
利用CRISPR/Cas9技术快速创制香型“秀水134”水稻
2019-12-13 08:26赵国超朱小方王彤李建粤
上海师范大学学报·自然科学版 2019年5期
赵国超 朱小方 王彤 李建粤


摘 要: 以目前上海市主栽的高产常规水稻“秀水134”为材料,利用CRISPR/Cas9技术成功敲除甜菜碱醛脱氢酶2基因,获得了两种类型纯合突变体植株.采用表达载体特异性结合的引物检测T1代转基因植株,成功获得6株不携带载体骨架的转基因植株.定量PCR分析显示,突变体植株甜菜碱醛脱氢酶2基因表达量极显著低于野生型对照(p<0.01),但突变体植株成熟种子香味物质2-乙酰-1-吡咯啉(2AP)含量极显著高于野生型对照(p<0.01).比较野生型对照与突变体植株的主要农艺性状和产量性状,两者间都没有显著差异(p>0.05).本研究可为加快高产香型水稻在上海及周边地区的推广应用,以及为今后利用CRISPR/Cas9技术快速培育其他高产香型水稻新品种研究奠定基础.
关键词: 水稻; 甜菜碱醛脱氢酶2基因; 2-乙酰-1-吡咯啉(2AP); CRISPR/Cas9基因编辑技术; “秀水134”
中图分类号: S 511 文献标志码: A 文章编号: 1000-5137(2019)05-0574-07
Abstract: In the study,fragrance rice mutant lines were developed with CRISPR/CAS9 by editing fragrance gene Badh2 in “Xiushui 134”,a major elite rice cultivar in Shanghai region.Six mutant lines without the vector backbone were successfully obtained by PCR detection.Quantitative PCR analysis showed that the expression of Badh2 in mutant plants was significantly lower than that in wild type (p<0.01),the content of 2AP in mature seed of mutant plants was significantly higher than that in wild type (p<0.01),but there were no significant differences on main agronomic traits between the mutant plants and wild type (p>0.05).This research can accelerate the application of high-yield fragrant rice in Shanghai areas,and lay the foundation for the future study on the rapid cultivation of other high-yield fragrant rice varieties using CRISPR/Cas9 technology.
Key words: rice; betaine aldehyde dehydrogenase 2 gene; 2AP; CRISPR/Cas9 gene editing technology; “Xiushui 134”
香味是稻米的重要食味品質性状之一.使用具有香味的米煮饭,稻米在蒸煮和食用过程中都会散发清香气味而深受消费者钟爱.由于市场对香米需求量的日益增加,国内外的香型水稻育种研究也越来越受到重视.
稻米香味由隐性基因控制,并位于第8号染色体上[1-3].BRADBURY等[4]以泰国香稻为材料研究发现:水稻香味的产生与第8号染色体上编码甜菜碱醛脱氢酶2(Betaine aldehyde dehydrogenase 2,BADH2)基因(Badh2)有关.SHI等[5]在不同香稻品种中发现Badh2编码链上不同位点的突变.CHEN等[6]和NIU等[7]分别通过Badh2基因互补实验和RNAi实验证实了Badh2就是水稻的香味基因.
文献[8-10]报道了利用分子标记辅助选育各种优质香型水稻的研究成果.尽管利用分子标记辅助育种技术可以有针对性地对水稻植株进行香味基因的追踪筛选,但其与传统常规育种所需时间相同,在本地和海南加代繁殖周期较长(3.5~4.0 a).
“秀水134”水稻是目前上海市主栽的高产常规水稻品种,表现为稳产,穗型较大,不易倒伏和高抗病性等特性[11],但其属于非香水稻.
本研究利用CRISPR/Cas9技术,以“秀水134”水稻为材料,对其香味基因Badh2进行编辑,获得了香型“秀水134”水稻,为加快高产香型水稻在上海及周边地区的推广应用,也为今后利用CRISPR/Cas9技术快速培育其他高产香型水稻新品种提供参考.
1 材料与方法
1.1 水稻材料和种植
以“秀水134”为遗传转化受体,开展常规的转基因实验,转基因材料种植于上海师范大学实验基地,常规田间种植及管理.
1.2 实验方法
1.2.1 Badh2敲除载体的构建
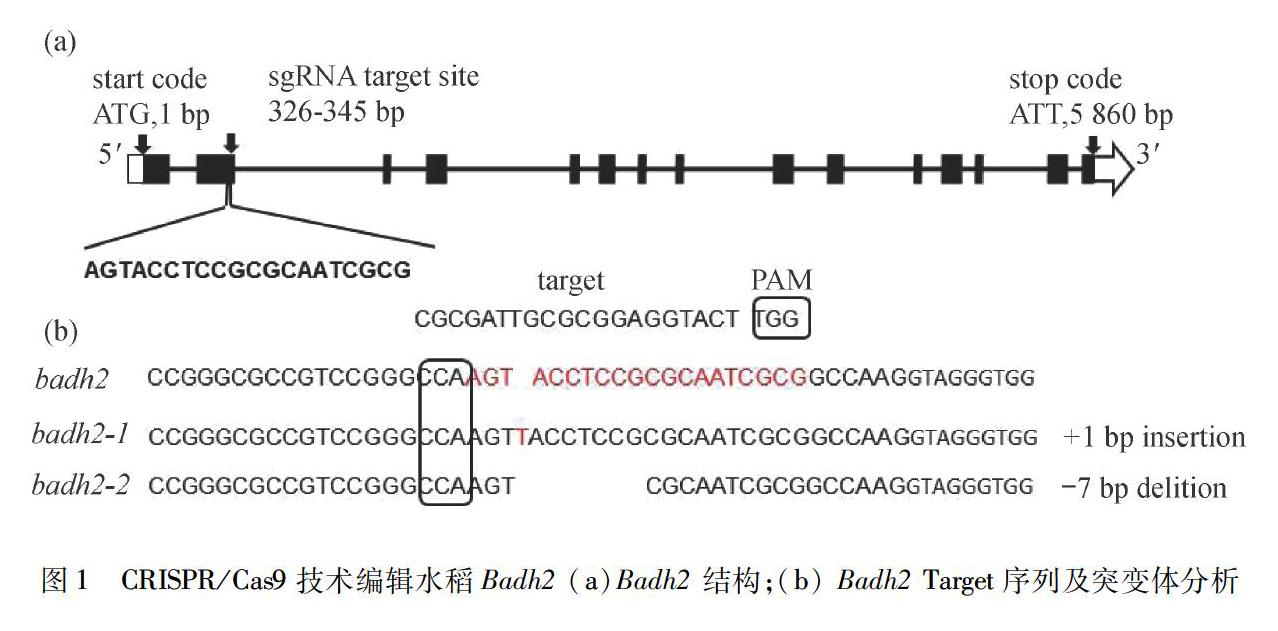
利用CRISPR-P网站设计Badh2基因(Xiushui 134Os08g0424500)一个Target序列(http://cbi.hzau.edu.cn/CRISPR/).Target序列为CGCGATTGCGCGGAGGTACT(对应基因互补序列:AGTACCTCCGCGCAATCGCG),如图1(a)所示.采用引物合成的方式获得所选的Target序列.引物合成时,在Target序列上游引物5′端加TGGC,下游引物5′端加AAAC.引物由上海生工生物工程公司合成.
按照常规分子生物学方法完成CH-Target中间载体和pCAMBIA1300-CH-Badh2敲除载体构建以及鉴定的工作.将pCAMBIA1300-CH-Badh2载体导入根癌农杆菌并完成鉴定(CH-Target中间载体由上海植物逆境生物学研究中心朱健康课题组提供).
1.2.2 根癌农杆菌介导转化水稻
分别取“秀水134”水稻幼穗或成熟胚诱导的愈伤组织为遗传转化外植体.根癌农杆菌介导水稻遗传转化中的培养基组成、愈伤组织诱导、转化操作、抗性筛选及植株再生方法都参照李建粤等[12]的报道.
1.2.3 转基因阳性植株和突变类型鉴定及无载体突变体植株的获得
设计一对能够与pCAMBIA1300-CH-Badh2载体T-DNA序列同源配对的引物F0/R0(表1),对转基因水稻叶片提取的基因组DNA进行扩增及电泳检测.
在目的基因靶点位置两侧设计引物F2a/R2a(表1),对转基因水稻阳性植株基因组DNA进行扩增.扩增产物送华大生物公司进行测序.测序结果利用DNAMAN软件进行分析,并与野生型Badh2基因序列进行比对.根据峰图和比对结果及网站(http://dsdecode.scgene.com/)判断突变类型.
利用F0/R0引物(表1),对转基因T1代植株叶片提取的基因组DNA进行PCR扩增及电泳检测.
1.2.4 突变体植株Badh2基因定量PCR检测
分别根据Actin和Badh2基因序列设计定量PCR检测引物(表1).分别收获“秀水134”和突变体水稻开花7 d后的种子,取颖果提取总RNA.利用TaKaRa公司的RNA提取试剂盒预处理RNA去除基因组DNA并反转录获得cDNA.以Actin基因为内参,分别以“秀水134”和突变体水稻cDNA为模板,分析Badh2基因的mRNA表达水平.采用2-ΔCt法分析数据,确定基因的相对表达量.
1.2.5 突变体植株成熟种子2-乙酰-1-吡咯啉(2AP)含量测定
采用GC-MS法测定“秀水134”水稻和突变体水稻成熟种子香味物质2AP(2-acetyl-1-pyrroline)含量.2AP提取和测定方法参照HUANG等[13]和应兴华等[14]的报道.
1.2.6 突变体植株主要农艺性状及产量性状分析
水稻成熟时,在野生型“秀水134”和纯合突变体植株种植的小区中,随机选取5棵单株,分别测量剑叶高和株高,调查每株的有效穗数和每穗实粒数,称量千粒重和单株重,最后利用Excel进行统计分析,并用t检测对数据进行统计分析.
2 结果与分析
2.1 转基因植株PCR检测及突变类型分析
使用F0和R0引物对转基因T0代植株进行PCR檢测.结果显示所有转基因植株都能够扩增出789 bp目标条带.
经PCR鉴定后的阳性植株分别使用可扩增相应Target位点的引物,对目的基因Badh2进行扩增和测序分析.通过DANMAN比对及测序峰图分析测序结果发现,在由含有Target-1序列的表达载体转化获得的转基因植株中,有两棵T0代转基因阳性植株表现出两种纯合突变,分别被定名为badh2-1和badh2-2.badh2-1突变是在目的基因第二外显子328 bp处有一个碱基T插入,badh2-2是在目的基因第二外显子329 bp和335 bp处缺失ACCTCCG 7个碱基,如图1(b)所示.
2.2 突变体目的基因编码的氨基酸序列分析
纯合突变体badh2-1目的基因编码的氨基酸相比野生型植株Badh2编码的氨基酸,在第77个氨基酸处开始发生移码突变,并提前终止.突变体badh2-1 Badh2基因编码的蛋白质由原来503个氨基酸变成468个氨基酸,但两者在第77个氨基酸后,氨基酸种类都发生了变化(图2).
纯合突变体badh2-2目的基因在第二外显子329 bp和335 bp处发生了7个碱基缺失,也同样使Badh2基因编码氨基酸时发生移码突变和提前终止,由原来503个氨基酸缩短为481个氨基酸,同时在第76个氨基酸后的氨基酸种类也全部发生了改变(图2).
2.3 突变体T1植株基因组载体检测
从两种纯合突变体T0植株上收获T1种子.将T1种子种植于上海师范大学校植物园内.苗期取叶片提取DNA,使用F0和R0引物,对野生型“秀水134”和两种纯合突变体T1植株进行载体扩增,结果发现在有些植株基因组中依然能够扩增出载体片段,有6棵T1植株无法扩增出载体(图3).由此表明,载体在T1植株中发生了分离.保留无载体骨架的T1植株.
2.4 突变体植株Badh2基因表达分析
对野生型“秀水134”水稻和两种突变体badh2-1和badh2-2植株Badh2基因进行定量PCR检测,结果显示两种不同突变类型植株的Badh2基因转录水平差异不显著(p>0.05),但两种不同突变类型植株的Badh2基因转录水平都比野生型“秀水134”水稻呈极显著下调(p<0.01)(图4).
2.5 突变体植株成熟种子2AP含量分析
检测野生型“秀水134”和两种突变体植株成熟种子的2AP含量,结果显示两种突变体植株成熟种子香味物质2AP的含量都极显著高于野生型“秀水134”水稻(p<0.01),但两种不同突变体植株成熟种子2AP的含量差异不显著(p>0.05)(图5).
2.6 突变体植株主要农艺性状及产量性状分析
为了明确香味基因Badh2突变是否影响水稻植株主要农艺性状及产量性状,对野生型“秀水134”及其突变体badh2-1植株主要农艺性状和产量性状进行了考查,发现野生型“秀水134”水稻与突变体植株的剑叶高、株高、有效穗数都没有显著差异(p>0.05)(表2).产量性状分析显示,野生型“秀水134”水稻与突变体植株的每穗实粒数、千粒重及单株重之间也没有显著差异(p>0.05)(表3).
3 討 论
过去对于香稻育种主要使用常规选育方法.大多先将香型水稻与具有多种有利性状的非香型水稻杂交并自交,然后通过咀嚼或氢氧化钾溶液浸泡对F2代植株米粒是否具有香味进行鉴定,保留米粒具有香味的单株,继续通过多代自交方式培育出香稻新品种.常规选育方法很难培育出高产、优质等综合性状好的香型水稻新品种.利用分子标记辅助育种技术选育香型水稻,虽然可以将杂交后的植株多次与具有多种有利性状的非香型水稻进行回交,使选育出的香稻品种聚集更多有利性状,但也无法避免育种周期长的问题.
快速培育香型水稻一直以来都是水稻育种家们追求的目标.分子生物学新技术的发展已实现了缩短香稻的育种进程.2015年,SHAN等[15]通过TALEN技术,对“日本晴”水稻香味基因Badh2进行编辑,获得了大约30%的Badh2基因杂合后代.2017年,邵高能等[16]利用CRISPR/Cas9技术对“中花11”水稻Badh2基因进行编辑,确认了利用CRISPR/Cas9技术能够快速培育香型水稻的可行性.2017年和2018年,周文甲等[17]和王付华等[18]也采用CRISPR/Cas9技术分别对黑龙江省和河南省水稻品种的Badh2基因进行编辑,也获得了相应的香型水稻.与TALEN技术相比,CRISPR/Cas9技术是一项能够对目的基因特定位点进行更有效编辑的技术,实验成本低,成功率高,并且可使育种时间更短.
本研究也采用CRISPR-Cas9技术,对上海市目前主栽的重要常规水稻“秀水134” Badh2基因进行编辑,也获得了两种Badh2基因纯合突变的“秀水134”水稻.检测分析显示,两种突变体植株成熟种子香味物质2AP含量都极显著高于野生型“秀水134”水稻.
BRADBURY等[4]最早报道了8号染色体上编码BADH2的基因与水稻香味直接相关.随后,不同的研究者又先后报道了12个位于Badh2不同编码链位点突变的等位基因[5,19-20].SHI等[21]在2014年报道了一个突变位于启动子和5UTR的新Badh2等位基因.香味新等位基因的发现为香稻育种提供了重要的基因资源.本研究结果也为今后香型水稻育种提供了两个有利用价值的新型香味等位基因.
2-乙酰-1-吡咯啉是水稻最重要的香味化合物[22].当Badh2编码链突变将导致BADH2原有功能丧失,使其作用底物2AP的代谢途径中断,2AP不断积累从而使水稻变香[4].MICHAEL等[20]进一步研究发现:Badh2不同编码链位点突变都能够增加水稻香味物质的2AP含量.本研究也进一步证明了这一的结果.
目前在生产上应用的香稻品种大多是通过常规育种获得,产量大多低于非香水稻.本研究通过对“秀水134”和“Badh2突变体”后代农艺性状及产量性状比较分析发现,香味基因Badh2突变并不会影响水稻农艺性状和降低水稻的产量.
本研究通过对T1代植株表达载体分析,已成功获得不具有载体骨架结构的香型“秀水134”水稻.香型“秀水134”水稻的成功培育,不仅可为加快高产香型水稻在上海市的推广应用,同时也为今后利用已成功构建的Badh2敲除载体快速培育其他高产优质香型水稻新品种的研究提供参考.
参考文献:
[1] GARLAND S,LEWIN S,BLAKENEY A,et al.PCR-based molecular markers for the fragrance gene in rice (Oryz sativa L.) [J].Theoretical and Applied Genetics,2000,101(3):364-371.
[2] JIN Q,WATERS D,CORDEIRO G M,et al.A single nucleotide polymorphism (SNP) marker lined to the fragrance gene in rice (Oryza sativa L.) [J].Plant Science,2003,165(2):359-364.
[3] CORDEIRO G M,CHRISTOPHER M J,HENRY R J,et al.Identication of microsatellite markers for fragrance in rice by analysis of the rice genome sequence [J].Molecular Breeding,2002,9(4):245-250.
[4] BRADBURY L M,FITZGERALD T L,HENRY R J,et al.The gene for fragrance in rice [J].Plant Biotechnology Journal,2005,3(3):363-370.
[5] SHI W,YANG Y,CHEN S,et al.Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties [J].Molecular Breeding,2008,22(2):185-192.
[6] CHEN S,YANG Y,SHI W,et al.Badh2,encoding betaine aldehyde dehydrogenase,inhibits the biosynthesis of 2-acetyl-1-pyrroline,a major component in rice fragrance [J].Plant Cell,2008,20(4):1850-1861.
[7] NIU X L,TANG W,HUANG W Z,et al.RNAi-directed downregulation of OsBADH2 results in aroma (2-acetyl-1-pyrroline) production in rice (Oryza sativa L.) [J].BMC Plant Biology,2008,8(1):100.
[8] 朱映东,时亚琼,周锋利,等.分子标记辅助选育香型巨胚水稻 [J].上海师范大学学报(自然科学版),2013,42(6):623-628.
ZHU Y D,SHI Y Q,ZHOU F L,et al.Development of aromatic giant-embryo rice molecular marker-assisted selection [J].Journal of Shanghai Normal University (Natural Sciences),2013,42(6):623-628.
[9] 蔡治君,黄菊,李建粤,等.分子标记辅助育种选育黑色香型巨胚水稻 [J].分子植物育种,2014,12(6):1112-1118.
CAI Z J,HUANG J,LI J Y,et al.Developing black aromatic giant-embryo rice by molecular marker assisted breeding [J].Molecular Plant Breeding,2014,12(6):1112-1118.
[10] ZHAO G C,XIE M X,LI J Y.Development of a new fragrant and good eating quality rice variety with stripe virus disease resistance by molecular marker-assisted gene pyramiding [J].Indian Journal of Genetics and Plant Breeding,2017,77(2):221-227.
[11] 周金玉,姚全甫.秀水134的特性及高产栽培技术 [J].中国稻米,2012,3:78-79.
[12] 李建粤,吕英海,杨丽君,等.转反义蜡质基因“湘晴”及其杂交稻米的直链淀粉含量研究 [J].西北植物学报,2008,28(6):1082-1087.
LI J Y,LYU Y H,YANG L J,et al.Reducing amylose content of “Xiang Qing” and hybrid rice seeds by introducing anti-waxy gene [J].Acta Botanica Boreali-Occidentalia Sinica,2008,28(6):1082-1087.
[13] HUANG T C,TENG C S,CHANG J L,et al.Biosynthetic mechanism of 2-Acetyl-1-pyrroline and its relationship with Δ1-pyrroline-5-carboxylic acid and methylglyoxal in aromatic rice (Oryza sativa L.) callus [J].Journal of Agricultural and Food Chemistry,2008,56(16):7399-7404.
[14] 应兴华,徐霞,陈铭学,等.气相色谱-质谱技术分析香稻特征化合物2-乙酰基吡咯啉 [J].色谱,2010,28(8):782-785.
YING X H,XU X,CHEN M X,et al.Determination of 2-acetyl-1-pyrroline in aroma rice using gas chromatography-mass spectrometry [J].Chinese Journal of Chromatography,2010,28(8):782-785.
[15] SHAN Q W,ZHANG Y,CHEN K L,et al.Creation of fragrant rice by targeted knockout of the OsBADH2 gene using TALEN technology [J].Plant Biotechnology Journal,2015,13(4):791-800.
[16] 邵高能,谢黎虹,焦桂爱,等.利用CRISPR/CAS9 技術编辑水稻香味基因Badh2 [J].中国水稻科学,2017,31(2):216-222.
SHAO G N,XIE L H,JIAO G A,et al.CRISPR/CAS9-mediated editing of the fragrant gene Badh2 in rice [J].China Journal Rice Science,2017,31(2):216-222.
[17] 周文甲,田晓杰,任月坤,等.利用CRISPR/CAS9创造早熟香味水稻 [J].土壤与作物,2017,6(2):146-152.
ZHOU W J,TIAN X J,REN Y K,et al.Breeding of early-maturatity and fragrant rice via CRISPR/Cas9 mediated genome editing [J].Soils and Crops,2017,6(2):146-152.
[18] 王付华,薛华政,王亚,等.利用CRISPR/CAS9基因编辑技术创制香型郑稻19新种质 [J].作物杂志,2018(6):36-42.
WANG F H,XUE H Z,WANG Y,et al.Breeding fragrant rice zhengdao19 using CRISPR/Cas9 mediated gene editing technology [J].Crops,2018(6):36-42.
[19] SHAO G N,TANG A,TANG S Q,et al.A new deletion mutation of fragrant gene and the development of three molecular markers for fragrance in rice [J].Plant Breeding,2011,130(2):172-176.
[20] MICHAEL J,KOVACH A,MARIAFE N,et al.The origin and evolution of fragrance in rice (Oryza sativa L.) [J].Proceedings of the National Academy of Sciences,2009,106 (34):14444-14449.
[21] SHI Y Q,ZHAO G C,LI J Y.Discovery of a new fragrance allele and development of functional markers for identifying diverse fragrant genotypes in rice [J].Molecular Breeding,2014,33(3):701-708.
[22] WIDJAJA R,CRASKE J D,WOOTTON M.Comparative studies on volatile components of non-fragrant and fragrant rice [J].Journal of the Science of Food and Agriculture,1996,70(2):151-161.
(責任编辑:顾浩然)
猜你喜欢
东坡赤壁诗词(2023年6期)2023-12-26
休闲读品·天下(2022年2期)2022-07-13
休闲读品·天下(2022年1期)2022-05-01
金山(2016年12期)2017-02-17
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
艺术评鉴(2016年14期)2016-05-30
天津医科大学学报(2015年2期)2015-12-22
读写算(中)(2015年25期)2015-11-07
山东医药(2015年40期)2015-02-28
