
HGF/c-Met信号通路调控生殖细胞发生的研究进展
2019-12-04 02:03李康梅陈秀榕戴明明黎明星黄愉淋白琳吴媛媛何国珍
实用医学杂志 2019年21期
李康梅 陈秀榕 戴明明 黎明星 黄愉淋 白琳 吴媛媛 何国珍
1广西中医药大学基础医学院(南宁530200);2广西中医药大学第一附属医院(南宁530023);3广西中医药大学第二附属医院(南宁530001)
卵巢的卵泡由卵母细胞、颗粒细胞及膜细胞组成,睾丸由生精小管和生精小管之间的睾丸间质组成,支持细胞、睾丸间质细胞及肌样细胞为其主要生殖细胞。肝细胞生长因子(hepatocyte growth factor,HGF)为多肽生长因子,肝细胞生长因子受体(HGF-receptor,也称c-Met)为HGF 唯一的受体,二者通过特异性结合,可调控性腺发育以及生殖细胞的增殖,促进胎盘滋养细胞、血管内皮细胞、肝细胞等多种细胞的增殖以及侵袭。本综述主要对HGF/c-Met 信号通号通路调控生殖细胞发生的研究进展进行概括。
1 HGF/c-Met 信号通路
HGF 最早从大鼠的血浆中发现,主要源自肝脏的储脂细胞,可强烈刺激肝细胞DNA 的合成,后又因能刺激原代培养肝细胞的生长与分化,被称为HGF[1-2]。HGF 为多肽生长因子,前体为单链无活性蛋白,由一条728 个氨基酸编码而成,经丝氨酸蛋白水解酶水解,再由二硫键连接,最终形成具备活性的双链分子(由α-β链组成的一个异质二聚体)[3-4](图1)。α链较长,含有一个典型的信号肽,在分泌过程中裂解形成5 个结构域:与纤溶酶原激活肽同源的一个N 端发夹环结构域(N)与4 个环状结构域(K);β链较短,与典型的丝氨酸蛋白酶结构域相似[5]。
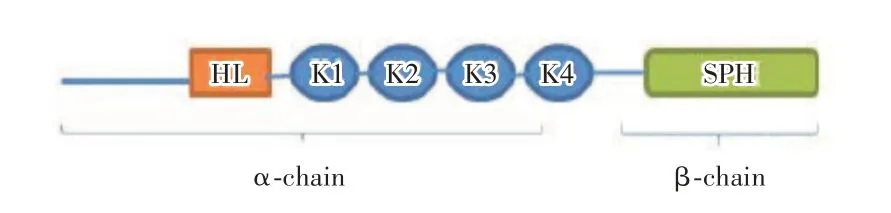
图1 HGF 的结构图[6]Fig.1 Molecular structure of the HGF[6]
c-Met 是受体酪氨酸激酶的独特亚家族,由原癌基因MET 编码产生,具备自主磷酸化活性;主链前体蛋白为Ⅱ型酪氨酸激酶受体,以二硫键结合α 和β 两个亚基所组成单链构成主要结构,包含细胞外配体结合区、细胞内胞质蛋白激酶结构区以及跨膜螺旋结构。c-Met 蛋白的细胞外区域(图2)由3 个功能区域组成:包含所有α 亚基和β 亚基N端部分β 螺旋折叠区域(Scma domain)、含4 个二硫键的富胱氨酸结构域(plcxins,scmaphorins and intcgrins domain)、与跨膜螺旋区相连接的4 个免疫球蛋白样结构域(immunoglobulin-plcxin-transcription domain);β 亚基的细胞内区具有酪氨酸激酶活性并能转换HGF 的所有效应。c-Met 蛋白的细胞内区则由近膜结构区、激酶催化结构区及C-末端结构区组成,近膜结构域有2 个磷酸化位点,参与c-Met 表达的下调和内化;酪氨酸激酶催化位(Tyr1235、Tyr1234 以及Tyr1230)通过磷酸化能正向调控酶的活性;C-末端结构域包含一个独特的结合位点,其中的Tyr1349 与Tyr1356 磷酸化能吸引多种信号转导蛋白和适配蛋白,进而激活多条下游信号通路[4,7]。
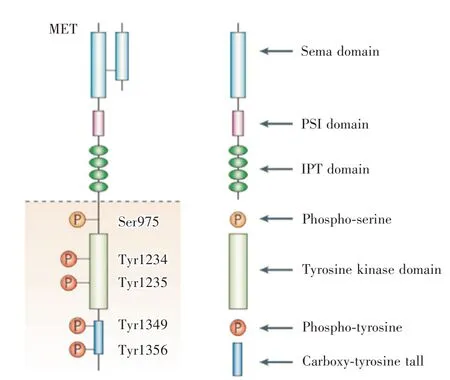
图2 c-Met 结构图[8]Fig.2 Molecular structure of the c-Met[8]
HGF 与c-Met 通过两个激酶位点产生特异性结合,一个是经过泛素连接酶(casitas B-lineage lymphoma,CBL)与酪氨酸激酶催化区(tyrosine kinase domain;consist of tyr 1234 and tyr1235,TKCD)结合,分泌泛素化酶,激活HGF-c-Met-CBL 通路的负调节作用,抑制c-Met 及其他下游受体酪氨酸激酶的活性[4,9];另一个则利用有SH2 结构域的多效应物质与位于Met 尾端的多功能激酶位点相结合,所形成的c-Met 复合物,再与如磷脂酰肌醇3-激酶(phosphatidyl inositol 3-kinase,PI3K)、IκBα-NF-κB 复合物、生长因子受体结合蛋白-2(Grb2-associated binder2,Gab2)等直接或间接结合,启动相应的信号通路,发挥细胞的增殖、侵袭能力等作用[4,10]。c-Met 与HGF 结合后,酪氨酸激酶活性被激活,产生二聚化,c-Met 胞质内酪氨酸残基发生自身磷酸化,胞质内的蛋白激酶结构区的酪氨酸激酶被激活,为下游信号传导底物(Src、Gabl、Gab2)提供停泊位点,激活如PI3K/丝氨酸/苏氨酸蛋白激酶(serine/threoninespecific protein kinase,AKT),Janus 激酶(Janus kinase,JAK)/信号转导和转录激活因子3(signal transducer and activator of transcription 3,STAT3),肾素-血管紧张素系统(renin-angiotensin system,Ras)/丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)等信号通路(图3)[11-12],并把增殖信号传递至细胞核,调控特定基因的表达,促进细胞的分裂增殖、分化、迁移、形态发生、蛋白质合成以及血管生成等[13]。HGF 表达于周围间质细胞,c-Met 则多表达于上皮来源细胞[3,7],对胎盘滋养细胞、卵泡发生相关细胞(如卵泡膜细胞、颗粒细胞等)、血管内皮细胞、肝细胞等多种细胞增殖有调节作用。在胚胎期与成人期均可在多个器官如肝脏、胰腺、前列腺中检测出c-Met,其表达对胎盘、肝、肾、脑等的正常发育及造血功能有重要影响[8]。
2 HGF/c-Met 信号通路对卵泡发生的调控
卵泡作为哺乳动物卵巢组织的基本功能单位,包含有膜细胞、上皮衍生的颗粒细胞以及周围的基质细胞所围绕的卵母细胞。卵泡发生是一个连续数周的进程,相继经历原始卵泡、初级卵泡、三级卵泡和成熟卵泡几个阶段。始于原始卵泡被激活,后募集一个或多个原始卵泡到生长卵泡库,细胞增大并增殖,最终止于排卵或者卵泡闭锁。通常将该进程划分为促性腺激素不依赖阶段与促性腺激素依赖阶段。
卵泡发生有着精密的时序调控。优势卵泡排卵后,卵泡破裂排出后,卵巢表面上皮的损伤以及残留在卵巢内的部分分化成为黄体。整个卵泡发生的过程实际上是一个持续进行的包含血管发生、损伤修复以及衰退的组织生长过程。HGF/c-Met 信号通路可能直接作用于血管内皮细胞调节血管形成、提高毛细血管密度及血流速度来影响损伤的修复,并影响胚胎的发育[14-15]。虽然哺乳动物卵巢的颗粒细胞不含血管,但膜细胞层外有丰富的血管,膜细胞外的血管发育与卵泡发育有较大联系。此外,HGF 与c-Met 还在多种动物如大鼠、猪、牛以及人类的前列腺、睾丸、子宫内膜、膜细胞、颗粒细胞内有不同程度表达,HGF 可通过自/旁分泌调节膜细胞和颗粒细胞的增殖,HGF 在卵巢的表达与妊娠率低、女性性腺疾病发病相关,还可能影响卵泡发生[14,16]。
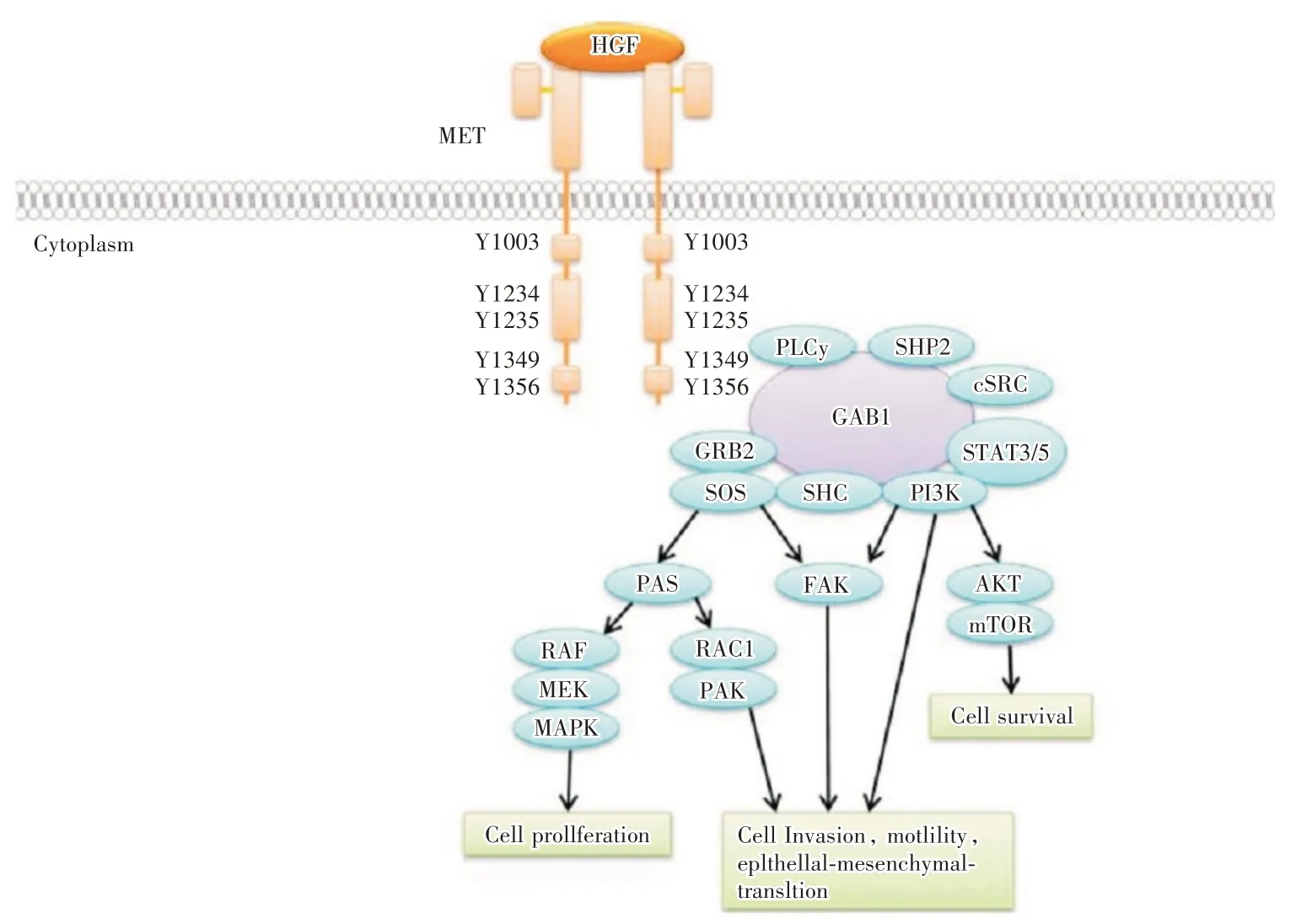
图3 HGF/c-Met 信号通路图[8]Fig.3 Molecular of the HGF/c-met signaling pathway
2.1 HGF/cMet 信号通路与卵泡膜细胞研究发现HGF 与c-Met 在人卵泡膜细胞和颗粒细胞上表达,HGF 可能直接通过改变类固醇形成基因近而调节膜细胞雄激素和孕酮的分泌[17]。在卵泡发生的早期启动中,HGF 可直接作用于膜细胞,但对膜细胞分泌类固醇激素调节是间接的[18-19]。将绒毛膜促性腺激素作用于未成熟大鼠,可提高HGF 与c-Met 在囊状卵泡膜细胞上的表达[5]。HGF 还可促进生殖细胞分化以及由间质细胞衍生的膜细胞分泌,在卵泡发育期间刺激上皮颗粒细胞增殖[16]。
2.2 HGF/c-Met 信号通路与颗粒细胞颗粒细胞可通过分泌多种生长因子、信号转导因子、旁分泌因子等来调控卵母细胞的成熟。最早报道HGF 在卵巢中的效应是其可诱导体外培养的牛颗粒细胞的增殖,并且有研究发现HGF 及其受体c-Met 的mRNA 在人排卵前卵泡颗粧细胞中表达[3]。HGF与c-Met 在小鼠早期卵巢就有表达,随年龄增大c-Met 表达逐渐增强,HGF 增强不明显,与干细胞因子/KIT 配体(KIT ligand)相互作用可促进颗粒细胞的增殖和早期卵泡发生[17]。 HGF 可正向调控颗粒细胞增殖与分化,抑制卵泡的闭锁,并可表达在不成熟的大鼠卵巢的窦前卵泡间质膜细胞和颗粒细胞中,促进窦前卵泡间质膜细胞分泌孕酮,但抑制了雄激素的产生,表明HGF 可逆性的抑制由黄体生成激素(luteinizing hormone,LH)刺激的雄激素的产生从而促进卵泡发生[19-20]。
2.3 HGF/cMet 信号通路与卵泡发生机制卵泡发生过程伴随着卵巢颗粒细胞和膜细胞的增殖和分化,产生各种卵巢局部生长因子,建立并维持卵母细胞成熟所需要的局部微环境。HGF 为旁分泌调节因子,对间质细胞来源的膜细胞和上皮性的颗粒细胞的相互影响有重要的意义,HGF 和c-Met可同时促进膜细胞和颗粒细胞的发育[16]。在未成熟大鼠卵巢中,应用免疫组化发现c-Met 在小的腔前卵泡的颗粒细胞中表达最强,在用孕马血清作用诱导囊状卵泡的生长之后,HGF 与c-Met 的表达显著转变,在颗粒细胞的表达相对下降,但在囊状卵泡外的膜细胞的表达相对增高[18],表明HGF 与c-Met 不仅可表达在颗粒细胞和膜细胞,同时二者的表达水平受体内激素的影响。膜细胞所分泌的HGF、转化生长因子(transforming growth factoralpha,TGF)、角质细胞生长因子(keratinocyte growth factor)等因子。雌激素和促性腺激素可以刺激这些细胞因子的表达,表明激素和局部的旁分泌因子都是通过调节细胞之间的相互作用,从而调节或刺激原始卵泡的发生以及发育[16,21]。
ZACHOW 等[22]研究显示,卵泡刺激素(follicle stimulating hormone,FSH)和HGF 均可各自降低体外培养大鼠颗粒细胞上c-Met 的表达,相反,环磷酸腺苷(cyclic adenosine monophosphate)则可增加c-Met 的表达程度。c-Met 在体外受精成功的卵母细胞颗粒细胞的表达远比受精不成功的卵母细胞颗粒细胞要强,提示HGF/cMet 信号通路可能是卵母细胞成熟并能受精成功的一个重要因素[23]。
3 HGF/cMet 信号通路对精子发生的调控
精子发生根据细胞增殖和分化特征分为3 个时期:(1)有丝分裂期:原始生殖细胞多次有丝分裂分化成为精原细胞;(2)减数分裂期:精原细胞再分化形成精母细胞,精母细胞又通过减数分裂形成圆形精子细胞;(3)精子形成期:圆形精子细胞变态分化形成蝌蚪状的精子,此时的精子已经具有一定的运动能力。精子从生精小管上皮释放进入管腔,再进入附睾继续发育,最终发育为成熟精子。
精子发生的主要受激素和旁分泌因子两方面调节。一是通过下丘脑-垂体-睾丸轴来调节精子发生的启动和维持,下丘脑分泌促性腺激素释放激素,刺激垂体分泌的FSH 和LH,LH 又促使睾九间质细胞分泌的睾酮,最终调节精子的发生。二是通过激素受体和受体后信号转导,局部的旁分泌、自分泌激素以及生长因子(如HGF、FGFs 等)调节精子发生。
HGF 为多生物学效应的生长因子,能够促进细胞迁移、增殖以及血管生成[24-25]。HGF/c-Met信号通路在睾丸发育的所有阶段均有活跃的表达[22,26]。HGF 为自/旁分泌生长因子,可调节机体的内分泌状态,可直接作用于男性生殖细胞,或间接对睾丸管状细胞和间质体细胞产生作用。HGF 可促进胎儿和成人睾酮的分泌。在离体培养的支持细胞系中加入外源性HGF 能够培养出类管样组织,猜测这可能与HGF 促分化和促血管形成的功能有关[27]。在啮齿类动物中,胎儿的男性化和血浆雄激素(睾酮)的增加发生在下丘脑-垂体-睾丸轴发挥作用之前。转基因缺乏促黄体生成素受体小鼠在出生时仍呈现正常的睾丸表型雄激素水平,表明在下丘脑-垂体-睾丸轴产生正常的生理作用之前,胚胎雄性睾酮的增加以及胎儿睾丸间质细胞的分化,主要受局部生长因子的调控,HGF 极有可能参与其中[12,28]。
3.1 HGF/cMet 信号通路与睾丸间质细胞睾丸间质细胞能合成和分泌雄激素-睾酮,调控精子发生过程[29]。睾丸间质细胞的凋亡或消失会造成雄性激素能力低下[30]。在胚胎形态发生时,HGF 在上皮-间充质相互作用中发挥重要作用。HGF 可诱导间充质细胞转化,并使间质细胞分泌得睾酮量显著增加,降低睾丸间质细胞凋亡,可能与HGF抑制caspase-3 的活性有关[11,13,31]。此外,HGF 还可促使间质细胞分泌蛋白酶,提高TGF-β的活性,调控支持细胞之间紧密连接,影响血-睾屏障的形成[27]。在离体实验培养的睾丸间质细胞中,发现HGF 能刺激睾酮合成,促进胚胎睾丸间质细胞发育并延长睾丸间质细胞的存活[11,31]。
3.2 HGF/cMe 信号通路与肌样细胞肌样细胞围绕在生精小管周围,负责管状收缩,帮助运送非活动精子进入附睾,其发育依赖于雄激素作用,可分泌因子调控精子发生和睾丸的发育。肌样细胞系是发现HGF/c-Met 系统的第一个分离的睾丸细胞类型,HGF 在胚胎时期和出生后的肌样细胞中均有表达[22],但c-Met 仅在出生后的肌样细胞中检测到,表明在胚胎发育过程中有旁分泌作用,在出生后的睾丸中有旁分泌/自分泌作用[1-2]。文献[27]显示在HGF 的存在下培养的肌样细胞,形态从圆形的、致密的形状转变为拉长的、扩大的形状,HGF 还可激活睾丸肌样细胞骨架重构和收缩,提高纯化的睾丸肌样细胞尿激酶型纤溶酶原激活物和尿激酶型纤溶酶原激活物受体的表达水平,提示HGF 作为肌样细胞功能的自分泌调节因子,可能是哺乳动物睾丸功能的调控因子。
3.3 HGF/cMet 信号通路与精子发生机制c-Met广泛表达于人和啮齿动物的生精细胞[11]。HGF/c-Met 信号通路可通过影响内分泌信号,参与生精细胞凋亡的调控。c-Met 在大鼠精子发生全程中持续表达,离体实验表明,HGF 可调控生殖细胞有丝分裂,显著促进增加8 ~30 d 龄的大鼠睾丸精原细胞的增殖,并参与出生后睾丸生殖细胞的代谢活动的调节,调节睾丸生殖细胞的增殖并阻止生殖细胞凋亡过程[22]。精子发生在睾丸,成熟于附睾,检测HGF 在大鼠附睾中的表达水平,在远端的附睾体、附睾尾中表达最高,其次是近端的附睾体,但在附睾头、输精管以及睾丸中几乎不表达,提示哺乳动物附睾内精子获能可能与HGF相关[12,32]。
4 结语与展望
目前对于卵子发生和精子发生的调控机制的研究很多,但其机制仍然未阐明,特别是对于原始卵泡的启动募集机制以及精原细胞增殖启动机制的研究。综上所述,HGF/c-Met 信号通路可通过调控生殖细胞的增殖和分化调控卵子发生和精子发生,通过对这条信号通路的研究以及其他自/旁分泌因子研究,有望阐明原始卵泡启动募集和精原细胞启动机制,为不孕不育、多囊卵巢综合征以及卵巢早衰等性腺疾病以及其他非性腺疾病的发病机制和治疗提供理论依据。
猜你喜欢
中国畜牧杂志(2022年11期)2022-11-17
右江医学(2021年9期)2021-10-22
天津医科大学学报(2021年3期)2021-07-21
世界科学技术-中医药现代化(2021年12期)2021-04-19
昆明医科大学学报(2021年1期)2021-02-07
中国生育健康杂志(2020年3期)2020-05-15
基础教育论坛·上旬(2019年7期)2019-09-10
中国临床医学影像杂志(2019年6期)2019-08-27
中成药(2017年9期)2017-12-19
中国医药生物技术(2015年4期)2015-12-26
