
槟榔Catalase基因的克隆及亚细胞定位
2020-02-22 03:19梁婷沈文涛庹德财言普黎小瑛唐庆华周鹏
热带作物学报 2020年12期
梁婷 沈文涛 庹德财 言普 黎小瑛 唐庆华 周鹏


摘 要:过氧化氢酶(Catalase)广泛存在于生物体内,主要功能是清除植物细胞内光呼吸、线粒体电子传递及脂肪β-氧化等过程中产生的H2O2,从而保护细胞免于过氧化氢酶的毒害。为了解槟榔过氧化氢酶基因的相关信息,利用植物总RNA提取试剂盒提取槟榔叶片总RNA,通过同源克隆和RACE技术获得槟榔全长cDNA,并对其进行生物信息学分析及亚细胞定位。结果表明:获得ArCAT1(MN692600)、ArCAT2(MN692601)、ArCAT3(MN692602)3个同源基因,其序列全长均为1476 bp,编码492个氨基酸。遗传进化分析显示,ArCATS与同为棕榈科的油棕(Elaeis guineensis)、海枣(Phoenix dactylifera)等植物的过氧化氢酶氨基酸序列具有较高的相似性,其中与油棕的亲缘关系最近。亚细胞定位结果表明,ArCAT1蛋白定位于过氧化物酶体中,而ArCAT2和ArCAT3既定位于过氧化物酶体又定位于细胞核中。这为进一步研究Catalase基因在槟榔中的生物学功能奠定基础。
关键词:槟榔;过氧化氢酶;亚细胞定位;基因克隆
中图分类号:Q23 文献标识码:A
Abstract: The main function of catalase, widely existed in organisms, is to remove H2O2 produced in the process of photorespiration, mitochondrial electron transmission and fat betao-oxidation in plant cells to protect cells from the toxicity of H2O2. In order to understand the relevant information of areca catalase gene, RNA was extracted from areca leaves, the full length of areca cDNA was obtained through the homologous cloning and RACE technology, and bioinformatics analysis and subcellular localization were performed. The results showed that the total length of ArCAT1 (MN692600), ArCAT2 (MN692601) and ArCAT3 (MN692602) gene sequence was 1476 bp, which could encode 492 amino acids. Genetic evolutionary analysis showed that ArCATS was closely related to Elaeis guineensis and Phoenix dactylifera. The results of subcellular localization showed that ArCAT1 protein was located in the peroxidosome, ArCAT2 and ArCAT3 were located in the peroxidosome and nucleus. This would lay foundation for further research on the biological function of the gene.
Keywords: Areca catechu L.; catalase; subcellular localization; gene cloning
DOI: 10.3969/j.issn.1000-2561.2020.12.002
过氧化氢酶是抗氧化酶中首先被发现的一类末端氧化酶,Loew[1]将其命名为Catalase,是由4个相同肽链亚基组成的四聚体血红素酶,它也是一种高度保守的金属酶,具有保守的活化中心和铁血红蛋白结合位点,存在于许多好氧和厌氧生物中,包括细菌、真菌、植物和动物细胞,在植物体内主要存在于过氧化物酶体、乙醛酸循环体和细胞质中,少数分布在线粒体中[2],主要功能是清除植物细胞内光呼吸、线粒体电子传递及脂肪β-氧化等过程中产生的H2O2,从而保护细胞免于过氧化氢酶的毒害[3]。通过提高植物体内抗氧化酶活性,增强抗氧化代谢水平,可提高植物自身的抗逆性[4]。Catalase与超氧化物歧化酶(SOD)、过氧化物酶(POD)、抗坏血酸酶(APX)被称为抗氧化酶保護系统[4]。近年来,已有研究结果发现,Catalase在植物生长发育、逆境胁迫防御应答、氧化衰老等生理过程中起着至关重要的作用[2]。Catalases家族由多基因编码,其表达和活性受许多因素的影响,如温度、光照、干旱、高盐、重金属等及生物因子等的影响[2]。目前已经在烟草[5]、拟南芥[6]、玉米[7]、南瓜[8]和水稻[9]等被子植物中发现了3种过氧化氢酶基因。在烟草中发现的3种过氧化氢酶中,CAT1负责清除光呼吸产生的H2O2,CAT2则清除氧化胁迫产生的H2O2,CAT3主要清除乙醛酸循环体中产生的H2O2[10]。
槟榔(Areca catechu L.)是槟榔属棕榈科槟榔亚科槟榔族槟榔亚族的常绿乔木,是一种典型的热带植物,一般生长于高温高湿的热带雨林中,在低海拔、温差变化不大的地区会生长得更好,原产于马来西亚,中国主要分布在云南、海南及台湾等热带地区、亚洲热带地区。槟榔含有生物碱、鞣质、黄酮、萜类等多种化学成分,是重要的中药材,具有祛痰止咳、消积、下气、行水、消肿、截疟、消食醒酒、宽胸止吐、驱虫等作用,是我国重要的四大南药资源之一,经济价值很高,在南方一些少数民族还有将果实作为一种咀嚼嗜好品,可御寒和消除紧张劳动后的疲劳,槟榔树易发生的病害有叶斑病、叶枯病、炭疽病、疫病、果穗枯萎病、叶细菌性条斑病、芽腐病、黄化病等[11-14]。已有研究结果表明,Catalase参与了许多植物的抗病反应,过表达Catalase可提高植物的抗逆性,通过增强抗氧化酶活性及增强活性氧代谢来增强植物的耐受性,但未见槟榔树在此方面的研究报道。本研究利用槟榔叶片提取RNA,通过同源克隆和RACE技术获得槟榔过氧化氢酶全长cDNA,并对其进行生物信息学分析及亚细胞定位,这为进一步研究过氧化氢酶基因在槟榔中的生物学功能和培育优质抗性的槟榔品种奠定基础。
1 材料与方法
1.1 材料
以‘热研1号槟榔(Areca catechu L.)为供试材料,采自中国热带农业科学院椰子研究所的槟榔园。将槟榔叶片剪碎后,在液氮中快速研磨至粉末状,于?80 ℃超低温冰箱中保存备用。
1.2 方法
1.2.1 叶片总RNA的提取及cDNA合成 采用RNA Prep Pure多糖多酚植物总RNA提取试剂盒(离心柱型)提取槟榔叶片总RNA。RNA提取后,用1%琼脂糖凝胶电泳和超微量核算蛋白测定仪检测RNA的质量和纯度。按照Prime ScriptTM cDNA第一链合成试剂盒说明书方法将RNA反转录成cDNA,于?20 ℃保存备用。
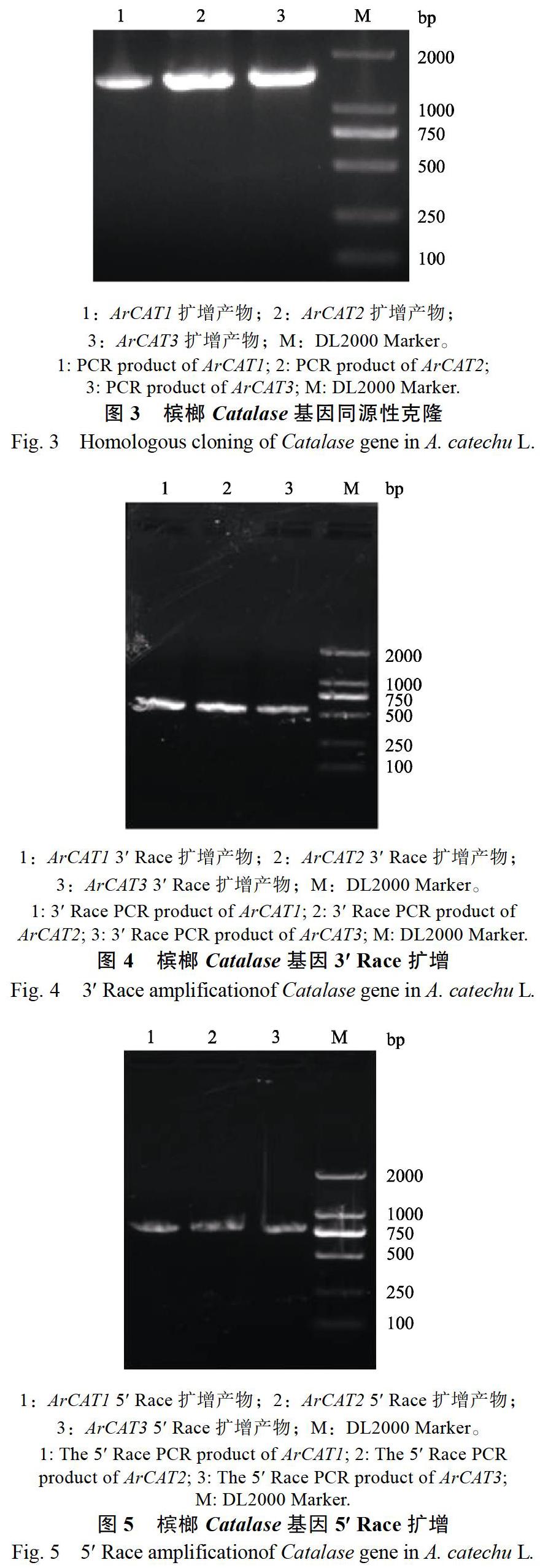
1.2.2 候选基因的同源克隆 利用GenBank数据库中与槟榔同为棕榈科的油棕(Elaeis guineensis)的ElCAT1(XM010919942.3)、ElCAT2(NM 001319913.1)、ElCAT3(XM010943120.3)的氨基酸序列,设计特异性引物CAT1F/CAT1R、CAT2F/CAT2R、CAT3F/CAT3R(表1),以基因组cDNA为模板,使用Prime STAR Max Premix (2) 高保真DNA聚合酶进行基因全长序列的扩增:98 ℃ 30 s;98 ℃ 15 s,53 ℃ 15 s,72 ℃ 5 s,扩增30个循环。扩增后的PCR产物,经1%琼脂糖凝胶电泳检测,用FastPure Gel DNA Extraction Mini Kit试剂盒(诺唯赞公司)回收目的基因片段,回收产物连接TA-Blunt载体(TaKaRa),并转化至DH5α大肠杆菌感受态中,挑选阳性克隆送赛默飞世尔科技有限公司进行测序。将测序正确的菌液提取质粒并于?80 ℃保存备用。
1.2.3 目的基因全长序列克隆 根据1.2.2测序所获得的CAT1、CAT2、CAT3序列,设计3RACE引物和5 RACE引物,按照FirstChoice? RLM- RACE Kit试剂盒说明书进行3 RACE PCR和5 RACE PCR,扩增后的PCR产物,经1%琼脂糖凝胶电泳检测,切胶回收后,回收产物与TA-Blunt载体连接后转化大肠杆菌DH5α,涂板之后筛选阳性单克隆送广州赛默飞世尔科技有限公司进行测序。
1.2.4 生物信息学分析 通过在线软件(http:// www.bio-soft.net/sms/index.html)对槟榔过氧化氢酶蛋白序列进行比对。
在NCBI中下载与槟榔(Areca catechu L.)同为棕榈科的油棕(Elaeis guineensis)和海枣(Phoenix dactylifera)的3种Catalase基因,利用MEGA 11.0软件,采用Neighbor-Joiniing (N-J邻近法)构建系统进化树,Bootstrap值设置为1000。
利用在线软件http://www.csbio.sjtu.edu.cn/中的INSP预测真核蛋白中的核定位信号。
1.2.5 农杆菌转化和亚细胞定位分析
(1)亚细胞定位载体的构建。根据1.2.3获得的槟榔ArCAT1、ArCAT2、ArCAT3编码基因的全长序列,利用Nimble Cloning[15]法分别连接载体pNC-Green-SubC(图1)和pNC-Green-SubN(图2),使GFP荧光蛋白分别位于基因的C端和N端,后转化至DH5α大肠杆菌感受态菌株上,挑选阳性克隆,经过菌液PCR鉴定后,挑选扩增条带最亮且与目的基因大小一致的样品送广州赛默飞世尔科技有限公司测序。
(2)农杆菌的转化。取1 μg质粒DNA加到100 μL农杆菌GV3101感受态细胞中,利用电击法转化农杆菌。加入700 μL LB液体培养基,28 ℃、180 r/min培养2 h后均匀涂布在含80 μg/mL卡那霉素和40 μg/mL利福平的平板上。28 ℃培养48 h形成单菌落。
(3)农杆菌注射本氏烟。挑取农杆菌单菌落接种于200 μL LB液体培养基中,28 ℃、180 r/min培养至OD600约为1.0。5000 r/min离心5 min弃上清,加入5 mL接种缓冲液(含有终浓度为10 mmol/L MgCl2,50 mmol/L MES,pH5.7和100 μmol/L acetosyringone)轻轻悬浮,5000 r/min离心5 min弃上清,重新加入接種缓冲液至OD600在0.3~0.4左右即可,室温静置暗处理3 h,进行接种。用一次性注射器分别吸取1 mL菌液,取针头在受体叶片上轻刺2~3个小孔,将注射器压在针孔部位,以手指抵住叶片下部,轻轻用力将注射器内菌液压送并渗透到叶片组织中,用记号笔在注射部位上做好标记。接种后的烟草25 ℃培养48 h,剪取注射部位15 mm2左右在激光共聚焦显微镜下观察,通过绿色荧光蛋白基因在烟草叶片表皮细胞中的表达和定位分布情况对目的基因进行亚细胞定位分析。
2 结果与分析
2.1 目的基因全长序列的克隆
使用相应的引物进行RT-PCR(图3)、3 RACE(图4)和5 RACE(图5)扩增,将扩增得到的片段进行拼接得到全长cDNA,并在NCBI数据库中通过Blast比对,结果显示与数据库中存在的同一科的油棕的Catalases核酸序列具有高度的相似性,同源率分别为:ArCAT1与ElCAT1为95%,ArCAT2与ElCAT2为95%,ArCAT3与ElCAT3为96%,由此推断克隆所得的序列为槟榔的Catalases编码序列,分别命名为ArCAT1、ArCAT2、ArCAT3,长度均为1476 bp,编码492个氨基酸,并将所得序列在GenBank上登录,登录号分别为ArCAT1(MN692600)、ArCAT2(MN 692601)、ArCAT3(MN692602)。分析结果发现,ArCAT1与ArCAT2、ArCAT3的氨基酸序列同源率分别为79%和90%,ArCAT2与ArCAT3编码的氨基酸序列同源率为77%;ArCAT1与ArCAT2、ArCAT3的核酸序列同源率分别为76%和85%,ArCAT2与ArCAT3的核酸同源率为76%。
2.2 ArCATS基因的生物信息学分析
通过对槟榔过氧化氢酶蛋白序列的比对,利用氨基酸序列比对分析ArCAT1、ArCAT2和 ArCAT3编码的氨基酸序列(图6),结果发现ArCAT1、ArCAT2和ArCAT3均具有与定位有关的三肽(图6,box1、box2和box3)。
从图7可看出,ArCAT1、ArCAT2、ArCAT3分别与油棕的ElCAT1、ElCAT2、ElCAT3和海枣的PhCAT1、PhCAT2、PhCAT3聚在同一分支上,其中,ArCAT1、ArCAT3分别与ElCATI、ElCAT3聚在同一亚分支上,说明槟榔和油棕的亲缘关系最近。
2.3 亚细胞定位
利用http://www.csbio.sjtu.edu.cn/中的INSP预测真核蛋白中的核定位信号,在ArCAT2和ArCAT3中均发现了核定位信号的结构域(图6中的box4)。亚细胞定位结果发现(图8),无论GFP荧光蛋白位于基因的C端还是N端,对定位的结果均无影响。在ArCAT1、ArCAT2、ArCAT3中均可观测到绿色荧光点状物,这符合过氧化物酶体的定位特征。但ArCAT2和ArCAT3比较特殊,不仅定位在过氧化物酶体中,还定位在细胞核中,这与徐娟等[16]所报道的植物细胞Catalase主要分布在过氧化物酶体、乙醛酸循环体和细胞质中结论一致,说明ArCAT2和ArCAT3可能在细胞核和细胞质中均发挥重要的功能。
3 讨论
众多研究结果表明,过氧化氢是植物发育和环境反应的重要信号分子,在烟草中发现的3种过氧化氢酶中,CAT1负责清除光呼吸产生的H2O2,CAT2则清除氧化胁迫产生的H2O2,CAT3主要清除乙醛酸循环体中产生的H2O2[5]。玉米CAT1和CAT3在籽粒发育过程中表达,而CAT2的表达在籽粒发育后期才可检测到[17]。而这些过氧化氢蛋白酶主要存在于细胞质中的过氧化物酶体和乙醛酸循环体[18]。而本研究利用http://www.csbio.sjtu.edu.cn/预测真核蛋白中的核定位信号,在ArCAT2和ArCAT3中均发现了核定位信号的结构域(图6,box4),表明ArCAT2和ArCAT3定位在细胞核中并有可能发挥着重要的功能,这有待于作进一步的研究,预期对槟榔Catalase基因(ArCAT)功能的探究提供重要的思路。
在本研究中,研究发现ArCAT1和ArCAT2中存在经典的P-T-S1基序S/N-R-L基序,而在ArCAT3中则存在T-R-F基序(图6,box2),这些三肽并不位于末端的C端而是在上游-7至-9,这种内部定位的基序可能加强过氧化物酶体的导入,但其本身并不起作用。在N-R-L基序的上游是一个Q-K-L/I/V序列(图6,box1)。据报道[19-20],基因序列会影响它们之间的相互作用,基于目前对PTS1通路机制的了解,内部的Q-K-L序列似乎不太可能直接与PTS1受体蛋白Pex5p作用[21]。另一项研究得出结论,南瓜的过氧化物酶体中的CAT1积累需要PTS1通路机制[22]。虽然知道不同过氧化物酶体蛋白PTS1三肽表现出相当大的变异性,不过槟榔过氧化物酶的相应序列(图6,box3)不属于其列。尽管如此,研究发现为了将过氧化氢酶高效导入槟榔叶片细胞过氧化物酶体,C端三肽(图6,box3)是必需的,这一研究结论为后续槟榔过氧化氢酶基因高效遗传转化体系的建立提供了重要的科学依據。
参考文献
Loew O. A new enzyme of general occurrence in organismis[J]. Science, 1900, 11(279): 701-702.
宋新华, 赵凤云. 植物体内过氧化氢酶的研究进展[J]. 安徽农业科学, 2007(31): 9824-9827.
刘灵芝, 钟广蓉, 熊 莲, 等. 过氧化氢酶的研究与应用新进展[J]. 化学与生物工程, 2009, 26(3): 15-18.
刘云芬, 王薇薇, 祖艳侠, 等. 过氧化氢酶在植物抗逆中的研究进展[J]. 大麦与谷类科学, 2019, 36(1): 5-8.
王升平, 杨金广, 战徊旭, 等. 烟草过氧化氢酶基因CAT1的克隆及表达特征分析[J]. 中国烟草学报, 2014, 20(5): 103-109.
Frugoli J A, Zhong H H, Nuccio M L, et al. Catalase is encoded by a multigene family in Arabidopsis thaliana (L.) Heynh[J]. Plant Physiology, 1996, 112(1): 327-336.
Guan L Q, Scandalios J G. Molecular evolution of maize catalases and their relationship to other eukaryotic and prokaryotic catalases[J]. Journal of Molecular Evolution, 1996, 42(5): 570-579.
Esaka M, Yamada N, Kitabayashi M, et al. cDNA longing and differential gene expression of three catalases in pumpkin[J]. Plant molecular biology, 1997, 33(1): 141-155.
Iwamoto M, Higo H, Higo K. Differential diurnal expression of rice catalase genes: the 5-flanking region of CatA is not sufficient for circadian control[J]. Plant Science, 2000, 151(1): 39-46.
Willekens H, Chamnongpol S, Davey M, et al. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants[J]. EMBO Journal, 1997, 16(16): 4806-4816.
李曉娜, 曾小红, 谢龙莲, 等. 槟榔科学研究近况分析[J]. 热带农业科学, 2017, 37(3): 79-82.
易 攀, 汤嫣然, 周 芳, 等. 槟榔的化学成分和药理活性研究进展[J]. 中草药, 2019, 50(10): 2498-2504.
栾 剑, 陶晓月. 关于传统中药槟榔生物活性的研究进展[J]. 安徽农学通报, 2018, 24(13): 8-10, 12.
孙 娟, 曹立幸, 陈志强, 等. 中药槟榔及其主要成分的药理和毒理研究概述[J]. 广州中医药大学学报, 2018, 35(6): 1143-1146.
曾妍静, 沈文涛, 庹德财, 等. 适用于Nimble Cloning系统的pCambia载体改造[J]. 生命科学研究, 2019, 23(4): 276-280.
徐 娟, 常雁红, 罗 晖. 过氧化氢酶的研究和固定化及其应用进展[J]. 安徽农业科学, 2014, 42(34): 12035-12038.
Redinbaugh M G, Wadsworth G J, Scandalios J G. Characterization of catalase transcripts and their differential expression in maize[J]. Biochimica et Biophysica Acta, 1988, 951(1): 104-116.
Purev M, Kim Y J, Kim M K, et al. Isolation of a novel catalase (Cat1) gene from Panax ginseng and analysis of the response of this gene to various stresses[J]. Plant Physiology and Biochemistry, 2010, 48(6): 451-460.
Chen N, Teng X L, Xiao X G. Subcellular localization of a plant catalase-phenol oxidase, AcCATPO, from amaranthus and identification of a non-canonical peroxisome targeting signal[J]. Frontiers in Plant Science, 2017, 8: 1345.
Mhamdi A, Queval G, Chaouch S, et al. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models[J]. Journal of Experimental Botany, 2010, 61(15): 4197-4220.
Oshima Y, Kamigaki A, Nakamori C, et al. Plant catalase is imported into peroxisomes by Pex5p but is distinct from typical PTS1 import[J]. Plant Cell Physiol, 2008, 49(4): 671-677.
Lingner T, Kataya A R, Antonicelli G E, et al. Identification of novel plant peroxisomal targeting signals by a combination of machine learning methods and in vivo subcellular targeting analyses(W)[J]. Plant Cell, 2011, 23(4): 1556-1572.
猜你喜欢
人民周刊(2021年18期)2021-10-22
世界热带农业信息(2018年5期)2018-11-09
世界热带农业信息(2018年1期)2018-06-22
世界热带农业信息(2018年6期)2018-03-05
读书文摘(2017年10期)2017-10-16
山东农业科学(2016年11期)2016-12-17
椰城(2016年2期)2016-09-21
江苏农业科学(2015年5期)2015-10-20
