
戴云山黄山松种群径级结构的海拔分布格局
2019-10-09 01:46何中声刘金福冯雪萍刘艳会陈文伟
福建农林大学学报(自然科学版) 2019年5期
江 蓝, 何中声, 刘金福, 冯雪萍, 刘艳会, 陈文伟
(1.福建农林大学林学院,福建福州350002;2.福建农林大学海峡自然保护区研究中心,福建福州350002;3.福建省高校生态与资源统计重点实验室,福建福州350002;4.戴云山国家级自然保护区,福建泉州362503)
种群是群落的基本组成单位,其径级结构既可反映种群各径级个体的分布情况,揭示种群数量动态特征,又可反映种群与环境间的互作过程及其在群落中的作用,对阐明种群更新对策、生态特性以及群落演替规律具有重要作用[1,2].种群结构及其动态变化一直都是植物生态学的研究热点之一.海拔梯度在较小空间范围内反映了气候、地形与土壤等因子的差异,造成水热条件的再分配,为研究植物种群数量动态对环境梯度的响应提供了良好平台[3,4].开展径级结构对海拔梯度响应的研究对了解物种不同生长阶段的海拔适应范围及分布规律具有重要作用,为指导森林保护和可持续经营提供理论基础.
黄山松(Pinus taiwanensis)也称台湾松,为中国特有种,材质好,耐干旱,抗风,是亚热带中山地区的主要优势种,主要分布在福建、台湾、浙江、江西与安徽等省份.戴云山保存有超过6000 hm2的黄山松群落,是中国大陆最南端、保存最好、面积最大的黄山松种质基因基地[5-7].目前,关于戴云山植物种群结构研究集中在当地优势树种的径级组成[8].段仁燕[9]研究发现太白红杉(Larix chinensisBeissn)年龄结构在不同海拔梯度下存在显著差异,低海拔以中到老龄个体为主,而高海拔以小径级为主;张维等[10]以胸径结构代替年龄结构,发现野核桃种群(Juglans cathayensisDode)的胸径级随海拔升高逐渐减小,高海拔以小径级个体居多;张育新等[11]研究发现,辽东栎(Quercus wutaishanicaMayr)种群的平均胸径以海拔1480 m为界点呈两段式分布,在每一段内随海拔的增加平均胸径也随之增大.为此,根据戴云山黄山松种群的分布范围,探讨戴云山黄山松种群结构随海拔梯度的变化情况,以及种群径级结构随海拔梯度的变化及其影响因素,旨在为其种群的可持续经营提供理论依据.
1 研究区概况
戴云山国家级自然保护区(25°38′07″—25°43′40″N,118°05′22″—118°20′15″E)总面积 13472.4 hm2,属南亚热带与中亚热带过渡区.保护区年均温15.6~19.5℃,年降水量1700~2000 mm[12].戴云山成土母质由花岗岩风化而成,土壤主要类型有山地红壤、山地黄壤、赤红壤、山地黄红壤和泥炭沼泽土.林地土壤腐殖质和枯枝落叶层较厚,表土质地为壤土,土壤呈酸性,有机质含量较高,含水量适宜,理化性质较好[12].植物资源丰富,种类繁多,森林覆盖率达93.4%.森林植被具有明显的垂直分布特征,随海拔升高依次出现常绿阔叶林(700~1000 m)、针阔混交林(1000~1200 m)、温性针叶林(1200~1650 m)、苔藓矮曲林(1650~1765 m)与草甸(1750 m以上)垂直带谱.植物区系兼具中、南亚热带性质,主要保护对象是东南沿海典型的山地森林生态系统[13].戴云山群落层次分明,群落乔木层主要有黄山松、罗浮栲(Castanopsis faberiHance)与江南山柳(Clethra cavalerieiLevl.)等,灌木层主要有岩柃(Eurya saxicolaH.T.Chang)、窄基红褐柃(E.rubiginosavar.attenuataH.T.Chang)与钝齿冬青(Ilex crenataThunb.)等,草本层主要有里白[Hicriopteris glauca(Thunb.) Ching]、狗脊蕨[Woodwardia japonica(L.f.) Sm.]与淡竹叶(Lophatherum gracileBrongn.)等[8].
2 方法
2.1 样方布设
在戴云山黄山松海拔分布区(1000~1700 m),自1000 m起,沿海拔将每100 m范围设置一个梯度,共8个梯度.每个梯度选择未受人为干扰的3个25 m×25 m固定样地,布设时尽可能保持各样地生境因子一致,并记录样地海拔、坡度与坡向等地形因子.调查林分郁闭度、黄山松和林分密度.对样方内胸径<1 cm的物种只记录种名、数量;对胸径≥1 cm物种,记录种名,测量胸径、树高和冠幅.记录样方内死亡黄山松枯木数量,并以黄山松枯木数与黄山松总数的比值表示黄山松死亡率.
2.2 径级结构划分
参照苏松锦对黄山松的研究方法[14],将黄山松按胸径大小以5 cm为间距划分为8个等级,统计各径级黄山松个体数,以纵坐标表示径级,以横坐标表示个体数,绘制黄山松种群径级结构图.等级划分如下:DBH≤2.5 为Ⅰ级;2.5 cm<DBH≤7.5 cm 为Ⅱ级;7.5 cm<DBH≤12.5 cm 为Ⅲ级;12.5 cm<DBH≤17.5 cm为Ⅳ级;17.5 cm<DBH≤22.5 cm 为Ⅴ级;22.5 cm<DBH≤27.5 cm 为Ⅵ级;27.5 cm<DBH≤32.5 cm 为Ⅶ级;32.5 cm<DBH≤37.5 cm 为Ⅶ级;DBH>37.5 cm 为Ⅷ级.
根据黄山松种群径级结构,按其生活史周期划分出4个生长阶段,分别为幼苗、幼树、中树和大树.按照空间代时间的方法,以胸径大小代替生存时间,胸径为48.5 cm(最大)的黄山松可看作是最大年龄个体.以黄山松最大年龄为基准,胸径2.5~12.5 cm的个体占总树龄的5%~25%,可视为黄山松幼龄阶段,故将Ⅱ和Ⅲ级划为黄山松幼树.中树和大树也按照此方法划分[14,15].因此,定义DBH≤2.5 cm为幼苗;2.5 cm<DBH≤12.5 cm 为幼树;12.5 cm<DBH≤22.5 cm 为中树;DBH>22.5 cm 为大树.
2.3 数据处理
为了解群落特征对黄山松种群的潜在影响,群落基本概况选取郁闭度、林分密度、黄山松密度、群落平均胸径、黄山松胸径、群落平均树高、黄山松树高及黄山松死亡率8个指标.其中,林分密度为单位面积上的林木株数,是影响林木发育的重要因子[16].群落平均胸径和平均树高为样方内所有林木胸径和树高的平均值,是群落概况的基础指标.
首先通过Excel计算整个海拔范围内黄山松的径级组成;然后统计黄山松幼苗、幼树、中树和大树比例随海拔梯度的变化情况,分析黄山松径级比例的海拔梯度响应;最后,利用SPSS 17.0探讨各指标之间的相关性.采用Pearson相关系数对黄山松幼苗比例与大树比例的相关性进行分析.
3 结果与分析
3.1 群落基本概况
从表1可知,林分密度随海拔的升高呈先升后降再升的变化趋势,在1000~1300 m海拔高度的林分密度均随海拔的升高逐渐增大,但在1200~1300 m第1个峰值后,在1300~1400 m海拔高度林分密度下降,随后开始上升.群落平均胸径、黄山松胸径、群落平均树高和黄山松树高随海拔的变化规律相似,均在1000~1100 m具有最大值,在1100~1200 m海拔逐渐下降,在1200~1300 m海拔上升,在1300~1600 m海拔再次降至最低值.而黄山松死亡率随海拔无明显变化.黄山松平均死亡率最高值为6.7%,出现在海拔1300~1400 m.

表1 黄山松群落基本概况1)Table 1 General information of P.taiwanensis community
3.2 黄山松种群径级结构
由图1可知,戴云山自然保护区黄山松种群幼苗(Ⅰ级)数量稀少,幼树(Ⅱ~Ⅲ级)与中树(Ⅳ~Ⅴ级)数量较多,大树(Ⅵ~Ⅷ级)较少.由图2可知,戴云山共调查到黄山松2333株,黄山松种群以幼树为主,有1658株,占总体数量的71.1%;中树次之,为600株,占黄山松总量的25.7%;幼苗和大树稀少,分别为16株(0.7%)和59株(2.5%).戴云山黄山松种群幼树数量最多,中树次之,幼苗和大树稀少.虽然黄山松幼苗稀少,但幼树、中树数量较多,表明目前黄山松种群处于增长型向稳定型的过渡阶段,在当地群落中仍将占优势地位.
3.3 黄山松径级比例的海拔梯度分布格局
由图3可知,黄山松幼苗在1000~1100 m和1200~1500 m海拔没有分布,在1500~1600 m海拔幼苗比例最大,占1500~1600 m黄山松总数的4%.幼树比例随海拔呈明显的规律性变化,1000~1200 m幼树比例随海拔的升高而升高,在1200~1300 m突然下降,在1300~1400 m又随海拔的升高而升高.黄山松幼树比例在1000~1100 m海拔最低,为30%;在1600~1700 m海拔最高,为98%.中树比例的海拔变化规律与幼树恰好相反,呈两段式下降分布.1000~1100 m海拔中树比例最高,为44%,在1000~1200 m海拔中树比例随海拔的升高而下降,在1200~1300 m突然上升,在1300~1400 m又随海拔的升高而降至最低值(2%).大树比例在1000~1100 m最高,为26%,且随海拔的升高呈下降趋势,最低值为0.
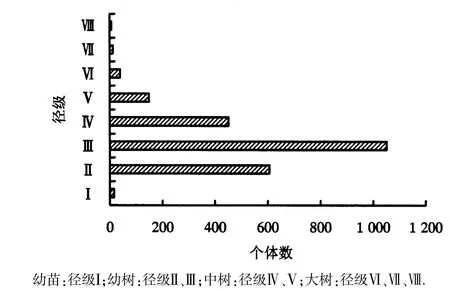
图1 黄山松种群的径级结构Fig.1 Diameter structure of P.taiwanensis community

图2 黄山松不同生长阶段个体数分布图Fig.2 Distribution of P.taiwanensis individuals at different growt

图3 黄山松种群径级比例沿海拔梯度变化Fig.3 Changes in the proportion of P.taiwanensis at different diameter along elevation gradients
3.4 相关性分析
为探讨各指标之间是否具有相互影响,采用相关分析进行研究,结果见表2.从表2可知,幼苗比例与黄山松胸径、黄山松树高和群落平均树高之间均呈显著负相关关系,即幼苗随群落内中大树的增多而逐渐减少.幼树比例与林分密度呈显著正相关,与群落平均胸径、黄山松胸径、群落平均树高与黄山松树高呈极显著负相关.中树比例与林分密度呈显著负相关,与幼树比例呈极显著负相关,与黄山松胸径呈显著正相关,与群落平均胸径、黄山松胸径、群落平均树高和黄山松树高呈极显著正相关.大树比例与群落平均胸径、黄山松胸径和黄山松树高呈极显著正相关,与幼树比例呈显著负相关.随大树比例的增大,幼树比例降低.随着群落内中大树的增多,幼苗、幼树比例逐渐下降,表明黄山松种群可能存在自疏效应.

表2 各指标之间的相关性分析1)Table 2 Correlation analysis between different indicators
4 小结与讨论
戴云山黄山松种群在1000~1700 m分布区以幼树和中树为主,幼树数量庞大,径级结构显示黄山松种群处在增长型向稳定型的过渡阶段,黄山松在群落演替过程中占据优势地位.但本研究发现,黄山松幼苗仅16株,占总体数量的0.7%,幼苗数量明显不足.黄山松幼苗数量稀少与种子自身特性、生境条件及林内竞争等有关.朱德煌等[17]对黄山松群落主要树种更新生态位的研究发现,黄山松与窄基红褐柃、短尾越桔(Vaccinium carlesiiDunn)等物种具有较大的生态位重叠,资源需求相似,林下资源与空间竞争激烈,阔叶种对黄山松排斥作用明显,不利于黄山松幼苗生长.此外,黄山松对光需求大,耐荫性弱,当林分郁闭度大于0.5时,幼苗难以存活[18,19].而戴云山黄山松群落平均郁闭度0.59,林下光照不足,从而限制了黄山松幼苗的生长.另外,群落灌草层植被丰富,黄山松种子难以到达土壤层,阻碍了种子的正常萌发,种子发芽率仅27%[18],低发芽率也是幼苗数量稀缺的原因之一.苏松锦等[20]对黄山松幼苗更新与土壤有机碳关系的研究发现,黄山松更新苗主要分布在特定的有机碳密度斑块中,土壤有机碳的空间异质性对幼苗更新具有抑制作用.黄山松抗人为干扰能力弱,恢复能力也不强,幼苗一旦受到干扰便很难更新[21].林下光照不足,种间、种内竞争,种子发芽率低,土壤理化特性及外界干扰等原因共同造成黄山松幼苗数量稀少的现状,这可能是未来黄山松群落面临的潜在威胁.
黄山松幼苗、幼树、中树和大树随海拔上升的分布具有不同的变化规律,幼苗主要分布在高海拔,幼树随海拔的升高比例逐渐增大,中树比例逐渐下降,大树分布于低海拔区.总体表现为低海拔以较大径级的中树、大树为主,中高海拔以小径级的幼苗、幼树为主,随海拔的升高呈大径级树逐渐减少、小径级树逐渐增多的规律,这与张维等[10]对不同海拔梯度上新疆野核桃种群研究的结果一致.黄山松种群的这种海拔分布特征是种群发育历史、物种特性、生长策略、环境和干扰等多种因素综合作用的结果.可以推测黄山松种群是从低海拔向高海拔演替的,高海拔黄山松是由低海拔母株扩散后生长繁殖的.此外,随海拔的升高温度降低也是造成低海拔以中树大树为主,以及高海拔小径级树占优势的原因之一[22].
林分密度对黄山松幼树和中树的影响效应不同,林分密度与黄山松幼树呈显著正相关,与中树呈显著负相关,表明随着海拔的升高林分密度增大,黄山松幼树比例增大,中树比例下降,致使群落内黄山松个体幼龄化.此外,林分密度与群落平均胸径和群落平均树高也有显著的负相关关系,再次说明密度增大会促使群落物种胸径减小、树高降低,个体幼龄化趋势明显.密度与胸径之间的关系在多数研究中均有发现,张育新等[11]发现不同海拔梯度辽东栎种群密度与种群平均胸径和聚集程度有显著负相关关系;周威等发现林分密度与群落中各径级平均胸径呈负相关关系,其关系式可用幂函数拟合[23];谌红辉等[24]研究发现林分胸径随冠幅随造林密度的增大而减小,自然稀疏强度增大.林分密度增大,植物个体数增多,林分郁闭度降低,导致植物生长空间受限,林分环境改变,光照、水肥等资源竞争加剧,产生种群自然稀疏效应,限制了植物生物量的积累,制约其生长.本研究也发现黄山松大树与幼树、中树与幼树之间存在资源竞争关系,该结果与苏松锦等[14]对黄山松种群的研究结果一致,表明戴云山黄山松种群存在自疏效应,同种个体竞争排斥作用明显.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
河北果树(2020年1期)2020-02-09
中华魂(2018年11期)2018-05-14
黄山学院学报(2017年5期)2017-11-03
现代园艺(2017年11期)2017-06-28
土壤与作物(2015年3期)2015-12-08
武夷学院学报(2015年3期)2015-07-18
西北植物学报(2015年9期)2015-07-05
中南林业科技大学学报(2015年8期)2015-05-22
