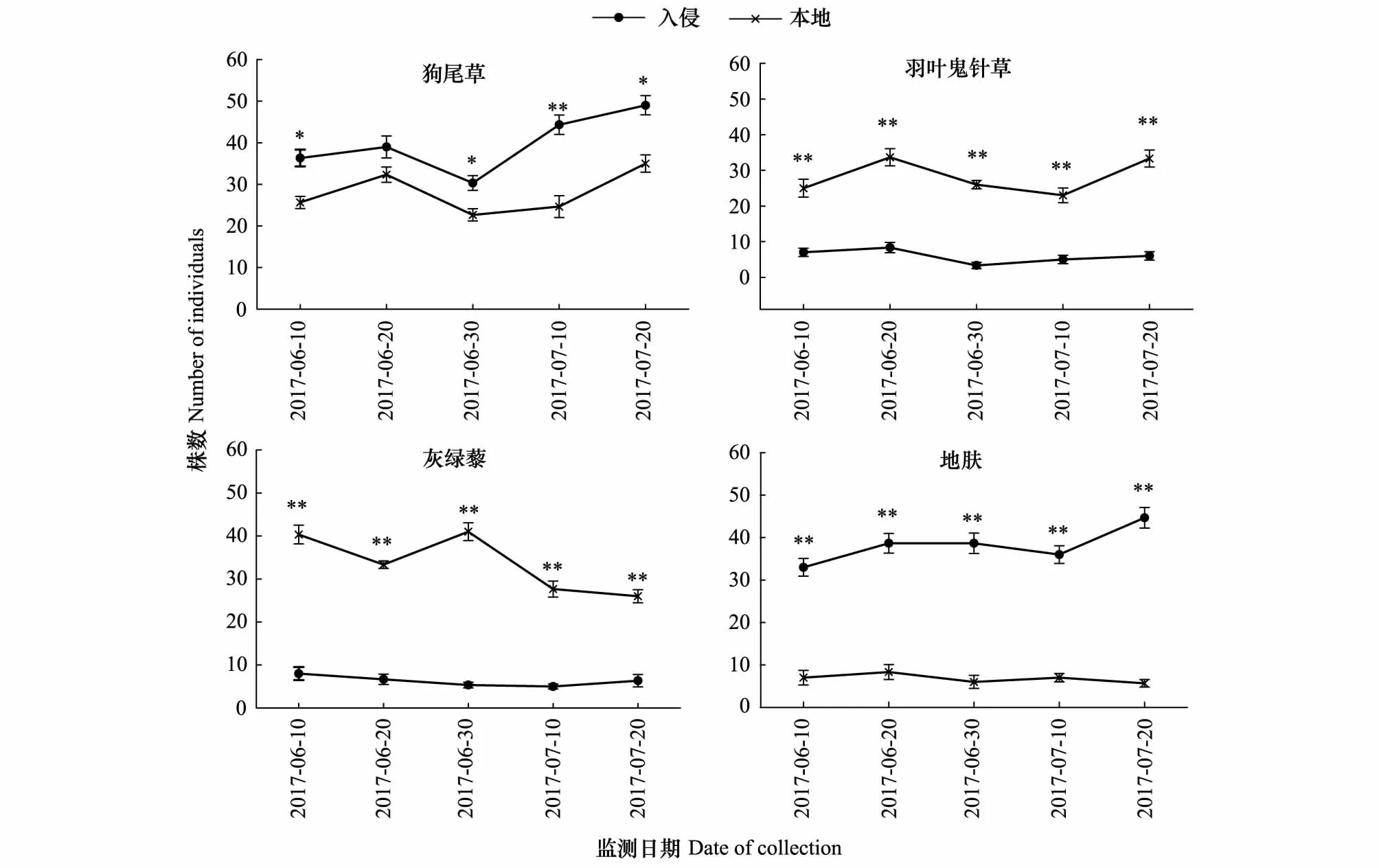
图3 黄顶菊入侵对本地物种株数的影响Fig.3 The effect of Flaveria bidentis invasion on individual number of the native species数值为平均值±标准误,其中标*为不同处理达到显著性差异(P<0.05),标**为不同处理达到极显著性差异(P<0.01)
黄顶菊入侵对狗尾草、灰绿藜、地肤3种本地植物地上生物量的影响随生育期推进影响逐渐增强,且有极显著差异(P<0.01),由图4可见,虽然羽叶鬼针草在生育初期本地土壤地上生物量与入侵土壤相当(P>0.05),但在生长后期入侵土壤生物量显著高于本地土壤112%(P<0.01)。
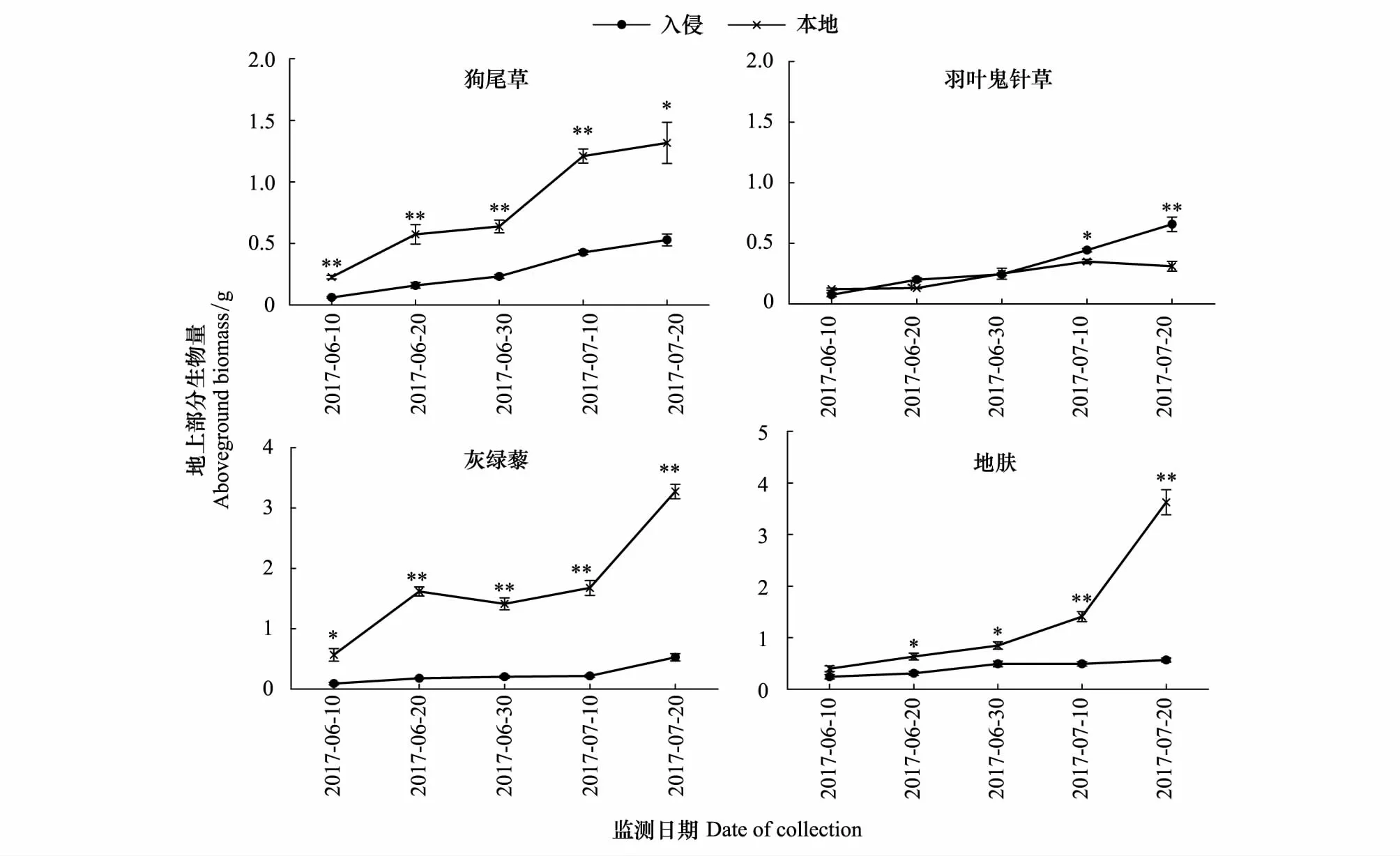
图4 黄顶菊入侵对本地物种生物量的影响Fig.4 The effect of Flaveria bidentis invasion on aboveground biomass of the native species数值为平均值±标准误,其中标*为不同处理达到显著性差异(P<0.05),标**为不同处理达到极显著性差异(P<0.01)
狗尾草、羽叶鬼针草、灰绿藜的重要值在本地土壤显著高于入侵土壤(图5,P<0.05),例如,生长初期本地土壤狗尾草比入侵土壤高86%(P<0.01),生长后期本地土壤生长比入侵土壤高25.5%(P<0.05),且差异有逐渐减小的趋势;地肤的重要值在五个时期内只有第四个时期有显著性差异(P<0.05),且狗尾草、羽叶鬼针草、灰绿藜在整个生育期内重要值的平均值本地土壤显著高于入侵地(P<0.05),分别高:50.3%,120%,202%,地肤在两种处理下没有显著差异。
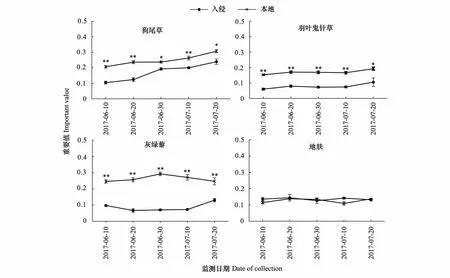
图5 黄顶菊入侵对本地物种重要值(IV)的影响Fig.5 The effect of Flaveria bidentis invasion on important values (IV) of the native species数值为平均值±标准误,其中标*为不同处理达到显著性差异(P<0.05),标**为不同处理达到极显著性差异(P<0.01)
2.4 本地主要物种的生理指标
由图6可以看出,狗尾草,羽叶鬼针草,灰绿藜,地肤在本地生长的净光合速率、气孔导度、蒸腾速率显著高于入侵土壤(P<0.05),净光合速率分别高31%、23%、123%、46%;气孔导度分别高44%、51%、122%、57%;蒸腾速率分别高106%、11%、65%、39%,胞间CO2浓度两种处理下没有显著性差异(P>0.05)。说明黄顶菊入侵对本地植物的生长有显著抑制作用,但存在物种差异。
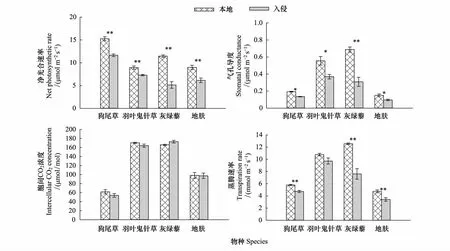
图6 黄顶菊入侵对本地植物光合特性的影响Fig.6 The effect of Flaveria bidentis invasion on photosynthesis characteristics of the native plants数值为平均值±标准误,其中标*为不同处理达到显著性差异(P<0.05),标**为不同处理达到极显著性差异(P<0.01)
根的结构或形态,如特定的根长和特定的根表面积,可以反映土壤资源的变化并与周围植物竞争密切相关。将比根长(SRL)和比根面积(SRA)计算为根长度和根表面积对根质量的影响,数值越大,根对土壤营养的吸收效率越高。由图7可以看出,狗尾草,羽叶鬼针草,灰绿藜、地肤在入侵地土壤生长的SRL均高于本地土壤,灰绿藜和地肤在两种土壤中的差异达到显著差异(P<0.05);狗尾草,羽叶鬼针草,灰绿藜、地肤的比根面积(SRA)入侵地土壤显著高于本地土壤(P<0.05);狗尾草在入侵地土壤的比叶面积(SLA)显著高于本地土壤(P<0.01)。
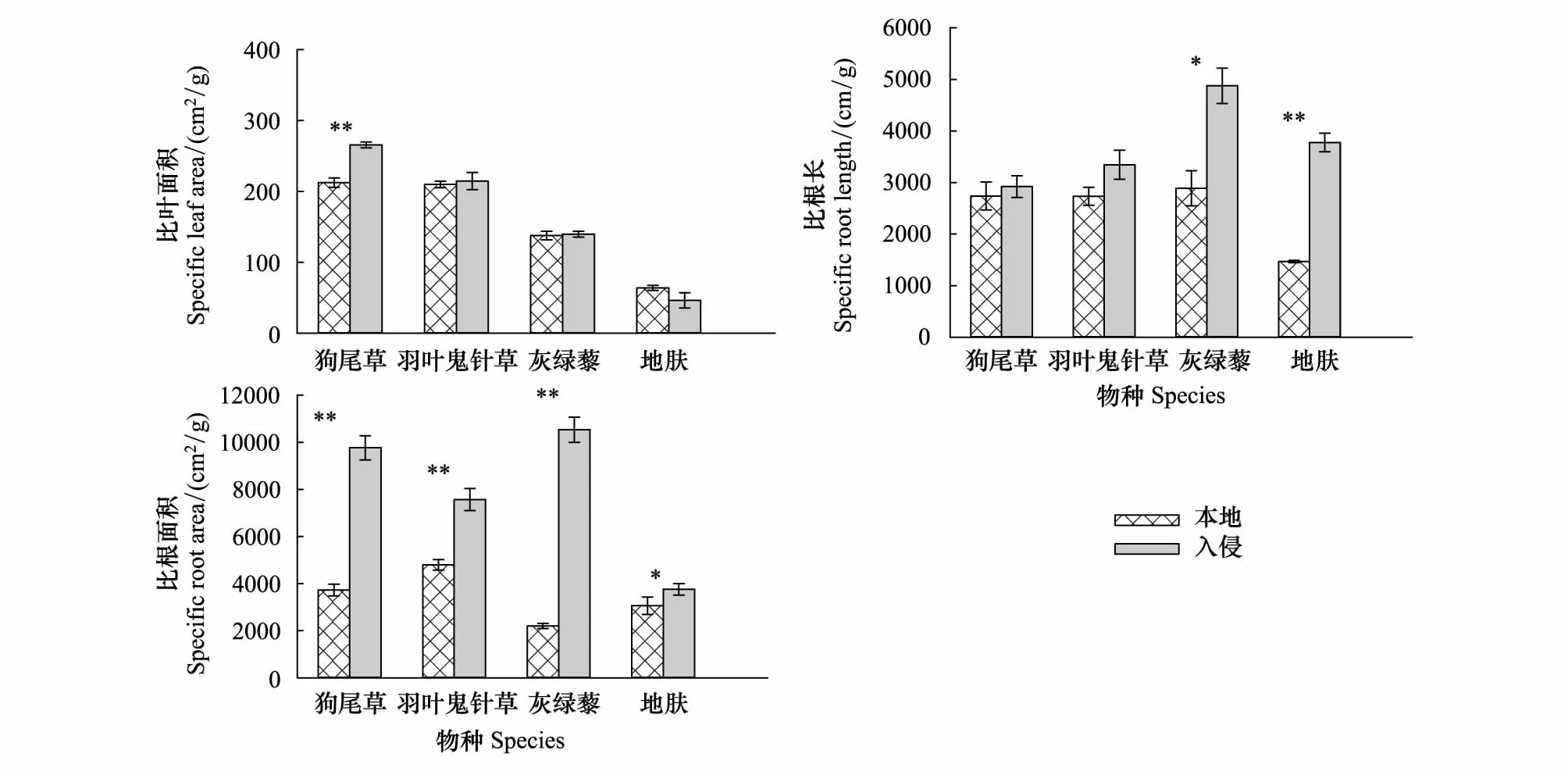
图7 黄顶菊入侵对本地植物叶、根特性的影响Fig 7 The effect of Flaveria bidentis invasion on leaf and root characteristics of the native plants数值为平均值±标准误。其中标*为不同处理达到显著性差异(P<0.05),标**为不同处理达到极显著性差异(P<0.01)
3 讨论
植物入侵可以改变自然群落生态系统基本生物学特征[26- 27],降低入侵地植物群落的丰富度和多样性[28- 29],本文研究了黄顶菊对入侵地植物群落以及4种土著植物生理生长特征的影响规律,结果表明,黄顶菊降低了入侵地群落物种多样性水平,而随着生育期推进,其对入侵群落丰富度和均匀度指数的影响逐渐减小;黄顶菊对4种土著植物的生长均有抑制作用,但这种抑制作用存在种间差异。
3.1 黄顶菊对入侵地土壤理化因子的影响
前人研究表明,植物入侵可以通过对氮素养分的选择性吸收[30]或凋落物输入和根系分泌物等[31- 32]方式改变土壤理化特性,而入侵地土壤养分改变又能创造对入侵种有利的条件来实现进一步入侵[33- 35]。本研究中,黄顶菊的入侵,增加了有机质和全氮含量,而硝态氮、铵态氮、全磷、pH呈下降趋势(表2),这和赵晓红等[14]的研究结果一致,与张天瑞等[12]的研究结果和本底数据(表1)显示铵态氮增加的结果有所不同,可能由于土壤采集时间为植物的生长盛期,养分利用能力更强,因此铵态氮的含量表现下降,而土壤pH下降主要是由于黄顶菊对铵态氮素选择性吸收引起的[30]。
3.2 黄顶菊入侵对本地植物群落多样性的影响
“增强竞争力进化假说”认为:外来物种被引入后由于缺少天敌的控制作用与竞争作用,更多地把资源用于自身的生长发育,增强自身竞争力[36]。例如:互花米草(Spartinaalterniflora)由于在入侵地竞争力显著高于土著物种,这种不对称竞争导致了土著植物种群分布面积的减少和种群数量明显降低[37]。本研究结果表明,在整个生育期内群落Patrick丰富度指数、Simpson 多样性指数、Shannon-Wiener 多样性指数和Pielou均匀度指数在非入侵土壤高于入侵土壤,说明黄顶菊的入侵降低了入侵地植物群落的多样性,此实验结果验证了前言中假设一;但随时间的推进这种差异逐渐减小,在试验后期(2017- 7- 20之后),入侵土壤植物群落Patrick丰富度指数和Pielou均匀度与本地土壤植物群落已无显著差异,马克平等[23]认为Simpson指数对于群落均匀度的敏感性高于Shannon-Wiener指数,而Shannon-Wiener指数对于不同物种丰富度的敏感性高于Simpson指数。本文结果显示:试验前期,Shannon-Wiener 多样性指数高于Simpson 多样性指数,且差异大于Simpson 多样性指数,表3显示Patrick丰富度指数随时间变化不显著,而其他三个多样性指数随时间变化显著,且4种本地植物的丰富度随时间变化显著,不同物种丰富度的变化最终会导致植物群落均匀度的变化,因此可以说明黄顶菊的入侵主要植物群落的均匀度。试验后期Pielou 匀均度指数已无显著差异,显示入侵土壤植物群落表现恢复趋势。入侵种对本地物种抑制作用通常认为与其较高郁闭度导致的对本地植物生长遮蔽效应有关[38- 39],黄顶菊由于出苗时间普遍早于其他物种,与其他物种的竞争作用小,可以将更多的资源用到自身的生长发育,当其他物种出苗时,黄顶菊植株已经形成覆盖,这可以从黄顶菊在群落中较高的覆盖度得到证实。加之其株高普遍高于多数共生本地种,能够促使其获得竞争优势的同时形成对后者光资源阻断,抑制后者生长。其次,化感作用可能也是导致入侵群落多样性降低的重要因素。已有研究表明黄顶菊能够通过凋落物分解[40]和根系分泌次生代谢物质[41- 42]抑制共生本地植物生长。生长后期入侵群落多样性指数恢复可能是由于耐弱光照环境和对黄顶菊化感作用表现耐受物种增加所致[43]。因此,可以推测黄顶菊入侵将改变群落物种组成,而对其入侵效应表现耐受的物种得以共存,但需要长期跟踪监测研究证实。
此外,凋落物输入也被认为是影响黄顶菊入侵地土著物种生长的重要因素[44- 45],本研究为一个生长季未考虑凋落物添加影响,因此可能低估了黄顶菊入侵对本地植物群落影响。
3.3 黄顶菊入侵对4种本地物种生长指标的影响
株高、生物量、株数、重要值[21- 22]是植物生长的重要生长指标,表征植物的生长状况。入侵植物与本地植物竞争光照等资源,抑制本地植物的生长,影响本地植物的株高、覆盖度、生物量,但对于本地植物优势种的株数没有显著影响[46],Florianová 等的研究结果表明,凤仙菊(Impatiensparviora)的入侵减少了入侵地土著植物的株高和覆盖度,对丰富度的影响不大[47]。本研究结果表明(图1—4),4种植物在本地土壤生长的株数、生物量显著高于入侵土壤(P<0.05),随生育期的推进差异逐渐变大,但不同物种之间表现差异。株高是影响植物获取光资源的主要因素,株高越高,植物获得光资源越丰富,可以合成更多利于自身生长的物质,因此可以积累更多的生物量。结果显示入侵地4种植物的株高显著低于非入侵地,且差异逐渐增大,说明土著植物与黄顶菊的竞争过程中由于光资源的获得减少,因此在入侵地所积累的生物量显著低于非入侵地(P<0.05)。有研究表明[12]黄顶菊分泌的化感物质可以抑制其伴生物种种子的萌发,因此在入侵地鬼针草和灰绿藜的丰富度显著小于非入侵地,可能是由于化感作用对本地植物种子萌发的影响,但其内部影响机制还需要进一步研究。黄顶菊对本地植物生长指标的影响,可能导致植物群落中喜光物种逐渐消失,耐弱光物种互逐渐增多,因此改变了植物群落的物种组成,使群落多样性指标随时间的变化差异逐渐减小。重要值是由相对高度、相对丰度、相对盖度计算的表征植物优势度的综合指标[21- 22],狗尾草、羽叶鬼针草、灰绿藜在入侵土壤的重要值显著小于本地土壤(P<0.05),地肤的重要值在两种土壤中没有显著差异(P>0.05)。因此,本实验的实验结果表明入侵植物对本地植物的主要影响为对光资源的竞争,支持增强竞争力假说[36],且本部分实验结果验证了假设二中的黄顶菊对本地植物的影响存在物种差异。
3.4 黄顶菊的入侵对本地植物生理指标的影响
植物的光合作用影响了植物在生长过程中的竞争能力[48],净光合速率则是植物光合机构功能效率和初级生产力高低的指标[49]。入侵植物与本地土著植物相比具有较高的资源利用能力[50],加之黄顶菊在群落中有较高的覆盖度,能够促使其获得竞争优势的同时形成对本地种光资源阻断,形成遮蔽效应,抑制本地物种生长[17]。本文研究结果表明(图6)4种植物的净光合速率、气孔导度、蒸腾速率在本地土壤生长显著高于入侵土壤,此部分实验结果验证了假设二。可以说明4种本地物种在入侵地由于有黄顶菊的遮蔽作用,所获得的光资源受到抑制,因此光合作用受到抑制,因此气孔导度、蒸腾速率在入侵土壤生长显著低于本地土壤,导致植物的净光合速率显著低于非入侵土壤(P<0.05)。在入侵地4种植物的光合作用受到显著抑制,所合成的碳水化合物显著低于非入侵土壤,因此4种本地植物在入侵地所积累的生物量显著低于非入侵土壤。
SLA是重要的叶片功能性状之一[51],较薄的叶片的植物一般具有较高叶片SLA,在合成纤维素、半纤维素、果胶等自身结构性碳水化合物时可以投入较少碳,叶片建成成本较低[52]。植物根的形态特征如比根长(SRL)、比根面积(SRA)能够反映土壤营养的变化[53- 54],表征植物与其他植物的竞争能力[55],因此可作为植物生长状况的参考指标。最优分配理论认为,植物在受到土壤内胁迫时,会增加根系资源的分配,在有限的空间和资源水平下获得了更多的土壤养分和水分,应对这种胁迫[56]。周雨露等认为本地植物为了应对入侵胁迫,利用自身特性即根部形态和叶部形态增加了资源利用效率,缓解抑制作用[57],本文结果表明入侵土壤的SLA显著高于本地土壤(P<0.05)(图7),由于黄顶菊的入侵胁迫,本地植物在入侵地可获得的光资源减少,影响本地植物进行光合作用,而高SLA会极显著增强植物净光合速率[51],因此本地植物为了减缓这种胁迫,进而将叶部形态向高SLA转化,提高光能利用效率,进而合成更多的有机物质用于自身的生长发育减缓黄顶菊的入侵胁迫。
狗尾草、羽叶鬼针草、灰绿藜、地肤的SRL和SRA在入侵土壤高于本地土壤(图7),显然是由于其为应对黄顶菊入侵这种胁迫因素,增加了根系资源的分配,使本地植物在这种胁迫作用下利用有限的空间和资源最大限度地吸收更多的养分和水分,维持自身的生长发育,缓解抑制作用。
综上所述,本文从植物生理和生长方面研究了黄顶菊入侵对本地植物生长的影响,结果表明4种本地物种在入侵地由于有黄顶菊的遮蔽作用,所获得的光资源受到抑制,气孔导度、蒸腾速率在入侵土壤生长显著低于本地土壤,导致植物的净光合速率显著低于非入侵土壤(P<0.05),在入侵地4种植物的光合作用受到显著抑制,所合成的碳水化合物显著低于非入侵土壤,因此4种本地植物在入侵地所积累的生物量显著低于非入侵土壤,减少了本地植株的株数、株高、生物量,最终影响本地植物群落的丰富度和生物多样性。但黄顶菊入侵对本地植物的影响存在物种差异。本研究丰富了黄顶菊入侵对土著植物的影响机制,为理解入侵种对群落结构影响和实现入侵生境恢复提供理论依据。本研究为同质园试验,与野外试验的条件不完全相同,因此要深入研究黄顶菊的入侵机制,还需要做长期的野外监测研究。
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
中国人兽共患病学报(2020年11期)2020-12-08
河南科学(2020年3期)2020-06-02
科学大众(中学)(2019年3期)2019-05-17
电子制作(2019年24期)2019-02-23
西南交通大学学报(2018年5期)2018-11-08
汽车观察(2018年10期)2018-11-06
科技知识动漫(2017年1期)2017-02-06
