
大熊猫转录因子Klf4在间充质干细胞中的转录调控作用研究
2019-11-22 07:39吴宏娟刘玉良梁雨桐沈富军侯蓉彭锐
四川动物 2019年6期
吴宏娟,刘玉良,梁雨桐,沈富军,侯蓉,彭锐*
(1.四川大学生命科学学院,生物资源与生态环境教育部重点实验室,成都610065; 2.成都大熊猫繁育研究基地,四川省濒危野生动物保护生物学重点实验室,四川省大熊猫科学研究院,成都610081)
大熊猫Ailuropodamelanoleuca是我国特有的濒危物种之一。由于栖息地破坏、遗传物质流失、自身繁殖率低下等一系列问题,大熊猫的生存一直受到威胁(Shenetal.,2010;李静,2017;Kang &Li,2018)。近年来,大熊猫的生理、行为生态学和再生生物学取得了很大的进步,但由于研究材料受限,对大熊猫基因功能的研究相对较少,这也限制了对大熊猫基因调控网络的深入认识(Zhangetal.,2008;Shenetal.,2010;Weietal.,2015;Kangetal.,2017)。
间充质干细胞(mesenchymal stem cells,MSCs)是一种具有自我更新和多向分化潜能的成体干细胞。由于这一特性,MSCs在再生医学和组织工程中备受关注(Wangetal.,2012)。Liu等(2013)已成功从大熊猫骨髓中分离出了MSCs,这为大熊猫细胞水平及细胞中不同生物进程的研究提供了原材料。但是关于大熊猫MSCs的细胞周期、生长、分化等生物进程的调控机制仍不清楚。
Krüppel样因子4(Krüppel-like factor 4,KLF4)是KLF家族中的一员(Schuhetal.,1986),是一个重要的锌指转录因子。KLF4的C端含有3个C2H2锌指结构,通过这3个锌指结构与靶DNA或蛋白结合,并调节其转录(Ghaleb &Yang,2017;Yangetal.,2017)。已有研究表明,KLF4在细胞生长、分化和正常组织稳态维持等细胞过程中发挥重要作用(Zhangetal.,2013;Shatatetal.,2014;Mallipattuetal.,2016;Ghaleb &Yang,2017)。同时,KLF4在诱导多能干细胞中也发挥着重要的作用(Takashietal.,2008)。Li等(2010)完成了大熊猫参考基因组的测序并公布了该基因组,但是,到目前为止,大熊猫Klf4基因的序列尚未得到实验验证。研究KLF4的特性和转录调控功能将有助于我们进一步了解大熊猫MSCs的增殖、分化及凋亡的分子机制。
为了进一步探究大熊猫Klf4基因的结构和功能,本文通过cDNA末端快速扩增(rapid amplification of cDNA ends,RACE)技术克隆获得了大熊猫Klf4基因的cDNA全长序列,并分析了其结构特征。此外,本文还通过病毒感染的方法实现在大熊猫MSCs中过表达KLF4,并通过RNA-seq方法进一步探索了KLF4在大熊猫MSCs中的功能。
1 材料与方法
1.1 实验材料
大熊猫MSCs以及睾丸、大脑、心脏、肝脏、脾脏、肺、肾脏组织由成都大熊猫繁育研究基地提供。实验所用EscherichiacoliDH5α感受态细胞购自成都诺维赞生物科技有限公司,慢病毒载体pLKO.3G、psPAX2以及pMD2.G购自Addgene。
1.2 实验试剂
细胞培养基购于美国HyClone;胎牛血清、青霉素/链霉素混合物、细胞生长因子购于美国Invitrogen;β-actin抗体、FLAG抗体购于成都正能生物有限公司;二抗(山羊抗鼠抗体、山羊抗兔抗体)购于美国Proteintech。RNAiso Plus、反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser、SMARTTMRACE cDNA Amplification Kit购于日本TaKaRa;质粒小量提取试剂盒购于天根生化试剂(北京)有限公司;琼脂糖凝胶DNA回收试剂盒购于成都擎科梓熙生物技术有限公司;ClonExpress Ultra One Step Cloning Kit购于南京诺维赞生物科技有限公司。
1.3 实验方法
1.3.1 大熊猫MSCs的培养大熊猫MSCs生长所需的低糖培养基包含10%胎牛血清、1%青霉素/链霉素、10 ng·mL-1表皮生长因子(Invitrogen)、5 ng·mL-1碱性成纤维细胞生长因子和7.5 mmol·mL-1谷氨酰胺-1(Invitrogen)。细胞在37 ℃,5%CO2浓度的细胞培养箱中培养。
1.3.2Klf4基因的克隆从大熊猫MSCs中提取RNA并根据SMARTTMRACE cDNA Amplification Kit的说明书反转录获得3’RACE和5’RACE的cDNA模板,然后通过基因特异性引物3’outer-KF、3’inner-KF和5’outer-KF,5’inner-KF与RACE通用引物进行巢式PCR(引物序列见表1)。PCR程序如下:94 ℃ 5 min;98 ℃ 30 s,70~60 ℃(每次循环递减0.5 ℃),72 ℃ 1 min,20个循环;98 ℃ 30 s,57~65 ℃温度梯度30 s,72 ℃ 1 min,10个循环;72 ℃ 5 min。获得Klf4基因的3’末端和5’末端产物,经纯化后进行TA克隆并测序。
1.3.3Klf4基因的序列分析将Klf4基因的3’末端和5’末端产物拼接后通过NCBI BLAST在线程序进行核苷酸和氨基酸序列同源性分析;用SWISS-MODEL对KLF4的氨基酸序列特点进行分析,并通过I-TASSER预测其蛋白质三维结构(Zhang,2008;Ambrishetal.,2010;Yangetal.,2015);通过MEGA 7.0构建系统进化树以分析大熊猫Klf4基因与其他物种的亲缘关系,基于KLF4的蛋白编码区(coding sequence,CDS)的核苷酸序列,采用邻接法,bootstrap重复1 000次计算各分支的置信度,各物种GenBank登录号为:人Homosapiens(NM_001314052.1)、家鼠Musmusculus(NM_010637.3)、狗Canislupusfamiliaris(XM_005626996.2)、黑猩猩Pantroglodytes(XM_01696 1392.2)、绿猴Chlorocebussabaeus(XM_0079684 89.1)、牛Bostaurus(NM_001105385.1)、猕猴Macacamulatta(NM_001142793.2)、野猪Susscrofa(NM_001031782.2)、非洲爪蟾Xenopustropicalis(NM_001017280.2)、家鸡Gallusgallusdomesticus(XM_00 4949369.3)、兔Oryctolaguscuniculus(XM_017347 259.1)、猫Feliscatus(NM_001173444.3)、褐家鼠Rattusnorvegicus(NM_053713.1)、羊Ovisaries(NM_001164219.1)、北极熊Ursusmaritimus(XM_008708 038.1)(Kumaretal.,2016)。
1.3.4Klf4基因过表达载体的构建以获得的3’RACE cDNA为模板,通过引物Full-length-Fw和Full-length-Rv经PCR扩增获得Klf4基因的CDS序列。以此为模板,再次通过PCR扩增的方法在KLF4 CDS的N端和C端分别加上1×FLAG标签和自剪切肽P2A序列。将获得的融合片段通过同源重组的方法克隆到酶切后的pLKO.3G载体中,组成慢病毒重组载体pLKO.Klf4。

表1 Klf4克隆及载体构建引物序列Table 1 Primer sequences used for Klf4 cloning and plasmid construction
1.3.5 KLF4重组慢病毒的产生及目的细胞的感染在293T细胞密度达到80%~90%时,通过脂质体将10 μg目的质粒、7.5 μg psPAX2以及2.5 μg pMD2.G转染入293T细胞中。转染后48 h和72 h时,分别收集病毒上清。病毒经浓缩后感染大熊猫MSCs。每8 h感染一次,共感染3次。
1.3.6 实时荧光定量PCR(qPCR)和Western blot分析取大熊猫睾丸、大脑、心脏、肝脏、脾脏、肺、肾脏组织,放入液氮中研磨,按照Trizol法提取总RNA,后用反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser分别将1 μg RNA反转录成cDNA。通过qPCR来检测目的基因及Klf4基因在大熊猫不同组织中的转录水平表达情况。qPCR(引物序列见表2)的反应程序如下:95 ℃ 5 min;95 ℃ 15 s,60 ℃ 30 s,40个循环。采用2-ΔΔCt的方法进行计算、分析结果。
在大熊猫MSCs中过表达了KLF4后,通过Western blot来检测KLF4的表达情况。收集感染pLKO.Klf4和pLKO.3G的细胞样品,经裂解液裂解变性后,用12%SDS-PAGE蛋白胶进行电泳,后经转膜、室温封闭后于4 ℃进行一抗过夜孵育。隔天,3次洗膜后,在室温下进行二抗孵育,1 h。洗膜后,通过蛋白质凝胶成像系统检测目的蛋白条带。

表2 qPCR所用引物Table 2 Primer sequences used for qPCR
1.3.7 RNA-Seq收集过表达KLF4的大熊猫MSCs和对照组pLKO.3G MSCs样品,由北京百迈克生物技术有限公司完成转录组测序分析工作。cDNA文库测序工作在Illumina Hiseq Xten平台上进行,使用FASTQC进行质量监控,获得了高质量的clean。通过Tophat2将这些clean与参考基因组序列比对,比对上的reads利用String Tie进行组装。然后使用EBseq进行差异分析,将满足筛选条件(错误发现率<0.01和差异倍数>2)的差异表达基因进行KEGG富集分析。
2 结果
2.1 Klf4基因的克隆与序列分析
大熊猫Klf4基因cDNA的全长为2 431 bp,其中,5’UTR长60 bp,3’UTR长934 bp,CDS长1 437 bp,编码479个氨基酸,分子质量约为51.9 kDa(GenBank登录号:MK075241)。Klf4基因的poly A尾上游18 bp处有1个多聚腺苷酸化信号(AATAAA)(图1:A)。KLF4蛋白的N端没有发现信号肽,C端则包含3个保守的C2H2锌指结构,分别位于氨基酸395~424、425~454、455~478处。此外,KLF4中还存在1个富含丝氨酸的结构域(图1:B)。KLF4的三级结构包含3个α-螺旋,每个α-螺旋中分布着1个锌指结构(图1:C)。系统发育树表明大熊猫Klf4基因与北极熊Ursusmaritimus的亲缘关系最近(图2)。
2.2 Klf4基因在大熊猫不同组织中的表达分析
为探索KLF4在大熊猫不同组织中的功能,通过qPCR的方法检测Klf4基因在大熊猫不同组织中的表达分布。结果发现,Klf4基因在睾丸中的表达量最高,其次是肺、脾脏、肾脏,而大脑、肝脏和心脏中的表达量相对较低(图3)。
2.3 KLF4对大熊猫MSCs的调控
为了进一步了解KLF4在大熊猫骨髓MSCs中的作用,通过慢病毒感染的方法过表达了KLF4(图4:A),并在转录组水平上分析其调控作用。转录组分析发现KLF4过表达后,大熊猫MSCs出现了179个差异表达基因,包括79个上调基因和100个下调基因(图4:B),这些差异表达基因的层次聚类分析结果见图4:C。对这些差异基因进行KEGG富集分析发现,差异性表达基因显著富集在军团杆菌病、Hippo信号通路、RNA转运、TNF信号通路以及咖啡因代谢等通路(图4:D)。
2.4 RNA-Seq结果的验证
为了验证转录组数据的可靠性,通过qPCR分别对上调基因MYL6B、TMC8、TMEM200B、SLAIN1、SOCS3以及下调基因CASP10、P2RY10的表达情况进行了验证(图5:A)。同时,还有一些与细胞周期调控相关基因(Cyclin D1和P21),以及与Klf4基因相互作用的Nestin、Pax6基因的表达情况(图5:B)。qPCR结果与转录组数据的结果保持一致,说明转录组数据可真实反映基因表达量的差异情况。

图1 大熊猫Klf4基因的序列及结构特点Fig.1 Gene sequence and predicted protein structural characteristics of Klf4 gene in Ailuropoda melanoleucaA.大熊猫Klf4 cDNA全长序列及对应氨基酸序列,起始密码子(ATG)和终止密码子(TAA)以粗体表示,推测的多聚腺苷酸化信号(AATAAA)以黄色阴影显示;B.KLF4蛋白的结构示意图,SER Rich代表丝氨酸富集区,ZF代表锌指结构域;C.预测的KLF4三级结构,包含3个α-螺旋,锌指结构分布在每个α-螺旋中A.The full-length of giant panda Klf4 cDNA and corresponding amino acid sequences,the initiation codon (ATG)and stop codon (TAA)are characterized in bold,and the putative polyadenylation signal (AATAAA)is displayed with yellow shadow;B.schematic features of KLF4 protein,SER Rich represents serine enrichment region,ZF represents zinc finger domains;C.predicted tertiary structure of KLF4 containing 3 α-helices with a zinc finger
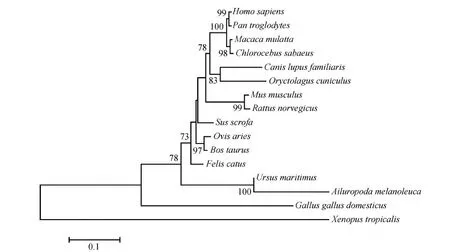
图2 采用邻接法构建不同物种KLF4的系统发育树Fig.2 The phylogenetic tree of KLF4 from various species constructed using the neighbor-joining method
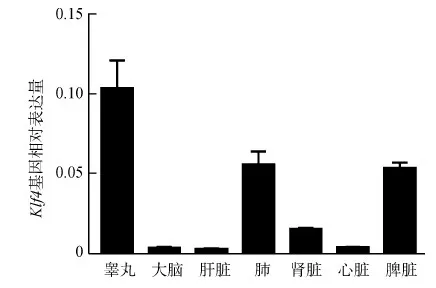
图3 Klf4基因在大熊猫不同组织中的表达Fig.3 Relative expression levels of Klf4 gene in different tissues of Ailuropoda melanoleuca

图4 KLF4对大熊猫间充质干细胞的转录调控作用Fig.4 The regulatory function of KLF4 in Ailuropoda melanoleuca mesenchymal stem cells (MSCs)A.通过qPCR和Western blot检测对照组(pLKO.3G)和KLF4过表达组(pLKO.Klf4)MSCs中KLF4的mRNA和蛋白表达水平;B.对照组和KLF4过表达组MSCs中差异表达基因(DEGs)的MA图,红、绿点分别代表上调、下调DEGs,黑色代表未发生改变的基因;C.对照组和KLF4过表达组MSCs中DEGs的聚类分析热图;D.对照组和KLF4过表达组MSCs中DEGs前20显著富集的KEGG通路;* P<0.05,** P<0.01A.MSCs were infected with control lentivirus (pLKO.3G)and KLF4 overexpression lentivirus (pLKO.Klf4),and then the mRNA and protein expression levels of KLF4 were measured by qPCR and Western blot;B.MA Plot of differentially expressed genes (DEGs)between pLKO.3G MSCs vs.pLKO.Klf4 MSCs;red,green and black dots represent up-regulated DEGs,down-regulated DEGs and unchanged genes,respectively;C.Heatmap clustering of DEGs between pLKO.3G MSCs vs.pLKO.Klf4 MSCs based on overall expression similarity;D.top 20 significantly enriched KEGG pathways of the DEGs between pLKO.3G MSCs vs.pLKO.Klf4 MSCs based on P-value;*P<0.05,** P<0.01

图5 对照组(pLKO.3G)和KLF4过表达组(pLKO.Klf4)中上、下调DEGs的qPCR验证Fig.5 qPCR verification of up- and down-regulated DEGs between pLKO.3G vs.pLKO.Klf4A.重要的DEGs表达量的qPCR验证;B.细胞周期调控相关基因及与KLF4互作的基因表达量的qPCR验证;* P<0.05,** P<0.01A.qPCR was employed to verify the expression of some important DEGs;B.the genes associated with cell cycle and KLF4;*P<0.05,** P<0.01
3 讨论
本研究从大熊猫MSCs中克隆获得了大熊猫Klf4基因cDNA全长,共2 431 bp,编码479个氨基酸。大熊猫KLF4的N端比人、鼠的少9个氨基酸残基,而C端具有KLF家族高度保守的C2H2结构,该结构介导KLF4与靶DNA或蛋白的结合。Klf4基因在大熊猫睾丸中的表达量最高,表明Klf4基因可能参与调控其中重要的生理过程,如精子的发生过程(Yangetal.,2017);此外,Klf4基因在脾脏、肾脏中的表达也较高,表明Klf4基因在脾脏、肾脏中可能同样也发挥着重要的调控作用,如KLF4会影响肾足状突细胞的表型和功能(Kaorietal.,2014)。KLF4在大熊猫不同组织中的分布差异可能是机体内稳态调控的结果。
基于转录组测序结果分析,在大熊猫MSCs中过表达KLF4后,差异表达基因最显著富集在军团杆菌病代谢通路。KLF4可能通过影响大熊猫MSCs中真核翻译延伸因子1α(eEF1A)的表达进而影响军团杆菌病代谢。eEF1A是一个多功能蛋白,在蛋白质的翻译延伸、细胞凋亡、细胞信号转导以及心血管疾病发生等过程中发挥重要作用(Khachoetal.,2008)。KLF4调控eEF1A1和eEF1A3的表达发生改变,进而引起军团杆菌病代谢的改变。
KLF4可显著影响大熊猫MSCs中的RNA转运过程。转录组结果表明,KLF4可显著下调输出蛋白XPOT的表达。XPOT参与成熟tRNA从细胞核到细胞质的转运过程(Guptaetal.,2016)。XPOT的表达量减少,因此与它的锚定蛋白RanGTP的结合减少,导致tRNA转运过程中的能量供应不足,进而影响细胞核与细胞质中RNA的运输。
此外,在KLF4过表达后,肿瘤抑制基因LATS1的表达量减少。LATS1是Hippo信号通路的重要组成成分,已有研究表明,低表达的LATS1会抑制Hippo信号通路,进而影响小鼠MSCs的增殖、迁移和分化过程(Qiuetal.,2015)。类似的,在大熊猫MSCs中,KLF4可能通过下调LATS1的表达,抑制Hippo信号通路。
与之前的报道不同(Wassmannetal.,2007;Chenetal.,2011),在大熊猫MSCs中,KLF4可通过上调细胞周期蛋白cyclin D1同时下调细胞周期激酶抑制基因P21的表达来促进细胞增殖。同时,KLF4可能通过影响细胞凋亡相关基因CASP10和TMC8的表达而参与调控大熊猫MSCs的凋亡过程。
本研究结果进一步揭示KLF4在大熊猫MSCs中发挥着重要的调控作用,后续可深入研究其具体的调控机制。本文扩展了大熊猫分子水平的相关研究,为今后的相关研究提供了理论依据,为大熊猫遗传资源的保护和利用提供了更多的思路。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
中国农学通报(2022年12期)2022-06-01
理财周刊(2022年4期)2022-04-30
昆明医科大学学报(2022年1期)2022-02-28
中国种业(2021年11期)2021-11-25
小哥白尼(野生动物)(2021年7期)2021-11-20
幼儿教育·父母孩子版(2020年8期)2020-03-04
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
