
尼洋河流域高山松次生林林分因子与林龄的相关性
2019-10-10 02:27王小兰陈甲瑞杨小林张传龙
西北农林科技大学学报(自然科学版) 2019年11期
王小兰,陈甲瑞,杨小林,张传龙
(西藏农牧学院 a 资源与环境学院,b高原生态研究所,c西藏高原森林生态教育部重点实验室,d西藏林芝高山森林生态系统国家野外科学观测研究站,西藏 林芝 860000)
高山松(Pinusdensata)是云南松(P.yunnanensis)与油松(P.tabulaeformis)自然杂交产生的二倍杂交种,其分布占据了两个亲本种都不能正常生长的高海拔、高寒地带[1-2],主要分布于青藏高原东南缘,包括四川西部、青海南部、西藏东部及云南西北部高山地区,为我国西部高山地区的特有树种[3]。在西藏主要集中分布于波密、朗县、林芝、米林等地海拔2 500~3 900 m的河滩、台地、阶地、沟谷以及阳坡、半阳坡、半阴坡地带,阴坡也有少量分布[4]。高山松生长迅速,树干通直饱满,木材坚韧。其结实丰盛,种子小而具翅,易飞散,对环境的适应性强,能在其他针叶树种不能生长的干燥、贫瘠立地上生长,是迹地更新和荒山绿化的重要先锋树种,在藏东南地区发挥着保持水土和涵养水源等重要生态功能[3]。
自20世纪80年代以来,很多学者围绕高山松的种实与繁殖特性[2]、幼苗生长特性[5-7]、天然林更新特征[8-11]、种群结构特征[12-14]等开展了一系列研究,为高山松的生态适应性、种群保护等提供了重要的科学依据。尼洋河作为雅鲁藏布江的重要支流之一,高山松林的面积约占该流域森林面积的37%,蓄积量约占27%[9]。由于过去的无计划采伐和人为干扰,尼洋河流域现有的高山松林多为天然次生林,但对这些次生林林分结构、密度、蓄积量及更新状况等林分特征的研究尚较为缺乏。为此,本研究调查了这一地区高山松林的生长特性及林分特征,分析胸径、树高、断面积、蓄积量、幼苗更新等主要林分因子随林龄的变化规律,以期为类似区域合理保护和科学经营高山松天然次生林提供科学依据。
1 研究区概况
研究区域气候属于高原温带半湿润至半干旱季风气候,春夏多雨,秋冬干燥少雨,大气相对湿度64%,年降水量600~800 mm,年平均气温7 ℃,最冷月(12月至翌年1月)平均气温-0.3 ℃,最热月(7-8月)平均气温15.6 ℃,无霜期150 d以上[15]。土壤以山地棕壤和酸性棕壤为主。植被垂直地带性较明显,主要包括各种针叶林以及硬叶常绿阔叶林、针阔混交林和落叶阔叶林[16]。高山松林主要分布在海拔3 700 m以下的尼洋河下游,在西藏林芝市的百巴镇、更张乡、八一镇等地常形成大面积纯林,主要由天然更新形成,其郁闭度在0.5左右,处于中幼龄林阶段。林内乔木层主要树种为高山松,伴生有川滇高山栎(Quercusaquifolioides)、糙皮桦(Betulautilis)、山杨(Populusdavidiana)、西藏落叶松(Larixgriffithiana)等,灌木层主要有雅致山蚂瑝(Desmodiumracemosum)、小叶丁香(Syringamicrophylla)、林芝杜鹃(Rhododendronnyingchiense)、米饭花(VacciniuimmandarinorumDiels)等,草本层主要有蕨(Pteridiumaquilinum)、柴胡(BupleurumtriadiatumAdams)、委陵菜(Potentillachinensis)等,另有藤本植物防己叶菝葜(Smilaxmenispermoidea)等。
2 研究方法
2.1 标准地设置与调查
研究区位于西藏东南部尼洋河流域,流域内主要行政区有林芝市的尼西镇、八一镇等地。从林芝市八一镇出发,沿尼洋河流域阳坡和半阳坡选择能代表区域种群特性的高山松纯林进行典型样地调查,共设置17个20 m×20 m的标准地。对标准地内所有乔木树种(胸径≥2 cm)进行每木检尺,并记录其树种、树高、胸径、第一枝下高、冠幅。然后选取接近平均胸径的3~5株标准木,利用生长锥钻取木芯,运用LINTABTM6年轮分析仪确定标准木年龄。在每个标准地内按照五点取样法设置5个2 m×2 m的小样方进行高山松更新苗调查。在每个小样方内调查所有树高<2 m或胸径<2 cm的更新苗,记录其数量、高度、地径等。同时记录样地的环境因子,包括经纬度、海拔、坡向、坡度。17个标准地主要林分因子见表1。
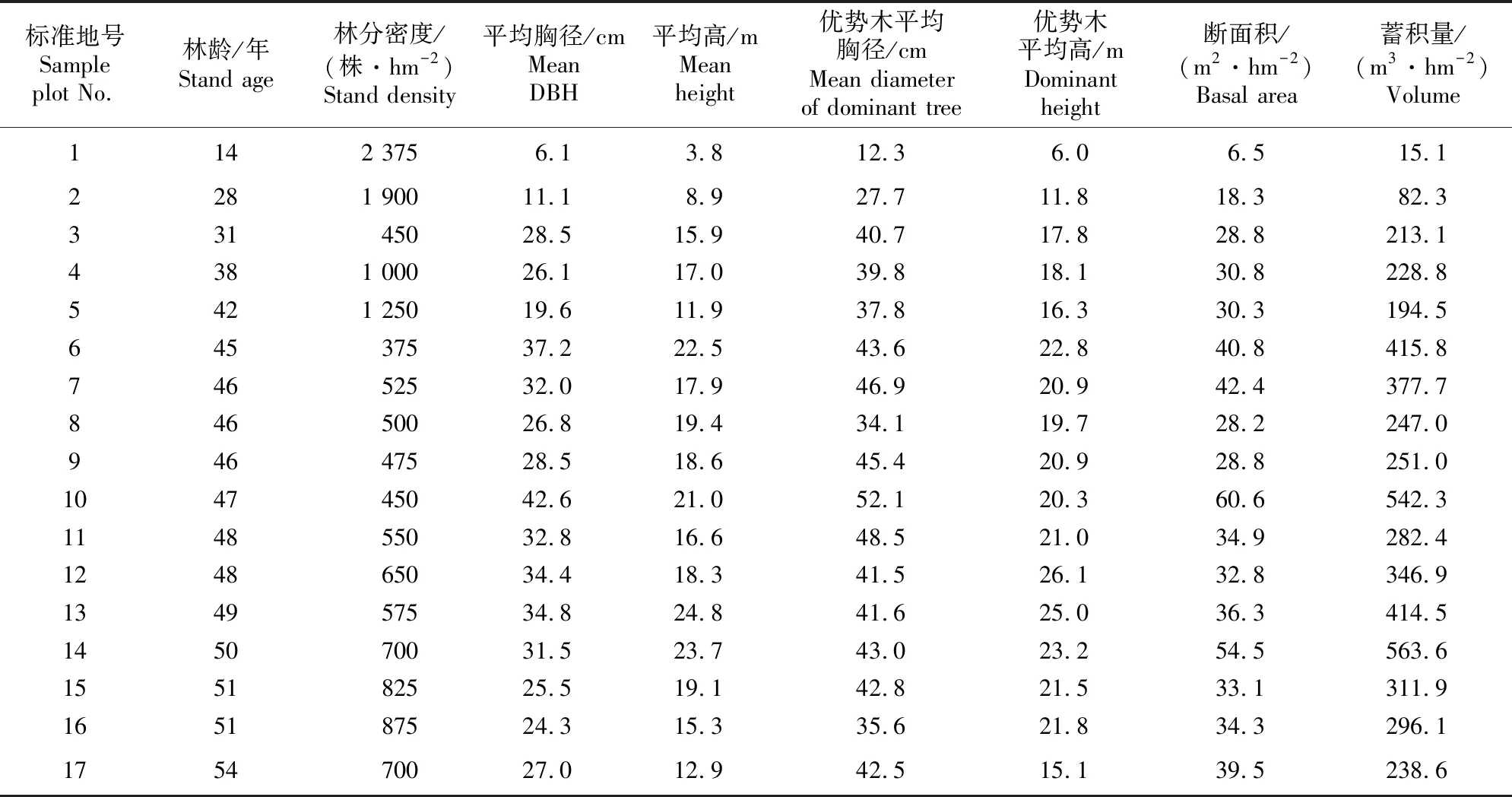
表1 尼洋河流域高山松次生林的主要林分因子
2.2 林分因子及林龄的测定
本研究中林分因子主要包括林分密度、林分平均胸径、林分平均高、优势木平均胸径、优势木平均高、林分胸高断面积和林分蓄积量,各林分因子的含义与计算方法如下[17]:
(1)林分平均胸径(Dg)。指林分中胸径≥2 cm的所有林木胸径的平均值。计算公式为:
式中:di表示第i株林木的胸径,N表示林分内林木的总株数。
(2)林分平均高。在树高曲线上,与林分平均直径相对应的树高称为林分平均高。此次调查中,在林分中选择3~5株与林分平均直径相近的“平均木”测量树高,以其算数平均数作为林分平均高。
(3)优势木平均胸径与平均高。指林分中优势木的胸径和树高的算数平均值。
(4)林分胸高断面积(G)。按下式计算:
式中:gi表示第i株林木的胸高断面积(m2),N表示林分内林木的总株数。
(5)林分蓄积量(V)。 参照西藏地区的高山松二元材积表[4]计算,公式如下:
V=0.463 136 93×10-4×D1.811 366 1×H1.140 230 3。
式中:D表示林木胸径(cm),H表示林木树高(m)。
(6)林分平均年龄(林龄)。在林分内,当查定树龄的林木株数较多时,采用断面积加权法计算林分平均年龄。即:
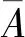
数据处理和分析均采用SPSS 23. 0和Excel 2010软件进行。
3 结果与分析
3.1 高山松次生林林分的胸径分布
林分生长过程中,随着林龄的增加和个体的生长,促使林木发生分化,从而出现径阶分布的明显差异[18]。将样地中的高山松按2 cm径阶分组整理,不同林龄高山松林分的胸径分布状况如图1所示。从图1可以看出,高山松林分在不同林龄阶段表现出不同的胸径分布规律。在幼龄林(林龄<30年)中,林分胸径分布呈现出左偏的近似正态分布曲线,林木主要分布在4~8 cm径阶,占林木总株数的58.2%,呈现“增长型”趋势;有少量胸径超过16 cm的林木出现在幼龄林中,占林木总株数的8.7%;从树种组成来看,幼林龄中其他伴生树种主要由高山栎、西藏落叶松、山杨等组成,占林木总株数的14.7%,主要分布在6~8 cm径阶。在中龄林(林龄40~50年)中,林分胸径分布同样呈现出中间高两边低的近似正态分布;与幼龄林相比,分布曲线的偏度变小,峰度也变小;林木主要集中在28~42 cm径阶,占总株数的54.6%;另外,小径阶林木(4~10 cm)在中龄林中占19.3%,呈现“稳定型”趋势,在这些小径阶林木中,其他伴生树种如川滇高山栎和山杨,所占比例略高于高山松,分别为56.7%和43.3%;而在中大径阶(胸径>14 cm)林木中,高山松为主要优势树种,并无其他树种出现。
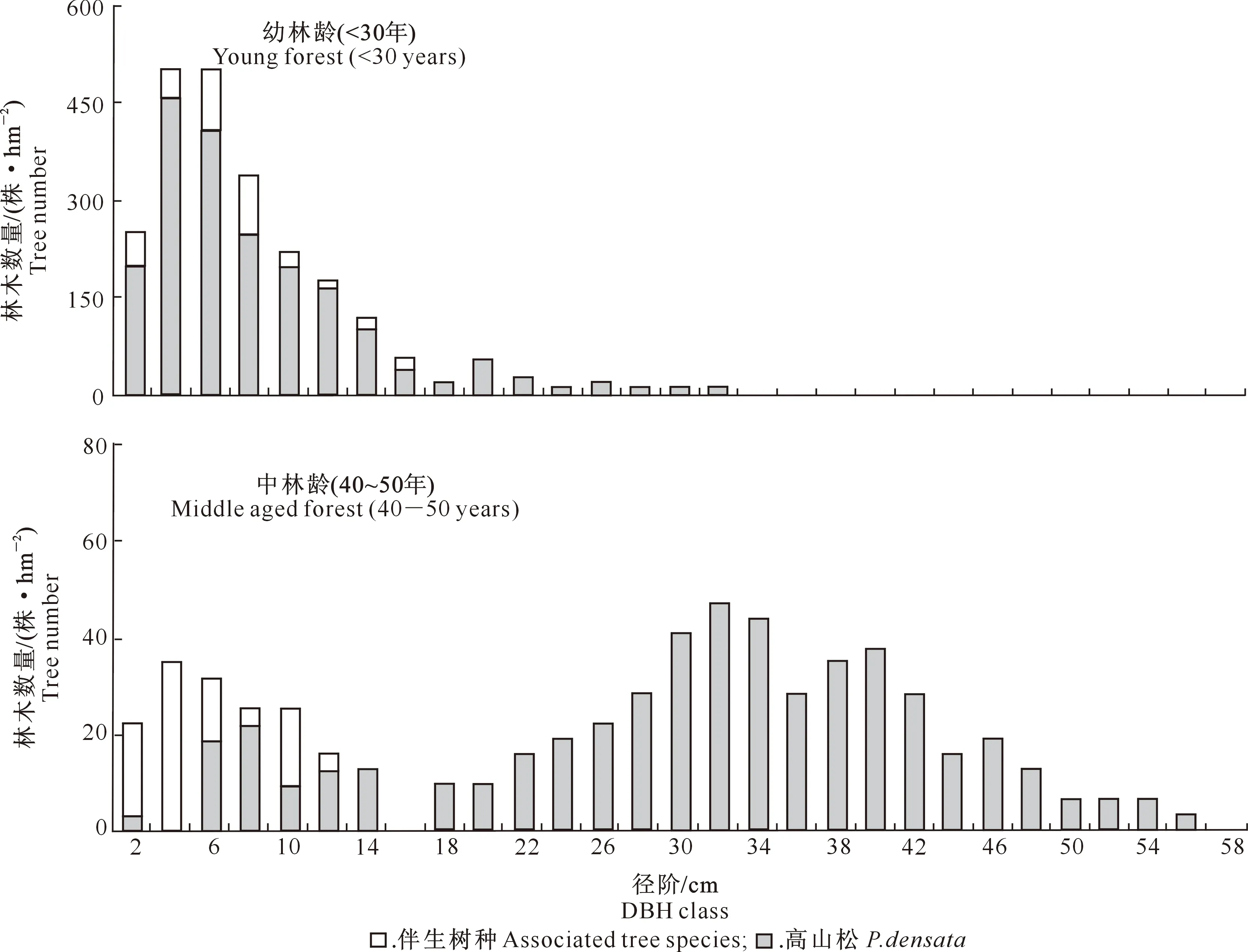
图1 尼洋河流域不同林龄高山松次生林的胸径分布
3.2 高山松次生林主要林分因子随林龄的变化
3.2.1 林分密度 林分密度是影响林分生长和造成林木间竞争的主要原因之一,同时密度也是营林工作中能够有效控制的因子之一[17]。试验区现有高山松纯林密度变化较大,平均为(833.8±546.1) 株/hm2,最小为375株/hm2,最大为2 375株/hm2,并且与林龄相关性显著(图2),即随着林龄的增长,林分密度逐渐下降。林分密度与林龄之间的关系以抛物线方程拟合最好,拟合方程为y=-145.961x+1.494x2+4 203.787,R2=0.816。高山松林在幼林龄阶段(<30年),林分密度随着年龄的增长急剧下降。从拟合方程计算得出,当林龄为15年时林分密度为2 350株/hm2,而林龄为30年时密度为1 169株/hm2;但是当高山松林进入中龄林阶段(40~50年),林分密度变化不大,平均为(600±152.6) 株/hm2。
3.2.2 林分平均胸径和平均高 高山松林林分平均胸径为(27.6±6.0) cm,平均高为(16.9±5.3) m,且与林龄呈显著的正相关关系(图3)。林分平均胸径与林龄之间的相关性以幂函数方程拟合最好,拟合方程为y=0.275x1.210,R2=0.762;林分平均高与林龄之间的相关性同样以幂函数方程拟合最好,拟合方程为y=0.152x1.254,R2=0.829。相对于林分平均胸径而言,林分平均高与林龄之间的拟合性更好,即林分平均高受林龄的影响较大,而林分平均胸径除了受林龄的影响外,还受林分内其他因子,如林分密度、地形等的影响。
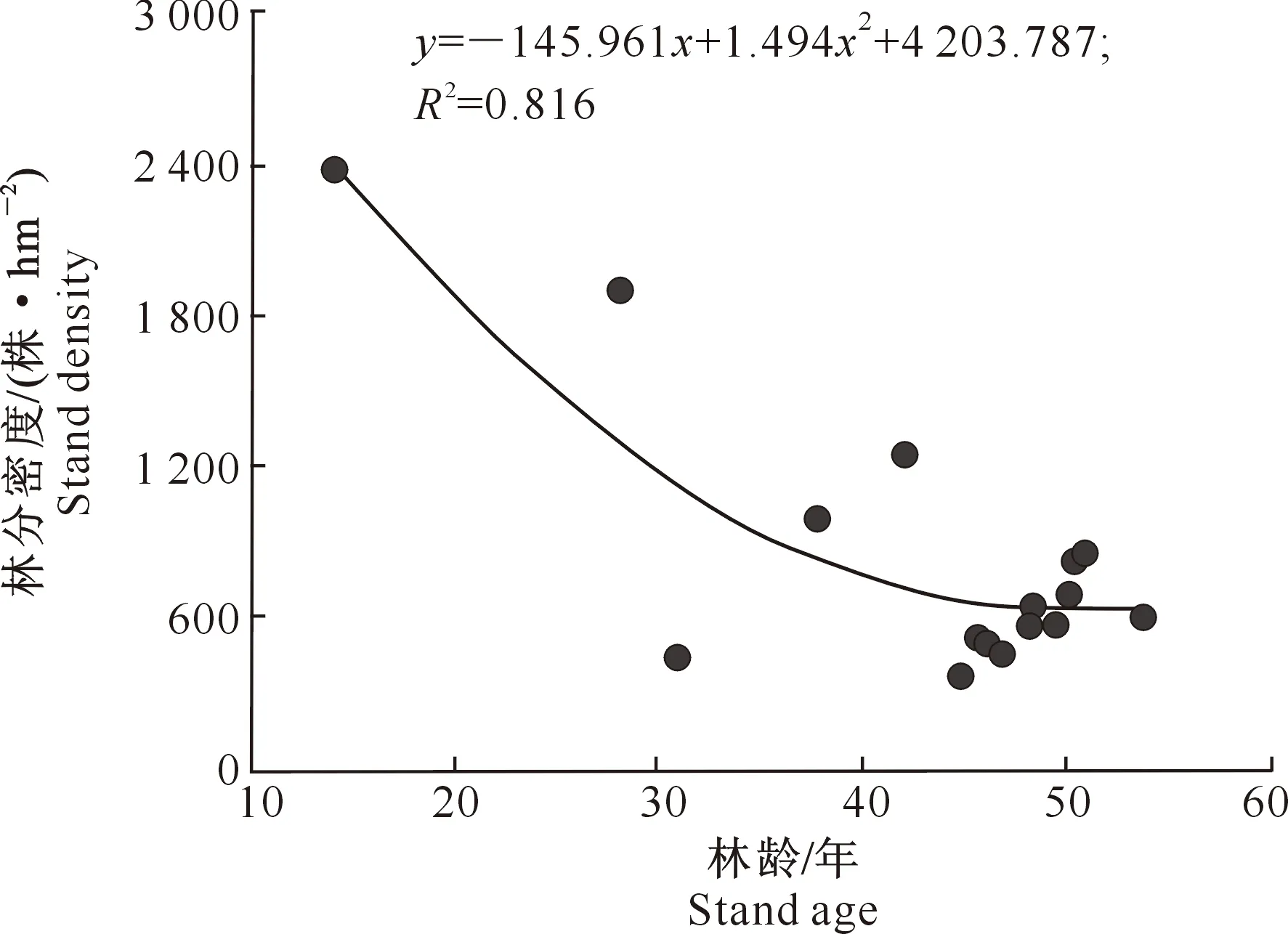
图2 尼洋河流域高山松次生林林分密度与林龄的关系
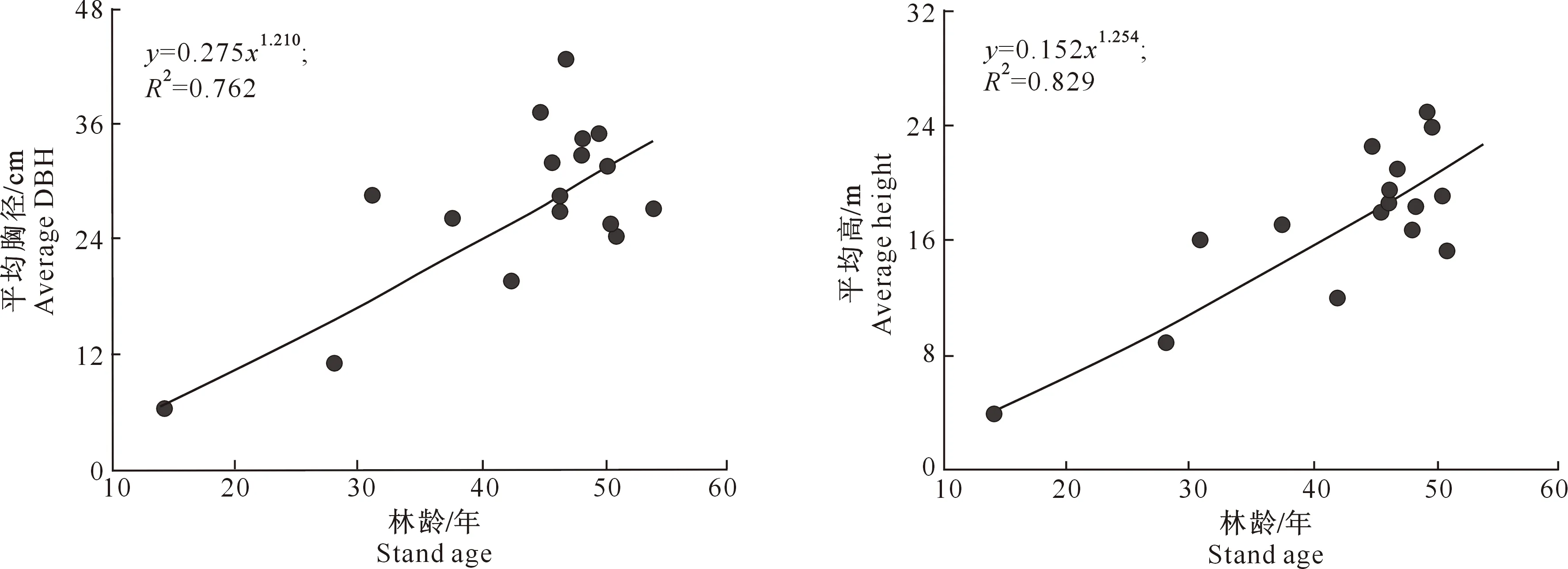
图3 尼洋河流域高山松次生林林分平均胸径和平均高与林龄的关系
3.2.3 优势木平均高和平均胸径 林分优势木平均树高、平均胸径是表征林分生产潜力的重要数据[19]。高山松林林分中优势木平均胸径和平均高分别为(39.8±9.1) cm和(19.3±4.9) m。同林分平均胸径和平均高一样,优势木的平均胸径和平均高与林龄也具有显著的相关性(图4),幂函数方程能很好地拟合优势木胸径和树高与林龄的关系,拟合方程分别为y=1.443x0.880(R2=0.790)和y=0.425x1.020(R2=0. 914)。相对于优势木平均胸径,优势木平均高与林龄之间的相关性更高。
3.2.4 林分胸高断面积和蓄积量 高山松林林分平均胸高断面积为(34.2±12.2) m2/hm2,平均蓄积量为(295.4±142.2) m3/hm2。高山松林林分胸高断面积和蓄积量与林龄呈极显著正相关(图5),二者与林龄之间的回归关系用幂函数方程拟合最好,拟合方程分别为y=0.255x1.284(R2=0.853)和y=0.027x2.453(R2=0.885),拟合精度均很高,R2达到0.85以上。从回归曲线可以看出,这一地区的高山松林具有很高的生产力,当林分进入中龄林阶段(40~50年)时,林分胸高断面积和蓄积量分别达到38.7 m2/hm2和397.1 m3/hm2,而且从林分蓄积量的每年增长量来看,随林龄的增加蓄积量的年增长量也相应增加。例如林龄从20到30年,林分蓄积量增加了71.5 m3/hm2;而林龄从40增加到50年,林分蓄积量增加了167.4 m3/hm2。由此可以看出,这一地区高山松林林地生产潜力巨大,蓄积量仍然保持着很高的增长量。
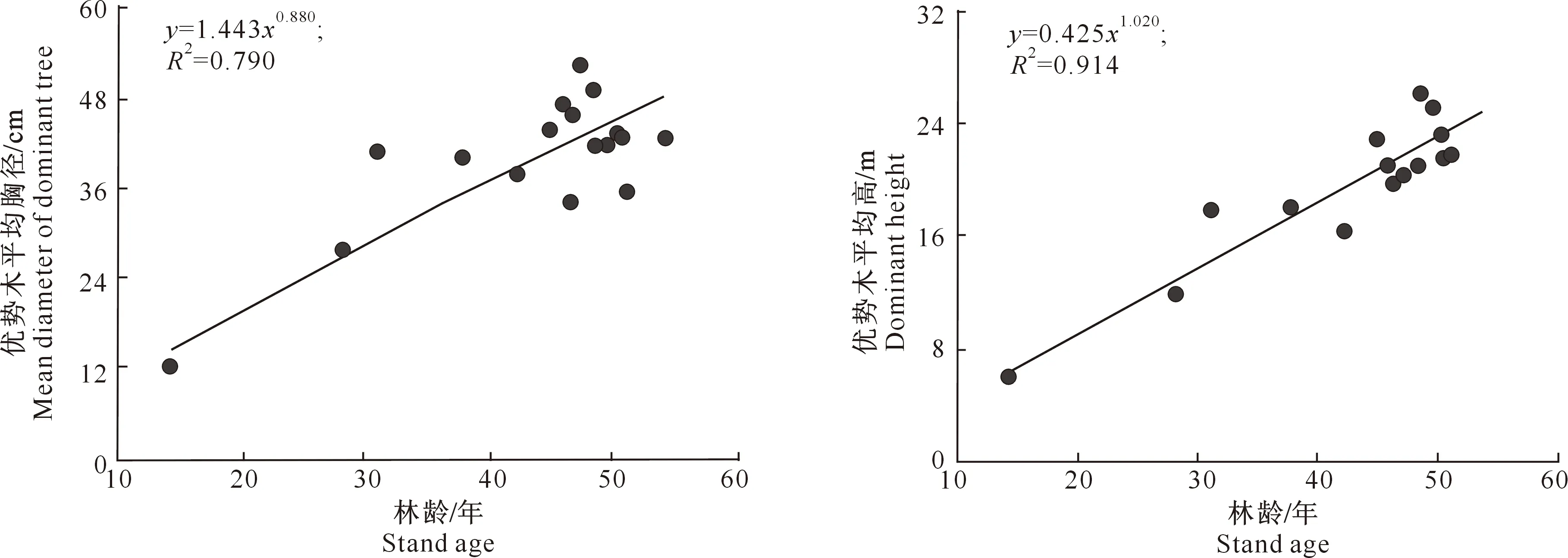
图4 尼洋河流域高山松次生林优势木平均胸径和平均高与林龄的关系
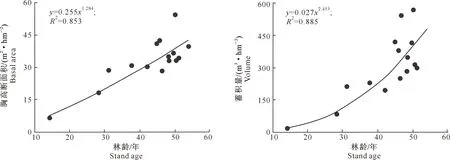
图5 尼洋河流域高山松次生林林分胸高断面积和蓄积量与林龄的关系
3.2.5 林下幼苗更新 高山松林林下更新幼苗密度与林龄的关系如图6所示,更新幼苗高度的数量分布情况见图7。
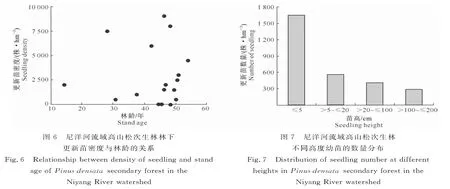
由图6可见,高山松林下以幼苗更新为主,不同样地间幼苗更新数量差异很大,每公顷更新苗密度最大为9 000株,最小为0株,平均为(2 912±2 986) 株,幼苗更新数量与林龄之间并无显著相关性。由图7可见,高山松林下不同高度更新幼苗的数量分布呈反“J”型。高度≤5 cm的1~2年生幼苗数量为1 647株/hm2,占总更新苗数量的56.6%;随着苗高的增加,更新苗数量逐渐减少,高度在100~200 cm的更新苗数量为294株/hm2,只占更新苗数量的10.1%。
4 结论与讨论
胸径结构反映林分内树木胸径的分配状态,将对林木的树高、干形、材积和树冠等因子产生直接影响。研究不同阶段的胸径分布规律可以掌握林分的生长状况,进而了解其生态进程。有研究表明,纯林直径正态分布曲线的形状随着林龄的增加而变化,即幼龄林平均直径较小,偏度为左偏;随着林龄的增加,林分平均直径逐渐增大,偏度变小,林分直径分布逐渐接近于标准正态分布曲线[20]。从本研究高山松林分的胸径分布来看,幼龄林的胸径分布呈现出偏左的近似正态分布;中林龄呈现出中间高两边低的近似正态分布,分布曲线的偏度和峰度均变小。在小径阶林木中偶尔有其他伴生树种,而在中大径阶林木中,高山松为主要优势树种,并无其他树种出现。左政等[21]研究发现,云南香格里拉地区成、过熟高山松林的直径结构为反“J”型的异龄结构,但中林龄呈偏左的正态分布,这与本研究结果一致。因此,尼洋河流域高山松次生纯林属于优势种群,且种群稳定,但树种较单一,尤其在中龄林内,这将影响林分生态功能的充分发挥。因此,建议在保持高山松天然次生林优势树种生长的基础上,还应促进其他阔叶树种的发展,提高其混交度。
本研究中,从林分因子与林龄的关系来看,高山松林分因子与林龄之间存在显著的相关性。林分密度随着林龄的增加呈下降趋势,当林分进入中龄林阶段,林分密度保持在(600±152.6) 株/hm2。林分胸径和林高与林龄之间以幂函数方式增长,且以树高与林龄的拟合精度更高。林木胸径的增长除受到树木年龄影响外,过高的林分密度对林木胸径生长也有显著影响[22],尤其是在林木出现竞争的中幼龄林内。树高主要由树木的生物学特性和树龄所决定,受林分密度的影响较小。本研究结果也表明,高山松林木胸径随林龄增加以幂函数方式增长且变异程度较大,除受到环境因子、生物因子、林分密度的影响外,也可能与研究区域高山松林分整体林龄较低有关。林分胸高断面积和蓄积量是林木胸径、树高、林木密度综合作用的结果,其与林龄之间的回归关系可用幂函数方程很好地拟合,决定系数R2均达到0.85以上。戎建涛等[23-24]基于森林资源二类调查中1 000多个不同林龄小班森林资源数据,发现东北天然次生针叶林的蓄积量与林龄之间的关系可用Schumache方程很好地拟合,而天然针阔混交林蓄积量与林龄之间为Richards函数关系,但蓄积量均随林龄的增长而呈现出增加趋势。本研究中,高山松林的林龄为18~54年,为中幼龄林,因此在这一年龄段内林分蓄积量与林龄间表现为幂函数增长。值得关注的是,本研究发现尼洋河流域高山松林分平均蓄积量为295.4 m3/hm2,当林分进入中龄林阶段(40~50年)蓄积量可以达到397.1 m3/hm2,远高于我国森林的平均蓄积量(89.79 m3/hm2)[25],也高于周彬等[26]报道的西南地区天然林蓄积量(123.0 m3/hm2),表明藏东南尼洋河流域高山松林整体质量高,生产潜力大,在藏东南地区发挥着重要的碳汇作用和生态作用。
高山松主要靠种子更新。此次研究表明,不同林龄高山松林下幼苗更新数量差异很大,更新苗密度在0~9 000株/hm2,平均为2 912株/hm2。幼苗更新数量与林龄之间并无显著相关性,各林龄基本一致、林分密度相似的林下更新苗数量也相差很大。通常情况下,幼苗数量会随着林龄的增加而增多,但是许多研究也表明,更新苗的数量与诸多因素有关,如地形[9]、地被物[27]、林分密度及林冠层郁闭度[28]等。另外,林内树木的分布情况,尤其是林隙的大小等均直接影响着林内光照情况,进而影响幼苗的生长发育[29]。高山松纯林内,随着林龄的增加地表枯落物明显增多,较厚的枯落物和阳坡、半阳坡等较干燥的地形环境阻碍了高山松种子的萌发和生长,可能是造成更新苗数量与林龄相关性不显著的原因。因此,还需要更深入地研究高山松天然更新的影响因素。另外,从更新幼苗的数量来看,其分布随幼苗高度的增大呈反“J”型,高度≤5 cm的1~2年生幼苗更新数量为1 647株/hm2,占总更新苗数量的一半(56.6%)以上;随着幼苗高度的增加,更新苗数量逐渐减少,高度在100~200 cm 的更新苗数量为294株/hm2,只占更新苗数量的10.1%,表明更新苗在生长过程中数量急剧减少。高山松虽属强阳性树种,但在其生长初期(1~3年),需要适当的庇荫,周围温度、湿度变幅太大会对其生长产生一定的影响,特别是对幼树幼苗的生长影响极大[9]。陈端[8]研究发现,高山松次生林林冠下的天然更新在林冠层郁闭度为0.6时更新较好,林下草本层和杂灌数量也对高山松幼苗的天然更新有很大影响,故不能寄希望于林冠下的天然更新来恢复第3代森林。因此,从提高林下天然更新幼苗的数量来看,可在郁闭度过大(0.7以上)的高山松纯林内进行间伐和择伐,以加速林木的生长和天然更新。另外,林窗的存在会改变群落物种组成,提高群落物种多样性水平,并且大林窗更有利于林下幼苗生长[28,30],可以通过开辟林窗等抚育措施来促进高山松次生林的天然更新。
猜你喜欢
Plasma Science and Technology(2022年9期)2022-08-29
防护林科技(2022年5期)2022-08-22
防护林科技(2020年10期)2020-12-22
四川林业科技(2020年1期)2020-08-31
绿色科技(2017年16期)2017-09-22
现代农业科技(2017年12期)2017-07-29
连环画报(2016年10期)2016-12-16
心潮诗词评论(2016年1期)2016-12-01
乡村地理(2016年2期)2016-06-15
林业与生态(2016年2期)2016-02-27
