
干旱胁迫对鼓节竹光合特性和叶绿素荧光参数的影响
2019-10-09 01:46赖金莉郑俊鸣何天友陈凌艳荣俊冬郑郁善
福建农林大学学报(自然科学版) 2019年5期
赖金莉,张 力,薛 磊,郑俊鸣,何天友,陈凌艳,荣俊冬,郑郁善,
(1.福建农林大学园林学院;2.福建农林大学林学院,福建福州350002)
鼓节竹(Bambusa tuldoides‘Swolleninternode’)为簕竹属青竿竹的一个栽培变种,与原栽培型的主要区别为竿的下部节间极为短缩,且基部肿胀,鼓节竹竿形的独特性使其具有很高的观赏价值,适宜小规模片植、与山石配景及制作盆景观赏[1-2],在南方城市的园林美化和绿化上具有很好的发展前景.
我国竹类资源丰富,但大部分分布于长江流域以南,北方引种区普遍存在着季节性干旱期,对竹类的生长和效益有很大的影响[3].干旱胁迫是影响植物生长和发育的重要因素之一,其中以叶片的光合作用对干旱的响应最为敏感[4].干旱胁迫影响植物光合作用中光能电子传递和转换、光合磷酸化及暗反应等一系列过程[5],严重的干旱胁迫会降低植物的光合机构活性,使PSⅡ光系统受到破坏和损伤[6].叶绿素荧光技术是研究植物光合生理及逆境胁迫下生理变化的内在探针,它在植物光合作用过程中对光能的吸收、传递、耗散和分配等方面有独特的作用,与“表观性”的气体交换参数相比,叶绿素荧光参数具有反映植物“内在性”的特点[5,7].目前关于鼓节竹的研究很少,相关的研究主要集中在抗寒性[8-10]、滞尘效应[11]、防风固沙[12]及根际土壤酶活性[13]等方面,仅有龙智慧[14]对其在自然干旱胁迫下的形态和生理生化指标进行了研究,而有关干旱胁迫下鼓节竹的光合特性和叶绿素荧光参数的研究尚未见报道.因此,本试验以一年生鼓节竹分株苗为材料,研究了4个不同梯度的土壤含水量模拟的干旱胁迫处理下鼓节竹叶片的光合特性和叶绿素荧光参数的变化,旨在了解鼓节竹对不同干旱胁迫程度的适应性,为鼓节竹的栽培管理及耐旱性品种的选育提供参考.
1 材料与方法
1.1 材料
供试材料为一年生鼓节竹分株苗,均取自福建省福州市南屿镇的福州青芳竹种园.该竹种园属于典型的海洋性亚热带季风气候,雨量充沛,年均降水量900~2100 mm,年平均气温16~20℃,无霜期326 d,年均日照时间1700~1980 h,土壤类型主要为红壤,土壤肥力较好,适合鼓节竹的生长[15-16].2017年11月将带土球的鼓节竹分株苗移栽到生长盆(直径55 cm、深35 cm)中生长,盆内基质为泥炭土与红壤(体积比为1∶2)的混合土,每盆装混合土30 kg.在试验前用多菌灵的1000倍水溶液对土壤进行消毒,并在阳光下暴晒2 d以达到更彻底的消毒杀菌的目的,自然风干后过筛装盆.试验苗种植在福建农林大学园林学院温室大棚内,在干旱胁迫处理前按常规大棚盆栽苗水分管理的要求浇水,并定期进行除虫、除草、施肥等措施,盆栽苗正常生长5个月后,对其进行土壤含水量的控制.
1.2 方法
1.2.1 试验设计 根据应叶青等[17]的方法(略有改动)设置如下4个处理:对照组(CK)(土壤含水量为最大田间持水量的75%~80%)、轻度干旱胁迫组(土壤含水量为最大田间持水量的55%~60%)、中度干旱胁迫组(土壤含水量为最大田间持水量的40%~45%)、重度干旱胁迫组(土壤含水量为最大田间持水量的20%~25%),每个处理3盆苗,每盆5株,计12盆(1盆苗1个重复,每个处理3盆3个重复).试验处理前5 d对试验苗停止浇水,开始进行干旱胁迫,处理时根据土壤重量和含水量补充水分使土壤含水量保持在设定的范围内.土壤含水量的控制方法为:试验期间于每天下午18:00对盆栽进行称重并用TDR 350便携式土壤水分速测仪(美国Spectrum公司)测定土壤含水量,根据测定结果及土壤重量向盆内补充水分,使土壤含水量在控制范围内.为了减少蒸发,在盆口套塑料袋防止水分蒸发.采用环刀法[18]测定最大田间持水量,通过公式计算出土壤最大田间持水量为44%.因此,CK,轻度、中度、重度干旱胁迫组的土壤含水量分别为 33.0%~35.2%、24.2%~26.4%、17.6%~19.8%、8.8%~11.0%.试验期间进行不定期的除草、除笋.分别于试验当天(0 d)及胁迫后的第7、14、21、28、35、42、49、56天测定鼓节竹光合特性和叶绿素荧光参数等各项指标.
1.2.2 外部形态的观察 在干旱胁迫处理期间,于每次测量的当天上午观察植株的叶片形态特征.
1.2.3 气体交换参数的测定 采用Li-6400XT便携式光合作用系统(美国LI-COR公司)测定气体交换参数.设定光合有效辐射为1000 μmol·m-2·s-1,气体流速为(500±0.5) mmol·s-1,外接 CO2缓冲瓶控制大气 CO2浓度为(400±0.5) μmol·mol-1,并于每个时间节点的上午 8:30—11:30 测定叶片的净光合速率(net photosynthetie rate, Pn)、蒸腾速率(transpiration rate, Tr)、气孔导度(stomatal conductance, Gs)、胞间CO2浓度(intercellular CO2concentration,Ci).测定的样叶为植株顶部往下第3~5片成熟的功能叶片,每个处理3个重复,每个重复待数据稳定后读取5个数据,计算水分利用效率(water use efficiency,WUE)、气孔限制值(stomatal limit value, Ls).WUE=Pn/Tr,Ls=1-Ci/Ca(Ca为大气 CO2浓度)[19].
1.2.4 叶绿素荧光参数的测定 采用OPTI-Sciences OS5P便携式脉冲调制叶绿素荧光仪(北京澳作生态仪器有限公司)测定叶绿素荧光参数.选取每个重复植株中上部受光均匀的成熟叶3片,挂牌标记.每个叶片暗适应 25 min后先测定吸光系数,照射测量光(0.5 μmol·m-2·s-1),测定初始荧光产量(F0),再照射饱和脉冲光(2500 μmol·m-2·s-1),测定最大荧光产量(Fm),并计算可变荧光(Fv),即可计算 PSⅡ光系统的潜在活性(Fv/F0).用暗适应夹对叶片进行20 min暗适应后,选择Fv/Fm测量模式,测得PSⅡ光系统的最大光化学效率(Fv/Fm);选择Yield模式,测得PSⅡ实际光化学量产量(ΦPSⅡ);选择Kinetic模式,测出光合电子传递速率(apparent photosynthetic electron transport rate,ETR)、光化学猝灭系数(photochemical quenching, qP)、非光化学猝灭系数(non photochemical quenching, qN).
1.2.5 数据处理 用Excel 2003软件对数据进行整理及作图,采用SPSS 20软件进行数据分析,取3个重复的平均值±标准差.
2 结果与分析
2.1 干旱胁迫对鼓节竹叶片形态的影响
在干旱胁迫的过程中,鼓节竹的叶片发生了失水、皱缩、黄化、长斑及脱落等现象.其中,CK保持正常的生长,叶色浓绿;轻度干旱胁迫下的生长状况与CK相差不大,叶片在胁迫结束后依然保持绿色,只有少部分叶片失水干枯;中度干旱胁迫前期植株生长较为正常,叶色变化不明显,叶缘部分失水,部分叶片的背面出现斑点,胁迫42 d时叶片开始出现萎蔫干枯的现象,至胁迫结束时依然有2/3的叶片保持绿色;在重度干旱胁迫的过程中,胁迫35 d后植株生长开始受到抑制,叶片出现失水皱缩的现象,胁迫结束时有1/2的叶片枯萎脱落,其余生长状况不佳.
2.2 干旱胁迫对鼓节竹叶片光合特性的影响
由图1A~1C可知,鼓节竹叶片的Pn、Gs、Tr在干旱胁迫下都随着胁迫时间的增加及胁迫强度的增大呈下降的趋势,但下降幅度略有不同.方差分析显示,干旱胁迫能显著降低Pn、Gs、Tr.胁迫56 d时,与CK相比,轻度、中度、重度干旱胁迫下的 Pn分别下降了 73.26%、91.24%、99.00%(P<0.05),Gs分别下降了48.83%、68.45%、90.95%(P<0.05),Tr分别下降了 67.09%、77.45%、85.58%(P<0.05).由此表明,Pn对干旱胁迫的响应比Gs、Tr敏感.
由图1D可知,鼓节竹叶片的Ci在干旱胁迫下随着胁迫时间的增加呈先下降后上升的趋势,分别在轻度干旱胁迫21 d、中度干旱胁迫14 d、重度干旱胁迫7 d时达到最小值,干旱胁迫56 d时的Ci大小为:重度干旱胁迫>中度干旱胁迫>轻度干旱胁迫>CK(P<0.05).由图1E可知,与CK相比,鼓节竹叶片的WUE在轻度、中度干旱胁迫下随着胁迫时间的增加呈先上升后下降的趋势,而在重度干旱胁迫下呈下降的趋势.由图1F可知,鼓节竹叶片的Ls随着干旱胁迫时间的增加呈先上升后下降的趋势,分别在轻度干旱胁迫21 d、中度干旱胁迫14 d、重度干旱胁迫7 d时达到最大值,此时,Ls与Ci出现了明显相反的趋势,表明在轻度干旱胁迫21 d、中度干旱胁迫14 d、重度干旱胁迫7 d时光合作用由气孔限制因素变为非气孔限制因素.方差分析显示,干旱胁迫对Ci的影响在胁迫21 d时开始显著,对WUE的影响在胁迫35 d时开始显著,对Ls的影响在胁迫14 d时开始显著.
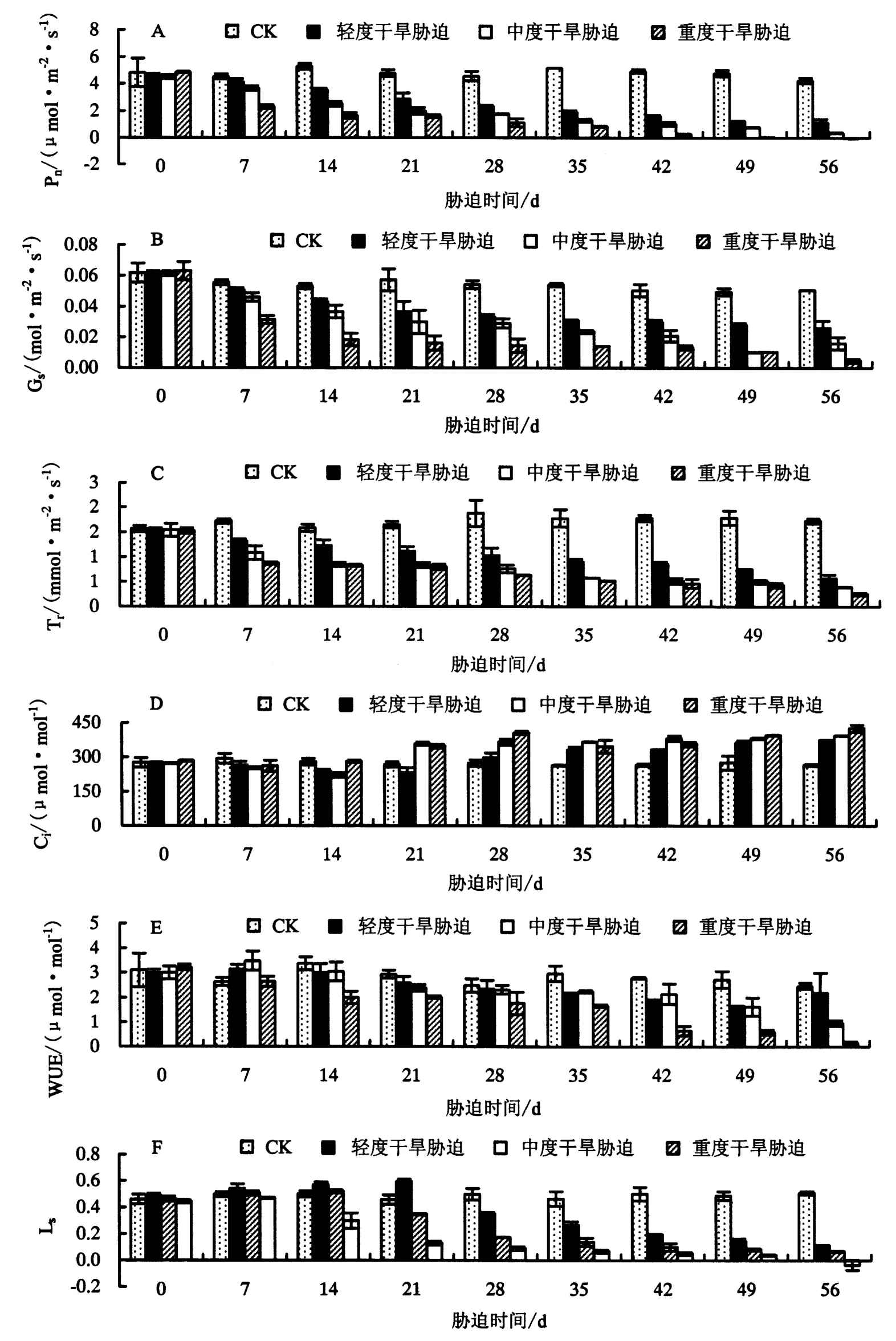
图1 干旱胁迫对鼓节竹叶片 Pn、Gs、Ci、Tr、WUE、Ls的影响Fig.1 Effect of drought stress on Pn, Gs, Ci, Tr, WUE, Lsof B.tuldoides ‘Swolleninternode’
2.3 干旱胁迫对鼓节竹叶片叶绿素荧光参数的影响
2.3.1 对 F0、Fm、Fv/Fm、Fv/F0的影响 由图2A 可知,不同程度的干旱胁迫对鼓节竹叶片 F0的影响程度是不同的.与CK相比,轻度、中度干旱胁迫仅在56 d时对F0的影响显著(P<0.05),在前期对其影响不明显(P>0.05);而在重度干旱胁迫的前28 d,F0均呈上升的趋势,在28 d时达到最大值后逐渐下降,且在胁迫 49、56 d 时分别下降22.06%、36.59%(P<0.05).由此可知,F0对重度干旱胁迫敏感,轻度、中度干旱胁迫对其影响不大.方差分析显示,不同干旱胁迫处理之间的F0差异不显著,可推测干旱胁迫对鼓节竹叶片PSⅡ光系统产生了一定的损害.
由图2B可知,随着干旱胁迫时间的增加及胁迫强度的增大,鼓节竹叶片的Fm呈下降的趋势.与CK相比,Fm在轻度、中度干旱胁迫28 d时开始显著下降(P<0.05),在重度干旱胁迫21 d时开始显著下降(P<0.05).方差分析显示,干旱胁迫28 d后不同胁迫处理之间的Fm差异显著,且随着胁迫时间的增加,Fm下降幅度增大,说明长期的干旱胁迫能显著降低鼓节竹叶片的Fm.
由图2C可知,鼓节竹叶片的Fv/Fm随着胁迫时间的增加及胁迫强度的增大呈下降的趋势.与CK相比,Fv/Fm在轻度、中度干旱胁迫14 d时开始显著下降(P<0.05),在重度干旱胁迫7 d时开始显著下降(P<0.05);胁迫 56 d 时,Fv/Fm在轻度、中度、重度干旱胁迫下分别下降了 34.65%、36.55%、48.66%(P<0.05).方差分析显示,干旱胁迫能显著降低鼓节竹叶片的Fv/Fm,即干旱胁迫可显著降低其PSⅡ光能转化效率.
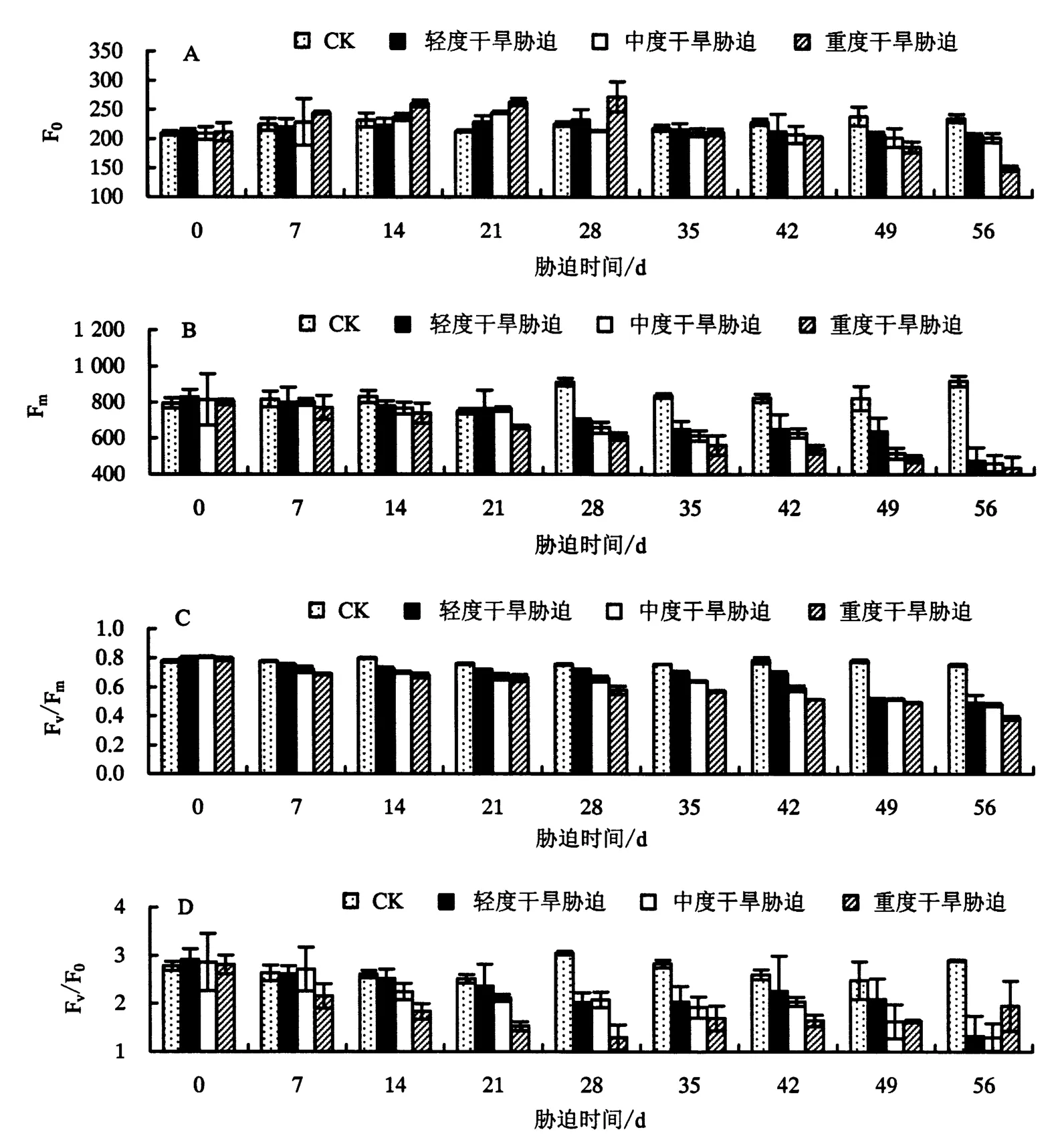
图2 干旱胁迫对鼓节竹叶片 F0、Fm、Fv/Fm、Fv/F0的影响Fig.2 Effect of drought stress on F0, Fm, Fv/Fmand Fv/F0of B.tuldoides ‘Swolleninternode’
由图2D可知:与CK相比,在轻度干旱胁迫下,鼓节竹叶片的Fv/F0随着胁迫时间的增加呈下降的趋势,但下降不显著(P>0.05);在中度干旱胁迫下,前21 d的Fv/F0随着胁迫时间的增加呈下降的趋势,在胁迫 21 d 时开始显著下降(P<0.05);在重度干旱胁迫下,Fv/F0在前 14 d 下降不显著(P>0.05),14~42 d显著下降(P<0.05),之后下降不显著(P>0.05),与胁迫49 d相比,胁迫56 d甚至有上升的趋势.方差分析显示,干旱胁迫14 d时,鼓节竹叶片的Fv/F0开始显著下降,即干旱胁迫14 d后其PSⅡ潜在活性才开始受到显著影响.
2.3.2 对ΦPSⅡ、ETR、qP、qN 的影响 由图3A 可知,在干旱胁迫下,鼓节竹叶片的 ΦPSⅡ随着胁迫时间的增加逐渐下降.方差分析显示,干旱胁迫14 d时,鼓节竹叶片的ΦPSⅡ开始显著下降,即干旱胁迫14 d后其对原初光能的捕获效率才开始受到显著影响.
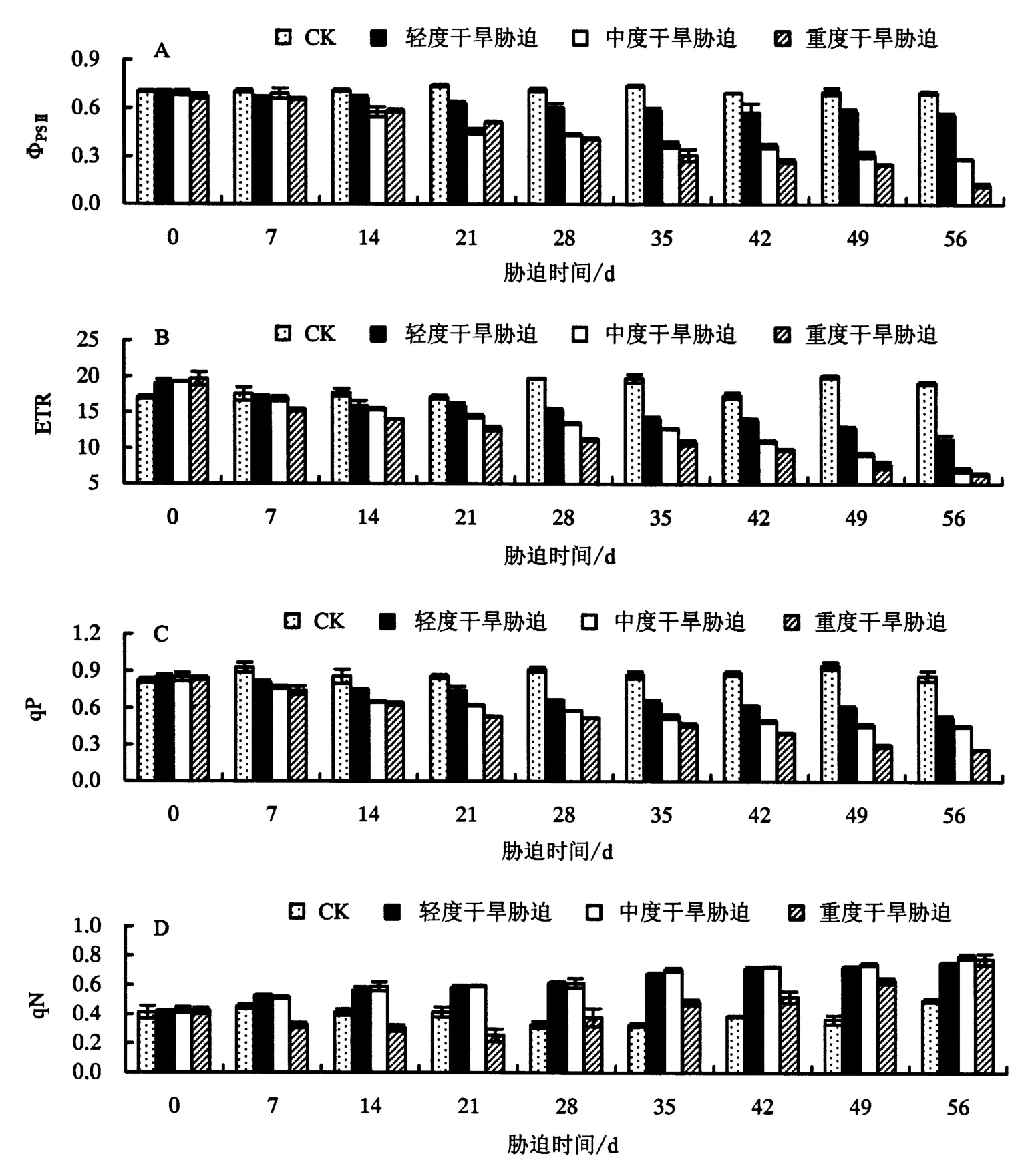
图3 干旱胁迫对鼓节竹叶片ΦPSⅡ、ETR、qP、qN的影响Fig.3 Effect of drought stress on ΦPSⅡ, ETR, qP and qN of B.tuldoides ‘Swolleninternode’
由图3B可知,鼓节竹叶片的ETR随着干旱胁迫时间的增加及胁迫强度的增大呈下降的趋势.与CK相比,在轻度干旱胁迫下,ETR在胁迫28 d时开始显著下降(P<0.05);在中度、重度干旱胁迫下,ETR在胁迫14 d时开始显著下降(P<0.05),且在中度胁迫 21 d、重度胁迫 28 d时开始极显著下降(P<0.01).方差分析显示,干旱胁迫可以显著降低鼓节竹叶片的ETR,且随着胁迫时间的增加,其下降幅度增大.
由图3C可知,鼓节竹叶片的qP随着干旱胁迫时间的增加及胁迫强度的增大呈下降的趋势.与CK相比,不同程度的干旱胁迫均使qP显著下降(P<0.05),其中,中度、重度干旱胁迫在胁迫后期对其影响较大.方差分析显示,干旱胁迫能显著降低鼓节竹叶片的qP,且随着胁迫时间的增加影响越大.
由图3D可知:CK的qN在生长过程中基本稳定,变化不大;随着胁迫时间的增加,qN在轻度、中度干旱胁迫下呈上升的趋势(P<0.05),而在重度干旱胁迫下呈先下降后上升的趋势(P<0.05).方差分析显示,干旱胁迫能显著增加鼓节竹叶片的qN.
3 讨论
干旱胁迫下植物叶片会发生一些列的变化.赵兰等[20]对4种观赏地被竹的研究发现,在胁迫后期,耐旱性最强的黄条金刚竹的叶片依然保持绿色,耐旱性次之的菲白竹少数叶片卷曲发黄,耐旱性最差的铺地竹多数植株干枯死亡.干旱胁迫下,植物叶片发生失水萎蔫的原因:一是胁迫降低叶片组织的含水量,从而影响正常的生理代谢,严重者甚至死亡[21];二是遭受干旱胁迫时,植物将构建更发达的根系吸收土壤中的水分,此时为了维持生长,叶片自上而下将营养物质运输到根系,提高根冠比来提高耐旱性[22].本试验中,鼓节竹叶片在轻度干旱胁迫下除了少数失水干枯外,其余均保持绿色,中度干旱胁迫下有2/3的叶片保持绿色,重度干旱胁迫下有1/2的叶片干枯脱落,其余生长不佳.因此从外部形态上分析,鼓节竹在轻度、中度干旱胁迫下依然保持有较多的绿色叶片,从而表现出较强的抗旱性,而在重度干旱胁迫下生长不佳,表现出对持续重度干旱胁迫的不耐受性.
光合作用与植物生长发育关系密切,被认为是自然环境中限制植物生长的最重要的因子之一.研究表明,光合作用对干旱胁迫很敏感,当干旱胁迫发生时,植物叶片的Pn下降,这种下降受气孔和非气孔因素的影响[23].本试验中,根据Farquhar et al[24]的理论,在干旱胁迫前期,鼓节竹叶片的Pn与Ci同时下降,且Ls增大,说明在胁迫前期Pn下降是由气孔因素导致的;而在胁迫后期,Pn下降的同时Ci反而上升,且Ls减小,说明在胁迫后期Pn下降是由自身叶肉细胞光合能力下降导致的,此结果与沈少炎[25]对花吊丝竹的研究结果相一致.一般来说,植物叶片的Gs与Tr之间存在正相关关系,干旱胁迫下随着Gs的下降,Tr也会下降.本试验中,鼓节竹叶片Tr的下降幅度大于Gs,原因可能是此时除了气孔阻力影响Tr外,还受到外界水蒸气浓度梯度的影响,随着土壤含水量的降低,干旱胁迫程度的增大,两者交换水蒸气的浓度梯度也逐渐减小,从而导致蒸腾作用比Gs的下降幅度大.WUE可表征植物的耐旱能力和适应能力[26].鼓节竹叶片的WUE在轻度、中度干旱胁迫下随着胁迫时间的增加呈先上升后下降的趋势,在重度干旱胁迫下则一直下降,说明鼓节竹在轻度、中度干旱胁迫前期(胁迫21 d前)可通过提高WUE来适应干旱的环境;在轻度、中度干旱胁迫的后期(胁迫21 d后)及重度干旱胁迫下,WUE下降,可能是由于长时间的干旱胁迫及严重的干旱胁迫使鼓节竹的光合同化能力受到抑制,降低了其生产能力.这与毛竹幼苗在干旱胁迫下的研究结果[17]类似.
叶绿素荧光动力学作为一种非破坏性的评价方法,可以在干旱胁迫的环境下准确、快速地了解植物的健康状况,反映植物对干旱胁迫的响应.叶绿素荧光参数与光合作用关系密切,任何对光合作用有影响的环境因子都可以通过叶绿素荧光动力学参数体现出来[27];此外,还可根据荧光参数的变化来判断植物光系统受损害的严重程度[28-29].本试验结果显示:鼓节竹叶片的F0在干旱胁迫下随着胁迫时间的增加呈先上升后下降的趋势,出现这种现象的原因可能是,植株在胁迫初期可以启动相应的恢复机制来抵御干旱胁迫,但随着胁迫时间的增加,这种机制逐渐遭到破坏,从而使F0下降,由此说明鼓节竹能抵御一定时间的干旱胁迫,这一现象也出现在杉木[30]上;Fm在不同程度的干旱胁迫下随着胁迫时间的增加均呈下降的趋势,说明在干旱的环境中,植株PSⅡ反应中心的活性及电子传递受到了抑制,导致其生长受到了一定程度的伤害.Fv/Fm可监测和揭示植物在遭受胁迫后PSⅡ反应中心的光能转化效率[31],非胁迫条件下不受物种和生长环境的影响,一般为0.80~0.85,而在植物遭受胁迫后明显下降[32].Fv/F0则反映了PSⅡ的潜在活性,Fv/Fm、Fv/F0是表明光化学反应状况的两个重要参数[33].鼓节竹叶片的Fv/Fm在干旱胁迫下随着胁迫时间的增加呈下降的趋势,在胁迫56 d后,轻度、中度、重度干旱胁迫下的Fv/Fm较CK分别下降到0.493、0.479、0.387,明显低于正常范围值(0.80~0.85);Fv/F0在轻度、中度干旱胁迫下随着胁迫时间的增加均呈下降的趋势,在重度干旱胁迫下呈先下降后上升的趋势.说明干旱胁迫对鼓节竹叶片的PSⅡ光化学系统有一定的伤害,抑制了PSⅡ反应中心的光能转化效率及潜在活性,但随着胁迫时间的增加,重度干旱胁迫下的Fv/F0有所回升,推测在重度胁迫后期PSⅡ反应中心可能启动了相应的防御机制来抵御干旱.因此,其潜在活性再一次被激发,其他植物,如胡杨、灰叶胡杨[34]也出现了类似的现象.
ΦPSⅡ反映了部分PSⅡ反应中心在关闭的情况下对原初光能的捕获效率;ETR则反映了PSⅡ反应中心的电子捕获效率[35].干旱胁迫降低了鼓节竹叶片的ΦPSⅡ、ETR,表明干旱胁迫降低了植株对原初光能的捕获效率及光合电子传递速率,进而影响其光合能力,这一结论与鼓节竹叶片Pn显著下降的结果相一致.qP反映了PSⅡ反应中心的开放程度及PSⅡ原初电子受体QA的还原状态,其值越高,说明PSⅡ反应中心的电子传递活性越大.qN是一种自我保护机制,反映了PSⅡ天线色素吸收了不能用于光合电子传递而以热能形式耗散掉的部分光能,从而对光合机构起到一定的保护作用[26,36].本试验中,干旱胁迫降低了鼓节竹叶片的qP,说明干旱胁迫降低了PSⅡ反应中心捕获能量转化为化学能的能力,导致PSⅡ的电子传递活性减小,从而降低了光合利用效率及对光能的利用,qP的降低,也反映出干旱胁迫降低了PSⅡ反应中心的开放程度.鼓节竹叶片的qN在轻度、中度干旱胁迫下呈上升的趋势,则说明植株在此时可通过增强热耗散能力以降低过多的光能对光合机构的损害,这是植物的自我保护机制;重度干旱胁迫下,由于胁迫程度较大,PSⅡ反应中心活性受到较大的损害,从而导致qN下降,之后随着胁迫时间的增加,植株已经对胁迫有了一定的适应性,由于qP也下降,从而导致激发能以qN的形式散发出去.qN增大体现的是植物的一种自我保护机制,这一现象与李娟等[19]对黄条金刚竹的研究结果相类似,其他植物,如胡杨[34]、赤霞珠葡萄[36]、决明属植物[37]、黑果枸杞[24]等也出现了qN增大的现象,由此说明这种保护机制可能出现在绝大部分植物身上.
4 结论
综合认为:在干旱胁迫下,鼓节竹可通过改变自身叶片的形态来抵御干旱胁迫带来的伤害,从而在轻度、中度干旱胁迫下表现出一定的抗旱性;干旱胁迫对鼓节竹的光合特性和叶绿素荧光参数的影响明显,气孔和非气孔因素都是影响鼓节竹Pn下降的因素;随着干旱胁迫时间的增加及胁迫强度的增大,鼓节竹可通过降低qP、ΦPSⅡ以及增加qN来降低PSⅡ反应中心的开放程度,降低PSⅡ反应中心在干旱胁迫下的伤害,这是鼓节竹在干旱胁迫下自我保护机制的体现,也是鼓节竹具有一定耐旱性的表现;维持鼓节竹具有较高的WUE的土壤含水量为17.6%~26.4%.
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06
电脑报(2020年12期)2020-06-30
中国果业信息(2019年1期)2019-01-05
森林工程(2018年4期)2018-08-04
时代农机(2018年11期)2018-03-17
生物学教学(2017年9期)2017-08-20
安徽医科大学学报(2016年12期)2017-01-15
癌症进展(2016年9期)2016-08-22
电源技术(2016年9期)2016-02-27
食品工业科技(2014年6期)2014-05-10
