
粒用高粱×苏丹草杂交F2代农艺性状的数量遗传分析
2019-12-16 01:42白晓倩卢华雨于澎湃裴忠有罗峰孙守钧
江苏农业科学 2019年19期
白晓倩 卢华雨 于澎湃 裴忠有 罗峰 孙守钧

摘要:选取各农艺性状均较大的国内粒用高粱品种忻粱52和从美国普渡大学引进的苏丹草品系美引-48进行杂交,得到F2代分离群体,对F2代的开花期、株高、穗长、穗柄长、旗叶鞘长、叶片数、平均茎节长等7个农艺性状进行测定。利用主基因-多基因遗传分析方法进行数据分析,得到7个性状的4个遗传备选模型,并进行适合性检验,从备选模型中选出控制性状遗传的最适遗传模型,并根据IECM(迭代ECM)算法计算主基因遗传率。结果表明,株高、穗柄长、旗叶鞘长、平均茎节长等性状均符合ModelA_0模型,均为微效多基因控制的数量性状;开花期、穗长均符合ModelB_6模型,这2个性状均符合2对主基因控制的等显性遗传模型,即da=db=ha=hb,2对主基因的遗传率为 38.35%;叶片数符合ModelB_1模型,是2对主基因控制的加性-显性-上位性混合遗传模型,2对主基因的加性效应之和为2.096 774,显性效应之和为0.403 226,主基因遗传率为99.22%,遗传率极高,可在育种后代中直接进行选择。
关键词:高粱;农艺性状;主基因-多基因分析;遗传率
中图分类号: S544+.103文献标志码: A
文章编号:1002-1302(2019)19-0188-06
收稿日期:2018-06-26
基金项目:天津市科技支撑计划(编号:16YFZCNC00630)。
作者简介:白晓倩(1993—),女,天津人,硕士研究生,主要从事饲用作物遗传改良研究。E-mail:1044235123@qq.com。
通信作者:孙守钧,博士,教授,主要从事饲用作物遗传改良研究,E-mail:sunshoujun@tjau.edu.cn;罗 峰,硕士,副教授,主要从事饲用作物遗传改良研究,E-mail:luofeng1868@126.com。
高粱和苏丹草是隶属于高粱属的2个种,两者均为喜温型一年生禾本科作物,且在形态学上差异较大,但是其生殖隔离不是很明显,染色体均为2n=20[1-6]。高粱是世界第五大粮食作物,也是重要的饲用作物和酿造原料[7],具有抗旱耐涝、耐贫瘠和耐盐碱等优良特性,但是其品质欠佳,茎秆中的粗蛋白含量较低且氢氰酸含量较高,不宜作为饲用原料来进行加工。苏丹草原产于非洲苏丹高原[8],是一种既具有高粱的抗旱、抗病虫害、抗倒伏等特点,又具有分蘖力强、再生性强、适应范围广、适口性好等优点的优质牧草,尤其是其营养价值较高,其茎秆中的粗蛋白含量较高,氢氰酸含量较低,是饲用的重要原料[9-12]。
饲草作物指用作家畜饲料的饲用作物,饲草作物是发展草食家畜的重要基础与原料,而粒用高粱与苏丹草杂交属于种间杂交,其杂交种简称为高丹草,综合了高粱和苏丹草的优良性状,具有很强的杂种优势,在饲用方面具有较大的发展前景[13]。
目前,国内外对于粒用高粱和甜高粱的研究较多,在饲草作物性状遗传方面的研究较少[14]。本研究通过粒用高粱与苏丹草杂交F2代的数量遗传分析,得出高丹草主要农艺性状的数量遗传规律,以期为饲草高粱育种提供理论依据。
1 材料与方法
1.1 试验材料
本试验选取性状差异较大的国内粒用高粱品种忻粱52和从美国引进的苏丹草品系美引-48為试验材料进行杂交,得到F1代杂交种,经F1代自交得到F2代分离群体。
1.2 试验设计
本试验于2016年11月在海南省乐东黎族自治县利国镇官村进行播种,同年12月对2个亲本进行杂交,得到F1代杂交种。2017年5月于天津市静海良种场种植F1代正反交组合,经检验,F1代正反交农艺性状长势一致,不存在细胞质遗传;2017年7月对F1代植株进行套袋,使其自交授粉,得到F2代分离群体,2017年11月在海南省乐东黎族自治县按照亲本、F1代正反交组合、F2代分离群体的顺序进行播种,所有种植材料的行长均为5.5 m,行间距为0.5 m,株距为0.3 m,田间管理同常规农田。2018年1月,在高粱完熟期测量各农艺性状。
1.3 测定方法
由于F2群体单株之间的表型性状和基因型均不同,因此以下性状测量的均为单株数据。
(1)开花期。在开花期,从F2群体开花第1天起,将第1天开花的单株记录为1,第2天开花的单株记录为2,以此类推(单位:d)。
(2)株高。在完熟期,用钢卷尺测量地面至穗顶端的距离(单位:cm)。
(3)穗长。在完熟期,用钢卷尺测量穗基部到穗顶端的距离(单位:cm)。
(4)穗柄长。在完熟期,用钢卷尺测量穗基部到第1节间的距离(单位:cm)。
(5)旗叶鞘长。用钢卷尺测量高粱完熟期旗叶叶鞘到第1节间的距离(单位:cm)。
(6)叶片数。在高粱完熟期,计算整株的叶片数量(单位:张)。
(7)平均茎节长。平均茎节长=(完熟期株高-完熟期穗长)/完熟期茎节数(单位:cm)。
1.4 数据处理
基本数据使用Excel 2010和SPSS Statistics 17.0进行处理,根据王建康等提出的单个分离世代群体的遗传模型方法(主基因-多基因混合遗传的分离分析方法[15-17])进一步对F2世代群体进行分析,选出最适遗传模型,根据IECM(迭代ECM)算法计算控制性状的主基因遗传率。
2 结果与分析
2.1 农艺性状间的相关性分析
由表1可知,开花期与叶片数呈极显著正相关,相关系数为0.225,与株高呈显著正相关,相关系数为0.130,与穗柄长、旗叶鞘长呈显著负相关,相关系数分别为-0.142、-0.122;株高与穗长、穗柄长、旗叶鞘长、叶片数和平均茎节长均呈极显著正相关,其中与旗叶鞘长的相关性最高,相关系数为 0.566;穗长与穗柄长、旗叶鞘长、叶片数、平均茎节长呈极显著正相关,相关系数最高的为旗叶鞘长,为0.622;穗柄长与旗叶鞘长、平均茎节长呈极显著正相关,相关系数分别为 0.266、0.510,与叶片数呈极显著负相关,相关系数为 -0.228;旗叶鞘长与叶片数、平均茎节长呈极显著正相关,相关系数分别为0.282、0.247;叶片数与平均茎节长呈现极显著负相关,相关系数为-0.433。
2.2 各农艺性状的表型分析
由表2和图1可以看出,开花期、株高、穗长、穗柄长、旗叶鞘长、叶片数及平均茎节长均呈现连续的正态分布,说明这6个性状符合数量性状的遗传特征,属于数量遗传性状。
2.3 农艺性状遗传模型的选择
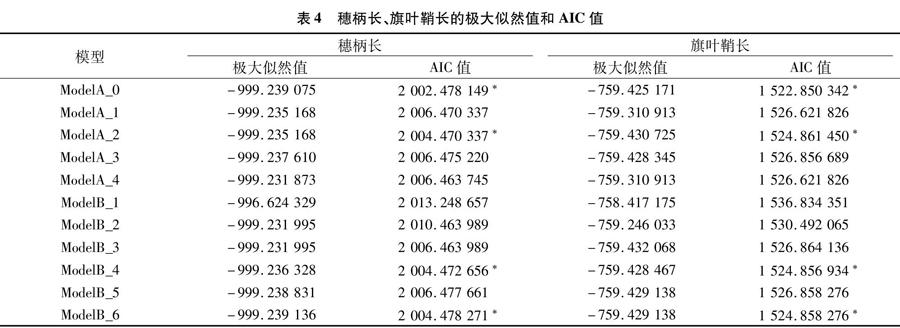
根据盖钧镒等的主基因-多基因混合遗传模型F2代单世代分离分析方法[18-19],得到开花期、株高、穗长、穗柄长、旗叶鞘长、叶片数、平均茎节长这7个农艺性状的极大似然值和AIC(Akaike information criterion)值(在表3、表4、表5),根据AIC值最小原则,选出4个遗传备选模型(表3、表4、表5中用“*”标注)进行下一步适合性检验,以期确定各性状的数量遗传模型。
2.4 农艺性状遗传模型的检测
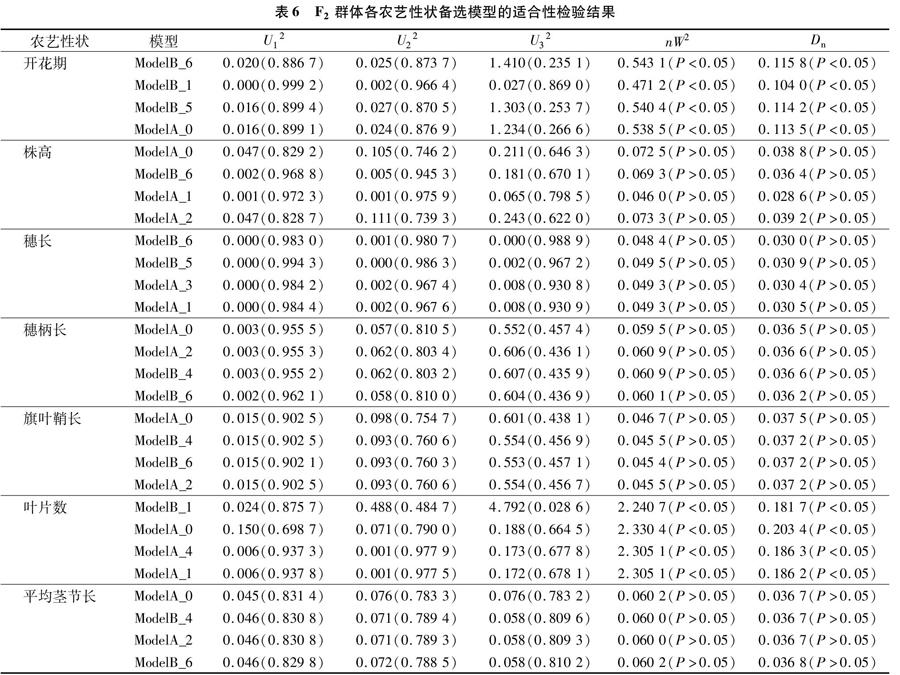
根据U12、U22、U32(均匀性检验)、nW2(Smirnov检验)、Dn(Kolmogorov检验),对开花期、株高、穗长、穗柄长、旗叶鞘长、叶片数、平均茎节长这7个农艺性状的4个备选模型进行适合性检验,结果见表6,选出7个性状最适的遗传模型。
(1)在开花期的4种遗传备选模型中,其nW2(Smirnov检验)和Dn(Kolmogorov检验)检验均达到显著水平(P<0.05),而U12、U22、U32(均匀性检验)未达到显著水平,且ModelB_6的AIC值最小;因此,选择Model B_6为开花期的最适遗传模型,说明在该群体中,开花期为2对主基 因+ 多基因混合遗传,为等显性混合遗传模型。(2)在株高的4种备选模型中,其均匀性检验、Smirnov检验和Kolmogorov检验均未达到显著水平,但是相较于另外3种模型来说,Model A_0的AIC值最小,因此选择Model A_0作为株高的最适遗传模型,说明该群体的株高不受主基因控制,而是受微效多基因控制。(3)在穗长的4个遗传备选模型中,其均匀性检验、Smirnov检验和Kolmogorov检验均未达到显著水平,但Model B_6的AIC值小于另外3个备选模型,因此选择Model B_6为该群体穗长的最适遗传模型,说明该性状受2对主基因+多基因混合遗传,为等显性混合遗传模型。(4)穗柄长的4个遗传备选模型的均匀性检验、Smirnov检验和Kolmogorov检验均未达到显著水平,但相较于其他3个备选模型,Model A_0 的AIC值最小,因此选择Model A_0作为该群体穗柄长的最适遗传模型,说明该性状没有主基因控制,而是受微效多基因控制。(5)在旗叶鞘长的4个遗传备选模型中,其U12、U22、U32(均匀性检验)与nW2(Smirnov检验)、Dn(Kolmogorov检验)均未达到显著水平,而Model A_0的AIC值是4个备选模型中最小的,所以选择Model A_0作为旗叶鞘长的最适遗传模型,解释了该群体旗叶鞘长是受微效多基因控制而非主基因控制。(6)在叶片数的4种遗传备选模型中,nW2(Smirnov检验)和Dn(Kolmogorov检验)均达到显著水平(P<0.05),其中Model B_1 的均匀性检验中的U32也达到显著水平(P<0.05),而其他统计量未达到显著水平,且在4个备选模型中Model B_1的AIC值最小,因此选择Model B_1作为该群体叶片数的最适遗传模型,说明在该群体中叶片数是2对主基因+多基因混合遗传,为加性-显性-上位性混合遗传模型。(7)在平均茎节长的4种备选模型中,所有统计量均未达到显著水平,结合AIC值最小原则,选择Model A_0 作为平均茎节长的最适遗传模型,该性状不存在主基因控制,受微效多基因控制。
2.5 农艺性状遗传参数估计
根据已经确定的各个性状的最优模型和IECM的估算方法[20-21],对F2群体农艺性状进行遗传参数估计,结果见表7。由于株高、穗柄长、旗叶鞘长和平均茎节长都符合Model A_0,不存在控制性状遗传的主基因,属于微效多基因控制的数量性状遗传,因此无法计算主基因遗传率。
F2分離群体的开花期遗传符合B_6模型,F2群体的开花期符合等显性合遗传模型,说明2对主基因的显性效应相等并与2对主基因的加性效应也相等,即da=db=ha=hb(显性度ha/da=hb/db=1),且上位性效应i=jab=jba=0;其中,开花期的平均值m为8.54,偏向于开花期较早的亲本忻 粱-52,控制开花期的2对主基因的加性效应和显性效应相等,均为1.299 9,主基因遗传率为38.35%,遗传率较低,说明开花期遗传不稳定,易受环境影响。
穗长符合Model B_6模型,说明该群体的穗长是受2对主基因控制的等显性遗传模型,即2对主基因的显性效应相等,da=db,这种情况下的9种基因型的比例[(AABB+AABb+AaBB+AaBb) ∶(AAbb+Aabb+aaBB+aaBb) ∶aabb]为9 ∶6 ∶1 ∶2,对主基因的遗传率为44.23%,遗传率较低,说明该群体的穗长遗传不稳定,受环境因素的影响较大。
叶片数符合Model B_1模型,即加性-显性-上位性混合遗传模型,受2对主基因控制;第1对主基因的加性效应(da)、显性效应(ha)、显性度(ha/da)分别为1.548 4、0.451 6、0.291 7,第2对主基因的加性效应(db)、显性效应(hb)、显性度(hb/db)分别为0.548 4、0.048 4、0.107 2;其中 |da|>|db|、|ha|>|hb|,说明第1对主基因的加性作用和显性作用均大于第2对主基因,且显性度均小于1,且2对主基因的加性效应之和为2.096 774,显性效应之和为0.403 226,说明控制该性状的2对主基因均以加性效应为主;主基因遗传率为99.22%,即主基因遗传率高,易在后代中遗传,受环境的影响较小。
3 结论与讨论
高粱农艺性状的研究,对高粱育种有至关重要的作用。本试验通过对粒用高粱忻粱-52和苏丹草美引-48杂交F2代的数量遗传分析,得到了主要农艺性状的遗传模型及遗传率,为今后的高粱育种提供了参考。
本研究结果表明,在忻粱-52和美引-48的F2代分离群体中,株高、穗柄长、旗叶鞘长和平均茎节长都符合Model A_0模型,说明这4个农艺性状均无主基因控制,属于微效多基因控制的数量性状。其中,旗叶鞘长与孙守钧等的研究结果[22]一致。在株高的数量遗传分析中,本研究结果与杜希朋等关于小麦的研究结果[23]一致。官延安等的研究结果[24-25]均为有2对主基因+多基因控制的B_1模型,即加性-显 性- 上位性混合遗传模型。詹秋文等的研究结果为有2对主基因+多基因控制的B_6模型,即等显性模型[13]。蒋峰等的研究结果为株高符合由2对主基因控制的B_2模型,为加性-显性混合遗传模型[26]。造成结果不一致的原因,一方面可能是选择杂交的亲本不同,虽然高粱、苏丹草均为高粱属作物且生殖隔离不明显,但二者均为高粱属下的2个不同种;另一方面是种植环境不同,本试验中的亲本、F1代、F2代均在海南省种植,南北方环境差异较大,包括光、温、气、热等,且海南省田间多为沙质土,保水保肥差,北方多为壤质土,保水肥能力强。
开花期符合Model B_6模型,为2对主基因+多基因控制的等显性混合遗传模型,即2对主基因的加性效应与显性效应相等,遗传率为38.35%,遗传率较低,说明该性状受环境影响较大。本研究与徐小万等对辣椒开花期的遗传分析结果[27]不同,其分析结果为开花期符合2对主基因控制的加 性- 显性-上位性混合遗传模型。穗长符合2对主基因+多基因控制的等显性模型,遗传率为44.23%,同样受环境影响较大,该结果与李延玲等对高粱的研究结果[25]一致,与杜希朋等关于小麦的研究结果[23,28-29]不一致。前者研究表明,小麦穗长为多基因控制,无主基因控制,后者采用质量-数量性状遗传的极大似然分析认为,小麦穗长受1对主基因以及若干微效多基因控制。叶片数符合Model B_1模型,属于2对主基因控制的加性-显性-上位性混合遗传模型,2对主基因的加性效应之和为2.096 8,显性效应之和为0.5,加性效应显著大于显性效应,说明控制该性状以加性效应为主,2对主基因的遗传率为99.22%,遗产率较高,在后代遗传中不易受到环境的影响。
综上所述,在忻粱-52×美引-48的F2代分离群体的7个主要农艺性状的遗传分析中,研究结果与前人的差异较大;只有开花期、穗长、叶片数存在主基因控制,而开花期和穗长的遗传率较低,在后代中受环境影响较大,不易稳定遗传,叶片数的遗传率较高,可以在早期世代进行选择。在高粱遗传育种中,数量性状不仅受到遗传因子的控制,外界环境因素也对性状的遗传有较大影响,因此,在杂交后代的遗传研究中,尤其应注意环境对各性状的影响。
参考文献:
[1]詹秋文. 高粱-苏丹草杂交种生物能源利用的前景分析[J]. 种子,2009,28(8):81-83.
[2]詹秋文,钱章强. 高粱与苏丹草杂种优势利用的研究[J]. 作物学报,2004,30(1):73-77.
[3]詹秋文,林 平,李 军,等. 高粱-苏丹草杂交种研究与利用前景[J]. 草业学报,2001,10(2):56-61.
[4]王和平,张福耀,程庆军,等. 高粱-苏丹草杂交草的研究与利用[J]. 杂粮作物,2000,20(4):20-23.
[5]MurtyU R. Apomictic and sexual reproduction in sorghum[J]. Indian Genet Plant Breed,1989,39(2):271-278.
[6]赵晓杰,于 卓,刘永伟,等. 高粱314A、13A与苏丹草杂种F1代的农艺特性及细胞学分析[J]. 西北植物学报,2005,25(6):1107-1113.
[7]高士杰,劉晓辉,李继洪,等. 我国粒用高粱育种现状、问题与对策[J]. 作物杂志,2006,3(4):11-13.
[8]支中生,张恩厚,高卫华,等. 苏丹草与高粱杂交后代特征及其主要经济性状[J]. 草地学报,2002,10(2):144-150.
[9]杨小翠,詹秋文,刘言龙,等. 苏丹草与高粱的判别分析和聚类分析[J]. 中国草地学报,2016,38(1):41-46.
[10]柯 梅,朱 昊,梁维维,等. 苏丹草农艺性状与产量、品质间的灰色关联度分析[J]. 草业科学,2016,33(5):949-955.
[11]李陈建,付彦博,万江春,等. 30份苏丹草种质资源农艺性状的遗传多样性分析[J]. 草业科学,2015,32(1):85-93.
[12]逯晓萍,云锦凤,肖宇红,等. 高丹草(高粱×苏丹草)产量及其构成因素的QTL定位与分析[J]. 华北农学报,2007,22(4):80-85.
[13]詹秋文,林 平,李 娣,等. 高丹草重要农艺性状的遗传分析[J]. 南京农业大学学报,2010,33(3):26-30.
[14]逯晓萍,米福贵,郭世华,等. 高丹草(高粱×苏丹草)主要农艺性状的遗传参数研究[J]. 华北农学报,2004,19(3):22-25.
[15]王建康,盖钧镒. 数量性状主-多基因混合遗传的P1、P2、F1、F2和F2 ∶3联合分析方法[J]. 作物学报,1998,24(6):651-659.
[16]盖钧镒,王建康. 利用回交或F2 ∶3家系世代鉴定数量性状主基因-多基因混合遗传模型[J]. 作物学报,1998,24(4):402-409.
[17]王建康,盖钧镒. 利用杂种F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应[J]. 遗传学报,1997,24(5):432-440.
[18]盖钧镒,章元明,王建康. QTL混合遗传模型扩展至2对主基 因+ 多基因时的多世代联合分析[J]. 作物学报,2000,26(4):385-391.
[19]章元明,盖钧镒. 利用DH或RIL群体检测QTL体系并估计其遗传效应[J]. 遗传学报,2000,27(7):634-640.
[20]章元明,盖钧镒. 数量性状分离分析中分布参数估计的IECM算法[J]. 作物学报,2000,26(6):699-706.
[21]盖钧镒. 植物数量性状遺传体系的分离分析方法研究[J]. 遗传,2005,27(1):130-136.
[22]孙守钧,张云华,马鸿图. 高粱旗叶鞘长及穗柄长的遗传研究[J]. 国外农学-杂粮作物,1998,18(6):9-12.
[23]杜希朋,闫媛媛,刘伟华,等. 蚂蚱麦×碧玉麦杂交F2代部分重要农艺性状的遗传分析[J]. 麦类作物学报,2011,31(4):624-629.
[24]管延安,张华文,樊庆琪,等. 普通高粱与甜高粱杂交组合株高、糖度的主基因多基因模型遗传效应分析[J]. 核农学报,2012,26(1):36-42.
[25]李延玲,白晓倩,于澎湃,等. 高粱株行性状数量遗传分析[J]. 华北农学报,2018,33(1):143-149.
[26]蒋 峰,刘鹏飞,曾慕衡,等. 甜玉米株高的多世代遗传分析与QTL定位[J]. 西北农林科技大学学报(自然科学版),2011,39(3):67-74.
[27]徐小万,曾 莉,李 颖,等. 辣椒开花期的主基因+多基因遗传分析[J]. 生物数学学报,2012,27(4):753-757.
[28]卢 翔,张锦鹏,王化俊,等. 小麦-冰草衍生后代3558-2穗部相关性状的遗传分析和QTL定位[J]. 植物遗传资源学报,2011,12(1):86-91.
[29]张安静,张俊祖,刘凤琴,等. 超大穗小麦穗长和小穗数的配合力及遗传模型分析[J]. 麦类作物学报,2006,26(4):41-43.
猜你喜欢
古今农业(2022年2期)2022-08-15
今日农业(2021年20期)2021-11-26
疯狂英语·新读写(2021年2期)2021-02-25
山东农业科学(2016年12期)2017-01-21
长江蔬菜·学术版(2016年12期)2017-01-12
