
崂山腺齿越橘居群表型多样性研究
2019-10-08 05:44张宁宁王一马燕
山东农业科学 2019年8期
张宁宁 王一 马燕
摘要:本研究以崂山野生腺齿越橘6个自然居群的27个样本为材料,选取22个表型性状进行综合分析,研究居群间和居群内的表型变异程度及规律。结果表明:6个居群表型性状的变异程度不同,平均变异系数在9%~36%之间,株高和果穗长性状的变异最明显,居群间的变异水平大于居群内;生殖器官性状间具有明显相关性,但与营养器官性状不相关;主成分分析共得到6个主成分,第一主成分的占比最大,为42.878%;表型性状在各居群间的分化不明显,光照在一定程度上影响了性状的表达。
关键词:腺齿越橘;居群;表型多样性
中图分类号:S663.901 文献标识号:A 文章编号:1001-4942(2019)08-0016-05
Abstract Using 27 samples from 6 natural populations of wild Vaccinium oldhamii Miq. in Laoshan as materials, twenty two phenotypic traits were selected for variance analysis, principal component analysis and correlation analysis to study the variation degree and regulation among and within populations. The results showed that the phenotypic traits of 6 populations had different variation degrees, and the average variation coefficients were between 9%~36%. The variations of plant height and ear length were more obvious and variation levels between populations were greater than that within populations. There was significant correlation between reproductive organ traits, but had no correlation with vegetative organ traits. Six principal components were obtained by principal component analysis, and the proportion of first principal component was maximum as 42.878%. The phenotypic traits were not clearly differentiated among populations, and light affected trait expression to a certain extent.
Keywords Vaccinium oldhamii Miq.; Population; Phenotypic diversity
腺齿越橘(Vaccinium oldhamii Miq.)属杜鹃花科越橘属,主要分布在我国山东青岛崂山、烟台昆嵛山,2015年被列为中国稀有树种[1]。腺齿越橘与蓝莓同属越橘属,果实中含有能缓解眼部疲劳及抗癌的花青素、黄酮醇、酚酸等物质,可直接食用或用于食品药品加工[2,3]。腺齿越橘果实个头大、结果率高,花青素含量高,是美国蓝莓的1.9倍[4],其花、叶、果都具有一定的观赏性,在园林应用上有良好前景,拥有巨大的经济开发价值。关于越橘属生理生态[5]、繁殖技术[6]、品种分类[7]等方面的研究已见报道,但关于腺齿越橘表型和遗传多样性的研究较为少见。
环境因素在居群进化过程中起着非常重要的作用,它通过选择适应环境的个体来改变和控制居群中的基因型频率[8]。腺齿越橘的表型变异就是对崂山生态环境长期适应的结果。形态标记是指肉眼可见的或仪器可测量的植物外部特征,包括株高、花色、叶形、果实大小等表型性状,广义上还包括抗虫性等。作为最早开始被使用的遗传标记方法,它具有简单直接且易于观察记录的优点,在早期经典遗传学研究中发挥重要作用。本研究以嶗山6个自然居群的27个腺齿越橘样品的叶形、果色等22个表型性状为研究对象,对各性状进行综合性分析,以期为腺齿越橘种质资源的保护和评价提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为腺齿越橘,分别取自6个天然居群(表1)。
1.2 试验方法
于2018年7月腺齿越橘果期进行调查,根据其生物学特性和主要性状的差异,选取遗传稳定、不易受环境影响的表型性状(包括11个质量性状和11个数量性状)作为标记对象。每居群依据其自然分布状况选取生长良好且间隔50 m以上的植株,在树冠中上部4个方位选取叶和成熟果进行相应性状观察和测量,共收集27株。
1.3 测定指标及方法
质量性状用同一标准观察,数量性状使用精确度为0.01的游标卡尺进行测量,参照文献[9]的方法进行取样、测量并记录相关表型性状,记录标准见表2。
1.4 数据分析
利用 Microsoft Excel 2010 计算各性状的平均值、标准差和变异系数,以表型变异系数作为居群内居群间各形态特征变异度的测度。用 SPSS 20.0进行各性状间的主成分分析和相关性分析。用UPGMA法进行样品间的聚类分析。
2 结果与分析
2.1 数量性状变异特征
各数量性状的变异分析结果见表3,6个居群表型性状的变异程度不同,平均变异系数在9%~36%之间。各性状平均变异系数由高到低依次为:果穗长>株高>顶生叶长>营养枝小叶宽>营养枝小叶长、顶生叶宽>生花枝小叶宽、果纵径>果横径>果横纵径比>生花枝小叶长。其中果穗长平均变异系数为36%,生花枝小叶长为9%。
同一种群内各性状的变异程度各不相同,同一生态环境会对不同性状产生不同程度的影响。蔚竹庵、潮音瀑居群各性状变异程度较大,变化范围分别为4%~ 59%、3%~58%,果穗长变异系数均为最大;滑溜口居群的变异幅度最小,在9%~28%之间,果穗长变异系数最大,顶生叶长与营养枝小叶长变异系数较小,均为9%(表3)。本研究中6个居群的性状变异幅度由高到低为蔚竹庵、潮音瀑、华严寺、栈道、太清宫、滑溜口。
同一性状在不同居群内的变异系数相差较大。变异幅度最大的性状是株高,变幅在4%~52%之间,该性状在华岩寺居群中变异系数最大,潮音瀑居群中最小;顶生叶宽的变幅次之,在3%~49%之间,栈华严寺居群的该性状变异系数为49%,太清宫居群为3%;果纵径的变幅最小,在不同居群的变异系数仅相差8%(表3)。说明株高和顶生叶宽这类性状在生长过程中受环境影响较大。
各数量性状在物种水平上的变异系数在16%~56%之间,与各居群平均变异系数(9%~36%)相比,变幅更大。在居群内变异最大的果穗长性状,在居群间的变异系数为33%,而居群内变异最小的果横径(2%),在物种水平上有25%(表3)。居群间株高、果纵径等性状也高于居群内水平。
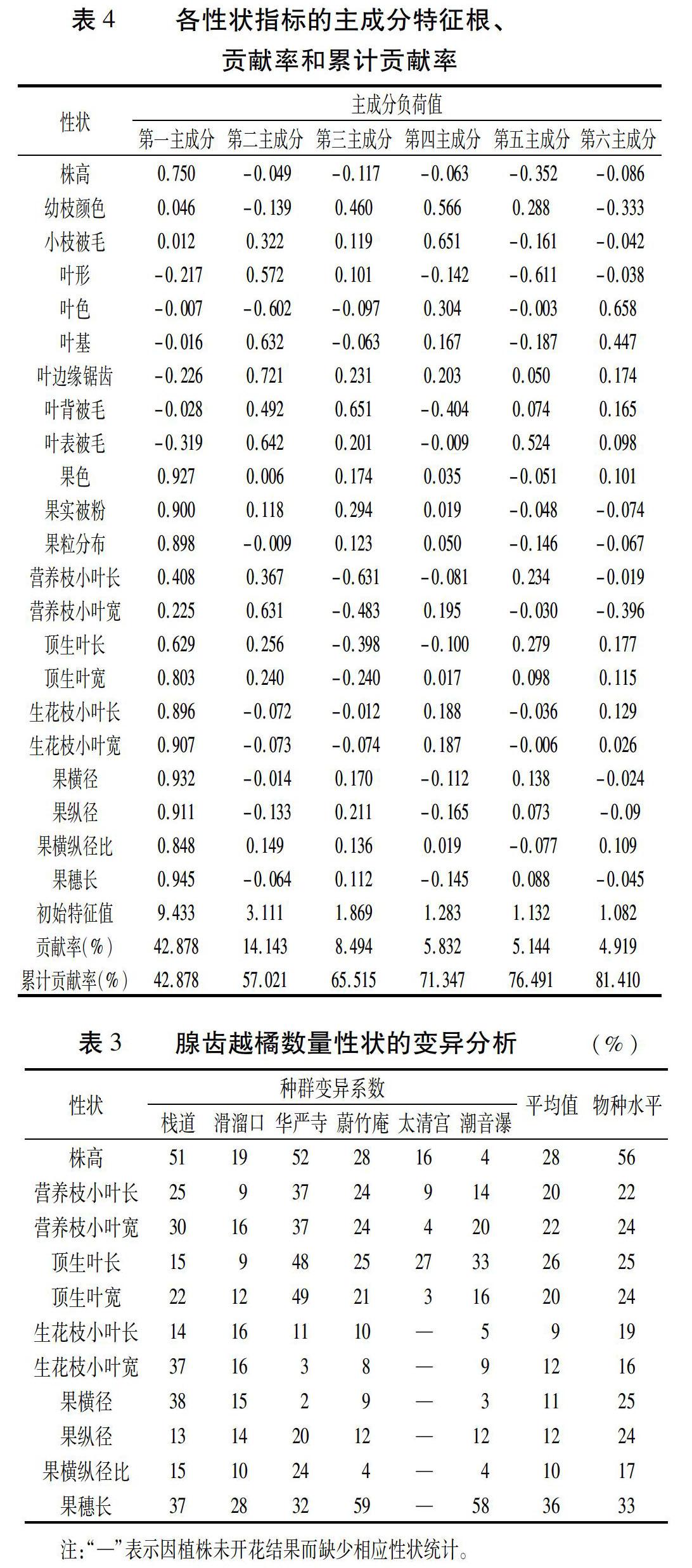
2.2 主成分分析
各性状指标的主成分分析结果如表4所示:共获得6个主成分,累计贡献率达到81.410%。其中第一主成分的贡献率最大,为42.878%,特征根值较大的性状有12个,反映了植株高度、果实生长状况以及顶生叶等情况,其中生花枝小叶长(0.896)、生花枝小叶宽(0.907)、果实被粉(0.900)、果粒分布(0.898)、果色(0.927)、果横径(0.932)、果纵径(0.911)、果横纵径比(0.848)及果穗长(0.945)起决定性作用;第二主成分贡献率为14.143%,特征向量较大性状反映叶形、叶色、叶表被毛等;第三主成分的贡献率为8.494%,主要反映营养枝的生长状况、叶背被毛;第四主成分的贡献率为5.832%,特征向量大于0.5的则是幼枝颜色及小枝被毛性状;第五、第六主成分的贡献率分别为5.144%、4.919%,反映了叶形、叶表被毛及叶色等情况。
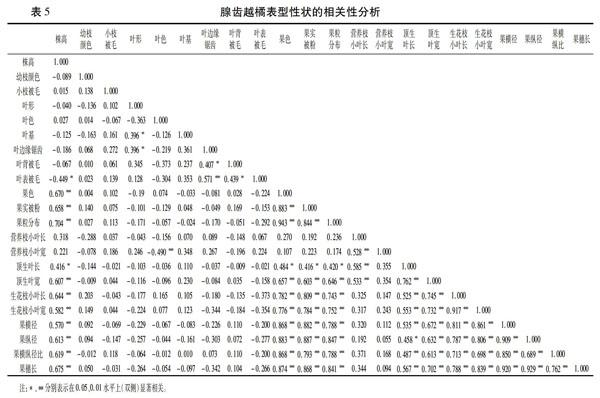
2.3 相关性分析
采用Pearson积矩相关系数分析22个性状之间的相关性,有76对性状的相关性达到显著水平。由表5可以看出,果色、果实被粉、果粒分布、顶生叶宽、生花枝小叶长、生花枝小叶宽、果横径、果纵径、果横纵径比、果穗长均与株高呈极显著正相关,顶生叶长与株高呈显著正相关,叶表被毛情况与株高呈显著负相关;叶形分别与叶基、叶边缘锯齿显著正相关,相关系数均为0.396;叶边缘锯齿、叶表被毛、叶背被毛之间两两相关,其中叶边缘锯齿与叶表被毛的相关性达极显著水平;营养枝小叶长与营养枝小叶宽、顶生叶长与顶生叶宽、生花枝小叶长与生花枝小叶宽之间呈极显著正相关,相关系数分别为为0.528、0.762、0.917,说明腺齿越橘各部位的叶片长宽之间均有相关性,叶形较稳定;果色、果实被粉、果粒分布、生花枝小叶长、生花枝小叶宽、果横径、果纵径、果横纵比、果穗长这些性状相互间均呈显著正相关,说明腺齿越橘生殖性状之间具有同步性,而顶生叶长、顶生叶宽与这些性状之间也呈显著正相关,说明长在有生花枝的营养枝上的顶生叶更大。幼枝颜色、小枝被毛情况、叶色、叶基、叶边缘锯齿之间无明显相关性,说明这些性状之间没有规律,在研究腺齿越橘表型性状时可舍弃,对研究遗传变异关系无意义。
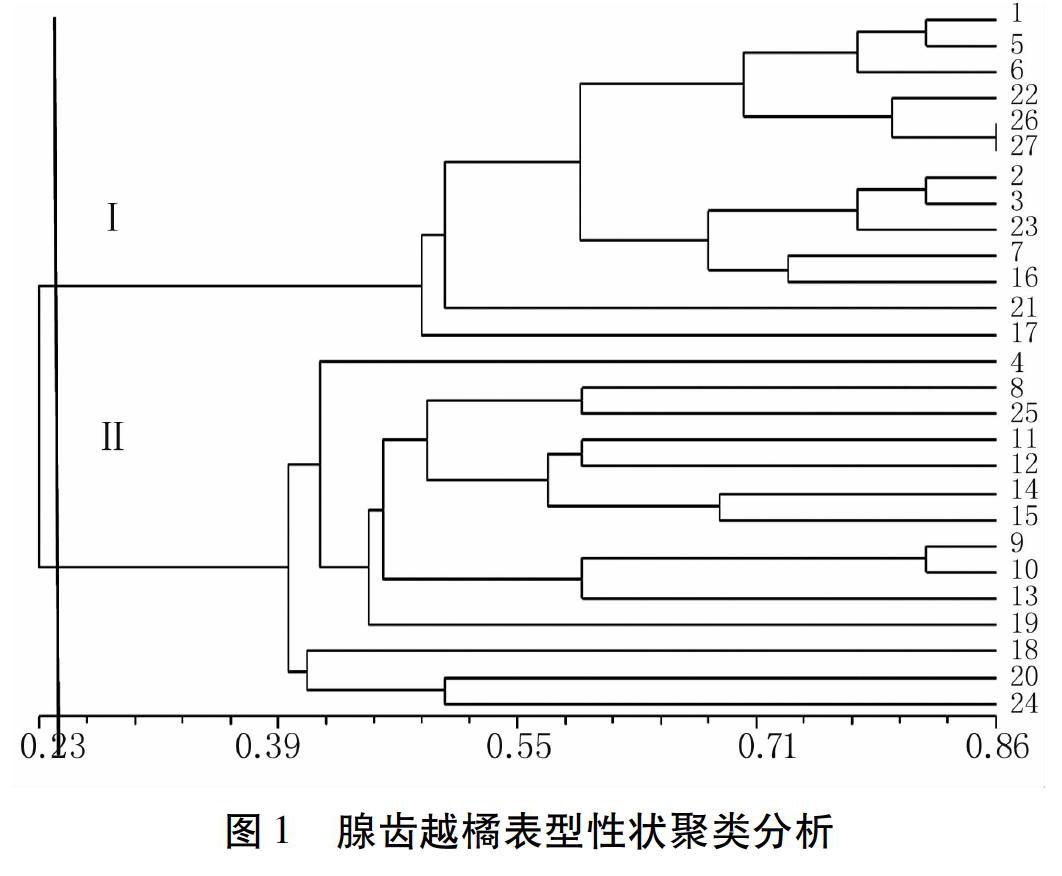
2.4 聚类分析
由图1可以看出,在0.23水平上腺齿越橘可分为两大类。来自滑溜口居群的编号8~15的样品聚为一类,来自潮音瀑居群的18、20样品聚为一类,来自太清宫居群的26、27样品聚为一类,而来自栈道居群的1~7样品没有聚为一类,来自华严寺居群的16、17、19样品没有聚为一类,来自蔚竹庵居群的20~25样品也未能聚为一类。说明崂山地区腺齿越橘的性状在各居群间的分化现象不明显。
3 讨论与结论
表型是物种进化和变异的表现,一般受到基因与生态气候环境共同作用[10],能最直观地反映物种的遗传多样性,而腺齿越橘各居群的生长环境差异正是种内进化过程中对环境适应的结果。本研究发现6个腺齿越橘居群的各性状均存在不同程度的差异,而同一性状在各居群间的差异程度也不相同,说明地理环境在一定程度上影响了表型性状的表达。果穗长能够反映结实率的变化,结实率往往受到植株生长情况、海拔、光照和人为干扰等因素的影響[11],因而海拔较低的蔚竹庵居群、栈道居群和潮音瀑居群的结实率变化较大,而滑溜口居群的生态环境受人为干扰程度较小,成年植株的结实率较好。腺齿越橘同其它越橘属植物一样,喜光耐荫,在林分郁闭度较大的环境也能生长,但在林分郁闭度低的环境中结实率高[12],这可能是太清宫居群没有结果的植株,而滑溜口居群结实率高,生长均匀,果穗长变异较小的原因。
通过主成分和相关性分析可知,相关性水平达到显著的性状有76对,生殖器官性状在腺齿越橘性状中占较重要地位。生殖器官性状如生花枝小叶长、生花枝小叶宽、果实被粉、果粒分布、果色、果横径、果纵径、果横纵比及果穗长之间均具有极显著相关性。株高与这些生殖器官性状之间显著相关,但与营养器官性状并无明显相关。营养器官性状与生殖器官性状之间也无明显相关,说明在腺齿越橘生长过程中,营养器官对生殖器官的影响不大,这与代英超[13]对大别山山核桃的研究结果一致。
经过聚类分析发现,来自不同居群的样本相似性系数更高,而来自同一居群的样品并不能聚为一类,居群之间相互夹杂,说明腺齿越橘性状在居群间分化并不明显,遗传变异主要在居群内。根据实地调查结果结合聚类分析得到,生长在阳坡或半阳坡的4、8、9、10、11、12、13、14、15、18、19、20、24、25聚为Ⅰ类,生长在阴坡或半阴坡的1、2、3、5、6、7、16、21、22、23、26、27聚为Ⅱ类,说明环境在一定程度上影响了腺齿越橘性状的表达,这也是腺齿越橘适应环境的表现。
对崂山野生腺齿越橘的表型多样性研究表明,不同样本的22个性状的变异程度存在差异,各性状间的相关性不等,聚类分析结果表明相同环境下生长的腺齿越橘聚为一类,说明生长环境对腺齿越橘的外部形态存在较明显的影响,研究结果可为今后越橘属栽培种蓝莓种质资源的育种选择提供参考价值。
参 考 文 献:
[1] 李文清,臧德奎,解孝满. 山东珍稀濒危保护树种[M]. 北京:科学出版社,2015: 55.
[2] Elisabetta B, Flavia G, Paolo F, et al. Nutritional profile and productivity of bilberry (Vaccinium myrtillus L.) in different habitats of a protected area of the Eastern Italian Alps[J].Food Science, 2013, 78(5): 673-678.
[3] Giovanelli G, Buratti S. Comparison of polyphenolic composition and antioxidant activity of wild Italian blueberries and some cultivated varieties[J]. Food Chem., 2009, 112(4): 903-908.
[4] Song J H, Cheong E J, Kim H S, et al. Variation of morphological characteristics and anthocyanin contents from fruit of Vaccinium oldhamii in Korea[J]. Journal of Korean Forest Society, 2015, 104(2): 193-197.
[5] 李彬彬,楊俊枫,高源,等. 越橘叶片秋季变色期间花青苷和叶绿素的变化特性[J]. 园艺学报,2017,44(12):2361-2371.
[6] 刘有春,杨艳敏,王莉,等. 越橘种子破除休眠、萌发及快速成苗方案优化[J]. 果树学报,2018,35(9): 1147-1154.
[7] Silva S D A, Antunes L E C, Anthonisen D G, et al. Characterization of blueberry genotypes using molecular markers[J]. Revista Brasileira De Fruticultura, 2008, 30(1): 180-184.
[8] 汪小凡,廖万金,宋志平. 小毛茛居群的遗传分化及其与空间隔离的相关性[J]. 生物多样性,2001, 9(2): 138-144.
[9] 童冉,姜丽娜,吴小龙,等. 野生玫瑰种质资源鉴定评价技术规范的研制[J].山东林业科技,2016(6):20-24.
[10] 马育华,盖钧镒. 江淮下游地区大豆地方品种的初步研究(三)数量性状的表型、遗传型相关,选择指数,及其育种意义[J]. 作物学报,1979,5(4): 1-11.
[11] 曹焱,王勇,翁海龙,等. 基于环境因子的笃斯越橘结实规律研究[J].安徽农业科学, 2015,43(16): 162-163, 259.
[12] 崔红. 笃斯越桔的经济价值及其集约经营措施[J]. 特种经济动植物,2007(7): 46-47.
[13] 代英超. 基于表型变异和ISSR标记的大别山山核桃遗传多样性研究[D]. 杭州:浙江农林大学,2015.
