
杉木全双列杂交试验数据分析方法比较
2019-10-09 01:46何贵平程亚平肖纪军王海蓉邱勇斌
福建农林大学学报(自然科学版) 2019年5期
齐 明, 何贵平, 程亚平, 肖纪军, 王海蓉, 邱勇斌
(1.中国林业科学研究院亚热带林业研究所,浙江富阳311400;2.浙江省林木育种技术研究重点实验室,浙江 富阳311400;3.浙江省庆元县庆元林场,浙江庆元323805;4.浙江省龙泉市林科院,浙江龙泉323700;5.浙江省遂昌县林业技术推广总站,浙江 遂昌323300;6.浙江省开化县林场,浙江 开化324300)
林木高世代育种方案的制订,需要一套包括反交效应在内的全双列杂交交配设计结果,而研究GriffingⅢ或Ⅰ模式非平衡资料的处理方法具有重要意义.一些学者[1-4]对双列杂交设计和配合力分析方法的模型进行了研制,并发表针对平衡试验数据的分析模型.Griffing[5]系统地提出了双列杂交设计的4种模式.1977年,Keuls et al[6,7]提出了平衡不平衡、规则不规则的GriffingⅢ模式的矩阵分析理论.1997年朱军[8]建议采用MINQUE(1)来处理GriffingⅢ模式的试验数据,极大地提高了试验效率,获得丰富的遗传信息,但仍未解决负的方差分量问题.在植物遗传育种实践中,通常令负的方差分量为零,再估计一系列遗传参数[9-11].齐明[12]针对林木田间试验获得的不规则、不平衡数据,提出了转化分析法,进行数据处理.对不平衡的全双列杂交试验数据处理,建议采用朱军的MINQUE(1)进行分析,当获得负的方差分量时,采用REML法进行迭代处理[11].不过REML迭代法在参试因子较多时常出现迭代不收敛情形.
从文献[13-14]可以看出,负的方差分量问题并未引起同行足够的重视.本文对杉木双列杂交试验的不平衡数据[同一试验数据(GriffingⅢ)],进行线性模型理论和方差分析;对于同一数据,构建随机模型和混合模型进行处理分析;对于同一试验(GriffingⅢ)非平衡数据,构建随机模型,以转化分析法的结果为参照对象,比较了不同分析方法的优劣,确定线性模型MINQUE(1)法中是否采用平衡数据或随机模型;针对同一试验(GriffingⅢ)非平衡数据,采用最小二乘法补平衡后,再采用Yates F方法进行方差分析,旨在为林木全双列交配试验的数据处理提供一个合适的分析方法,为杉木高世代育种方案的制订提供可靠的遗传参数.
1 材料与方法
1.1 试验点概况
试验点设在浙江省遂昌县大桥村,气候属中亚热带季风类型,冬冷夏热,四季分明,雨量充沛,空气湿润,山地垂直高度500 m,气候差异明显.全年平均气温16.8℃,年降水量1510 mm,降水日数172 d,年太阳总辐射量422 kJ·cm-2,年日照时数1755 h,年无霜期251 d.造林地为马尾松采伐基地,立地条件中等,挖穴40 cm×40 cm×40 cm,造林密度2 m×2 m.1997年春季造林,36个家系,外加一代混种和龙15两个对照,共38份供试材料.田间试验设计为单因素随机区组,单株小区,30次重复.2002年秋季进行全林调查.
1.2 供试材料
杉木6×6全双列交配(GriffingⅢ)试验如表1所示,30个组合,单株小区,抽取28次重复.调查了5年生试验林的胸径、树高、枝下高,估算材积和纵贯比等5个性状.交配设计中的亲本是通过子代测定评选出的 6 个速生亲本,即 A1、A2、A3、A4、A5 和 A6.
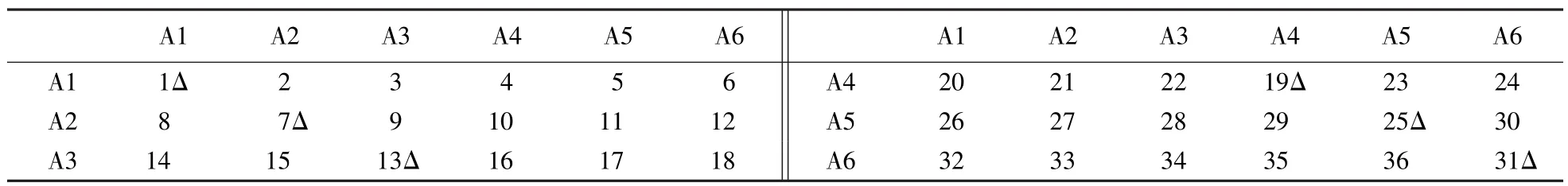
表1 杉木6×6全双列交配设计试验1)Table 16×6 diallel cross design of Chinese fir
1.3 统计方法
1.3.1 方案Ⅰ 采用线性模型理论来处理试验不平衡数据,构建如下随机模型:
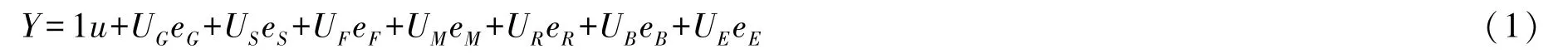
式中,Y为表型值向量,u为群体平均值,UG为变量G已知系数设计矩阵,eG为独立随机变量G向量,US为变量s已知系数设计矩阵,eS为独立随机变量S向量,UF为变量F已知系数设计矩阵,eF为独立随机变量F向量,UM为变量M已知系数设计矩阵,eM为独立随机变量M向量,UR为变量R已知系数设计矩阵,eR为独立随机变量R向量,UB为变量B已知系数设计矩阵,eB为独立随机变量向量,UE为已知系数单位矩阵,eE为独立随机误差向量.所有变量遵循正态分布.
根据欧氏范数最小这一要求,可推出如下方程组:

式中,V1=Ug×Ug′+Us×Us′+Uf×Uf′+Um×Um′+Ur×Ur′+Ub×Ub′+Ue×Ue′

式中,X为固定效应的系数设计矩阵;b为固定效应向量.
按MINQUE(1)法估计各因子的方差分量[8].
1.3.2 方案Ⅱ 采用最小二乘法将缺失数据补平衡[10],构建随机模型和混合模型,采用MINQUE(1)估计各因子的方差分量[8].
1.3.3 方案Ⅲ 采用转化分析法[11]处理不平衡数据,先按模型(3)进行方差分析,然后按GriffingⅢ方法估计各因子的方差分量[式(4)].

式中,Yijk为第i个母本与第j个父本杂交子代在第k个区组中的观察值;u为群体平均值;Fij为第i个母本与第j个父本间的组合值;Bk为第k个区组效应值;Eijk为随机误差.
式中,Yij为组合的平均值;u为群体平均值;gi为第i个亲本的一般配合力;gj为第j个亲本的一般配合力;sij为第ij个组合的特殊配合力;rij为第i个母本与第j个父本的正反交效应.
1.3.4 方案Ⅳ 利用方案Ⅱ的平衡资料,采用Yates混合模型,估计各因子的方差分量,并与线性模型理论结果、GriffingⅢ的转化分析结果进行比较.
Yates和GriffingⅢ方差分析原理见文献[1,12,16];线性模型理论的原理和分析程序参考文献[8,12,16].所有计算过程均采用 matlab7.0平台以及自行开发的程序[12,16]进行数据处理.
为先验值,取值1.
Qa=Va-Va×1×inv(1′×Va×1)×1′×Va, 通过解方程得到参试因子的方差分量.
式(1)~式(4)及 Yates方法的详细计算过程可参考文献[1,8,13,14].
混合模型表示如下:
2 结果与分析
2.1 采用不平衡数据线性模型法获得各因子的方差分量
随机模型条件下,杉木各性状配合力分析结果如表2所示.从表2可知:在6×6全双列交配试验中,生长性状上一般配合力(GCA)的作用居于次要地位,但是特殊配合力(SCA)及正反交(REC)效应的作用均达到极显著水平;干形性状上,GCA的作用极其显著,而SCA及REC效应的作用较弱;而母本细胞质效应对生长性状和纵贯比的表达有显著作用;父系非核基因效应均不存在.采用MINQUE(1)系统处理缺失数据,令表2中负的方差分量为0[9-11].

表2 杉木主要经济性状配合力分析结果(不平衡数据ꎬ随机模式)Table 2 Combination ability of main economic traits of Chinese fir(unbalanced data, random model)
混合模型条件下,采用MINQUE(1)得到的各因子的方差分量估计结果如表3所示.从表3可知:树高和枝下高具有相同的遗传控制方式,一般配合力和母体细胞质效应占主导地位,并存在上位效应,其它因子的作用不显著;材积、胸径和纵贯比、特殊配合力和父本花粉效应占主导地位,一般配合力、母本细胞质效应和正反交效应作用不显著.
通过比较表2和表3的结果发现不同性状各参试因子的遗传方差分量大小和相对份量存在较大差异.材积、胸径和纵贯比3个性状在两种分析方法中,除了特殊配合力方差极显著外,一般配合力、母本细胞质效应的效应相反;混合模型中一般配合力GCA、母本细胞质Female和正反交效应REC不存在;树高和枝下高的结果一致(表2),而表3中树高的GCA极显著,Female和REC效应有一定的作用,但SCA不存在,Male效应在两种模型中均不存在.在相同的性状、数据、方法下采用随机模型和混合模型得到的分析结果差异大.

表3 杉木主要经济性状配合力分析结果(不平衡数据ꎬ混合模型)Table 3 Combination ability of main economic traits of Chinese fir(unbalanced data, mixed model)
2.2 采用平衡数据线性模型法获得的各因子方差分量
根据最小二乘法,将各组合中的缺株值进行填充,使之平衡.采用随机模型条件下的MINQUE(1),估计各因子的方差分量,结果列于表4.从表4可知,材积、树高、胸径和纵贯比都是一般配合力占主导地位,并存在显著的母本细胞质效应,其它遗传因子的作用不显著;而枝下高的特殊配合力占主导地位,并存在显著的一般配力效应和母本细胞质效应;其它遗传因子作用不显著.另外,区组的方差分量作用显著,表明试验地存在明显的林地异质性.

表4 杉木主要经济性状配合力分析结果(平衡数据ꎬ随机模式)Table 4 Combination ability of main economic traits of Chinese fir(balance data, random model)
在平衡数据的混合模型条件下,MINQUE(1)分析结果列于表5.从表5可知:材积以特殊配合力效应占主导地位,其次是正反交效应,一般配合力对材积也有显著影响,父母本非核基因作用不明显;树高以上位性、特殊配合力占主导地位,其它遗传因子作用不显著;胸径则以一般配合力占主导地位,特殊配合力显著,但居次要地位,父母本非核基因效应不显著;枝下高以母本细胞质基因效应占主导地位,上位性和特殊配合力也有显著作用,其它遗传因子作用不明显;纵贯比以上位性方差占主导地位,其次是一般配合力,特殊配合力作用居其后,其它遗传因子作用不明显.
2.3 非平衡条件下杉木6×6全双列杂交试验的转化
杉木6×6各性状在组合间的差异F检验结果列于表6.从表6可知,6×6全双列交配试验中,杉木主要经济性状在不同组合间都存在极显著差异,这是全同胞家系选择的依据,也是进一步对其遗传差异的成因进行配合力分析的前提.
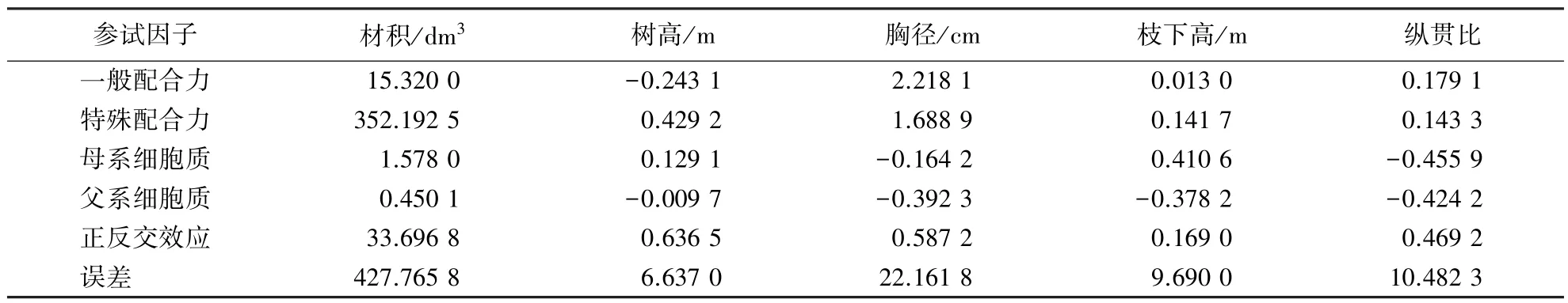
表5 杉木主要经济性状配合力分析结果(平衡数据ꎬ混合模型)Table 5 Combination ability of main economic traits of Chinese fir(balance data, mixed model)

表6 杉木6×6全双列交配试验各性状方差分析结果1)Table 6 Analysis of variance of Chinese fir traits in 6×6 diallel cross design
根据Griffing方法Ⅲ的随机模型以及配合力分析结果,进一步估计杉木主要经济性状的遗传参数,结果列于表7.从表7可知:在杉木6×6全双列交配试验中,在生长性状上,非加性遗传基因效应起主导作用,加性基因效应也起一定作用;而在干形性状上,加性基因效应起主导作用,非加性基因效应居次要地位.结果表明,杉木选择改良性状(生长性状)的基因作用方式,与改良的性状有关.在杂合体居多的交配群体中,非加性遗传方差占主导地位,加性遗传方差居次要地位,研究性状的遗传控制方式是由交配亲本群体的遗传结构决定的[17].

表7 杉木6×6全双列交配试验主要经济性状的遗传方差Table 7 Genetic variance of main economic traits of Chinese fir in 6×6 diallel cross design
2.4 平衡数据混合模型条件下各参数方差分量的Yates估计结果
采用最小二乘法,将各杂交组合的缺值补齐,形成一个平衡试验数据.采用Yates提出的混合模型进行方差分析,估计各参试因子的方差分量,结果列于表8.

表8 采用Yates法估计的各参试因子的方差分量1)Table 8 Variance component size of each parameter estimated by Yates method
由表8可知,生长性状以特殊配合力占主导地位,一般配合力居次要地位,母体细胞质效应也有显著作用;而枝下高和纵贯比有着相反的遗传控制方式,一般配合力居主导地位,特殊配合力也有显著的作用,母体细胞质效应的作用不明显.比较表8和表7的结果可以发现,在所有研究性状上,方差分析获得的各参试遗传因子的方差分量的相对大小完全一致;且能获得母本细胞质效应.从获得的信息量看,该方法比转化分析法和线性模型法更好.在平衡数据的情形下,比较表8和表5的结果可以发现,材积和树高的结果相同,而胸径、枝下高和纵贯比的参试遗传因子的方差分量相对大小不同.由于表8提供的信息量和参数的分量大小与表2、7中的结果一致,且没有负的方差分量,所以Yates法的结果(表8)更具有科学性、合理性和可靠性.
3 小结
本研究结果表明:(1)对于杉木全双列设计(GriffingⅢ)的非平衡试验资料,构建随机模型,采用MINQUE(1)法获得最多的信息量,参试因子的方差分量与方差分析结果有一致的变化趋势,但常获得负的方差分量;同时采用线性模型法没有必要对不平衡资料进行补平.
(2)对于杉木全双列设计(GriffingⅢ)的非平衡试验资料,采用转化分析法可以获得合适、可靠的方差分量,并避免了负的方差分量,但获得的信息量低,参试因子的方差分量可能有较大的抽样方差[18].
(3)对于杉木全双列杂交(GriffingⅢ)试验的非平衡数据处理方法的选择,如果试验目的是为了获得方差分量和参试因子的效应值,可采用最小二乘法将数据补平衡,建立混合模型,采用Yates法处理杉木全双列杂交试验(GriffingⅢ)资料.平衡资料没有负的方差分量,而且可获得母本细胞质基因的信息;更重要的是平衡数据的方差分析具有参试因子方差最小和估计值无偏性[10,18,19]的优点.杉木全双列杂交(GriffingⅢ)试验资料的数据处理,Yates法优于其它方法.
随机模型条件下采用MINQUE(1)法得到的结果信息量高,但会出现负的方差分量.转化分析法是对试验设计的角度进行转化,从而改进参试因子的正态性.Yates法通过缺株补齐来改变参试因子的正态性,从而避免了负的方差分量.综上所述,Yates法具有最小方差和参数无偏性优点,权衡获得信息量和克服负的方差分量,对于杉木全双列杂交试验(GriffingⅢ)非平衡数据处理,Yates法最佳.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
中学生数理化(高中版.高考数学)(2021年3期)2021-06-09
农民致富之友(2020年8期)2020-05-11
中学生数理化·七年级数学人教版(2019年6期)2019-06-25
中学生数理化·七年级数学人教版(2019年6期)2019-06-25
中国种业(2018年11期)2018-11-16
猪业科学(2018年5期)2018-07-17
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
初中生世界·九年级(2017年10期)2017-11-08
