
粉色系月季杂交后代花色性状遗传分析
2020-02-22 02:54姜珊吴钰滢易星湾徐庭亮程堂仁王佳张启翔潘会堂
江苏农业学报 2020年6期
关键词:花色
姜珊 吴钰滢 易星湾 徐庭亮 程堂仁 王佳 张启翔 潘会堂


摘要:月季(Rosa spp.)是具有重要经济和观赏价值的植物,花色是月季最重要的观赏性状之一,解析月季花色的遗传规律可为月季花色改良提供依据。本研究为解析月季粉色花的遗传变异机制,以粉色系的窄叶藤本月季花(Rosa chinensis ‘Zhaiye Tengben Yuejihua)为母本、月月粉(Rosa chinensis ‘Old Blush)为父本杂交获得的F1群体为材料,利用色差仪对杂交群体的花色表型值进行测定,计算其色光值和花色分级值,并采用主基因+多基因混合遗传模型分析月季花色性状的遗传规律。结果表明,月季杂交群体花色出现从白色到暗红色的分离,利用ISCC-NBS色名表示法将其花色分为白色、粉色、红色、红紫色和墨色5个色系,并确定了各色系的参数分布范围,建立了杂交群体色系与色差仪CIELab表色系统明度(L*)、红度(a*)和黄度(b*)的对应关系;此外,确定该月季杂交群体花色受1对加性-显性主基因控制,遗传模型为1MG-AD,利用色光值和花色分级值2个参数计算的花色主基因遗传率较高,分别为81.92%和96.86%,不易受环境影响。研究结果可为月季花色育种提供参考。
关键词:月季;花色;表型变异;遗传分析
中图分类号:S685.12文献标识码:A文章编号:1000-4440(2020)06-1529-08
Abstract:Roses (Rosa spp.) are important economic and ornamental plants with flower color being one of the most important ornamental traits. Elucidating the genetic rule of rose color can provide a foundation for rose color improvement. To study the genetic variation mechanism of pink rose flowers, the F1 population material was got by hybridization, taking pink Rosa chinensis ‘Zhaiye Tengben Yuejihua as the female parent and pink Rosa chinensis 'Old Blush' as the male parent. The phenotypic value of flower color of hybrid population was determined by colorimeter, the color-light value and color classification value were calculated. The genetic rule of flower color traits of roses was analyzed by using mixed genetic model of major genes plus multi-genes. The results showed that the colors of the hybrid rose population showed segregation from white to dark red, divided into five color systems such as white, pink, red, red-purple and dark red, using Inter-Society Color Council-National Bureau of Standards (ISCC-NBS), and the distribution ranges of parameters in all color systems were determined. The relationship between color system and luminosity (L*), redness (a*), yellowness(b*) detected by CIELab of the hybrid population was established. In addition, it was confirmed that the flower color of the hybrid rose population was controlled by a pair of additive-dominant master gene, and the genetic model was 1MG-AD. The heritabilities of major flower color genes calculated by two parameters such as color-light value and color classification value were high, which was 81.92% and 96.86% respectively, and it was not easy to be influenced by the environment. The results can provide reference for flower color targeted rose breeding.
Key words:Rosa spp.;flower color;phenotypic variation;genetic analysis
月季屬于蔷薇科(Rosaceae)蔷薇属(Rosa L.),是世界上最具观赏价值的植物之一,已有3 000多年的栽培历史。目前全世界约有30 000~35 000个栽培月季品种,长期的种间杂交使月季具有丰富艳丽的色彩。研究月季花色的遗传规律有助于了解月季的选育过程,对提高月季品质以及培育不同色系的月季新品种具有重要意义[1]。
当前针对月季花色形成机制的研究多集中于色素种类及生物合成代谢途径等方面。研究发现,类胡萝卜素及类黄酮中花青苷和黄酮醇相对含量的差异和色素之间的相互作用使月季呈现不同的花色[2]。红色花或粉色花的呈色物质主要是花青苷,黄色花的呈色物质为类胡萝卜素,橙色花通常同时含有以上两类色素[3-5]。在花青素合成代谢途径研究中,人们发现了天竺葵素、矢车菊素和芍药素这3种主要的花青素,这些化合物性质主要通过糖基化作用得到稳定,对花青素糖基化水平的调控决定细胞中花青苷的积累[4,6-7],对花青素代谢途径的调控大部分处于转录水平,其表达主要受结构基因(CHS、PAL等)和转录因子(MYB、WD40、bHLH 等)的影响[8-9]。
月季遗传组成非常复杂、倍性水平差异较大、花青素合成代谢途径复杂,加上有性繁殖困难,其花色的遗传规律尚不明确[10-11]。研究发现,适宜的遗传群体是探究月季花色遗传规律的必要条件。Marshall 等[12]对47个群体1 200多个后代进行花青素色素分析,发现花色表现为数量遗传。粉色是野生蔷薇属植物和栽培月季品种中分布最广泛的颜色 [13]。因此,解析月季粉色花的遗传变异机制对月季花色育种具有重要意义。Debener等[14]在R. multiflora 93/1-117(粉色)×93/1-119(粉色)杂交群体研究中发现,杂交后代出现白色∶粉色为1∶3的分离比,并将控制月季粉色性状的1个位点(Blfa)定位在IM-LG2上,认为月季粉色性状受单个显性基因或1对显性互补基因控制,但其未记录杂交后代粉色和红色以及不同程度的粉色之间的差异。Shupert等[15]利用3个回交群体研究了月季白色/粉色性状的遗传规律,发现粉色由1个主要的共显性基因控制。Henz等[16]和Gitonga等[17]则均选用粉色和红色月季进行杂交,通过遗传定位分析,发现了一些控制月季花色的数量性状位点。尽管前人对粉色系月季进行了相关研究,但由于受到杂交群体数量的限制,同时缺乏对杂交后代花色的精细测量和分类,月季花色的遗传变异规律仍未明确。鉴于CIELab颜色系统可以准确反映物质颜色,且和色素组成直接相关[18-19],花色的数量化已经成为研究花色分类及遗传的重要内容。近年来,在植物数量性状的遗传研究中,主基因+多基因混合遗传模型已被广泛应用于分析植物的多种农艺性状[20-22],可根据不同性状的不同基因效应,选择适合的育种方法,为性状选育提供参考依据[23-25],而在月季的研究中,较少采用遗传模型。
为阐明粉色系月季花色的遗传机制,本研究首先将粉色系的窄叶藤本月季花与粉色系的月月粉进行杂交,杂交群体出现了从白色到暗红色的显著分离;其次,测定杂交群体的花色表型并进行分类;最后,采用主基因+多基因混合遗传模型对月季花色性状的遗传规律进行相关分析研究。研究目的是解析粉色系月季遗传变异规律,为月季花色机理研究及花色遗传分析提供参考依据。
1材料与方法
1.1试验材料
杂交母本为窄叶藤本月季花(Rosa chinensis ‘Zhaiye Tengben Yuejihua)[26],父本为月月粉(R. chinensis ‘Old Blush),二者均为月月粉类品种[27],均为粉色重瓣花。2015年4-8月在国家花卉工程技术研究中心小汤山基地进行杂交,花粉贮藏与杂交方法、种子处理和杂种苗养护管理参见《中国月季》[1]。
1.2颜色测定
2018年和2019年的6-8月,连续2年用英国皇家园艺学会的标准比色卡(Royal Horticultural Society Color Chart,RHSCC)和色差仪(NF555,Nippon Denshoku Industries Co. Ltd.,Japan)对杂交亲本以及F1代163个株系的花色进行测量。每个株系选取3朵刚刚充分开放(花药裂开)的花,每朵选取3个花瓣进行测定,若是重瓣花则取中轮花瓣。在C/2°光源条件下,测量花瓣正面中间部位的颜色,测得CIELab值以及孟赛尔颜色系统(Munsell color system)的参数值。
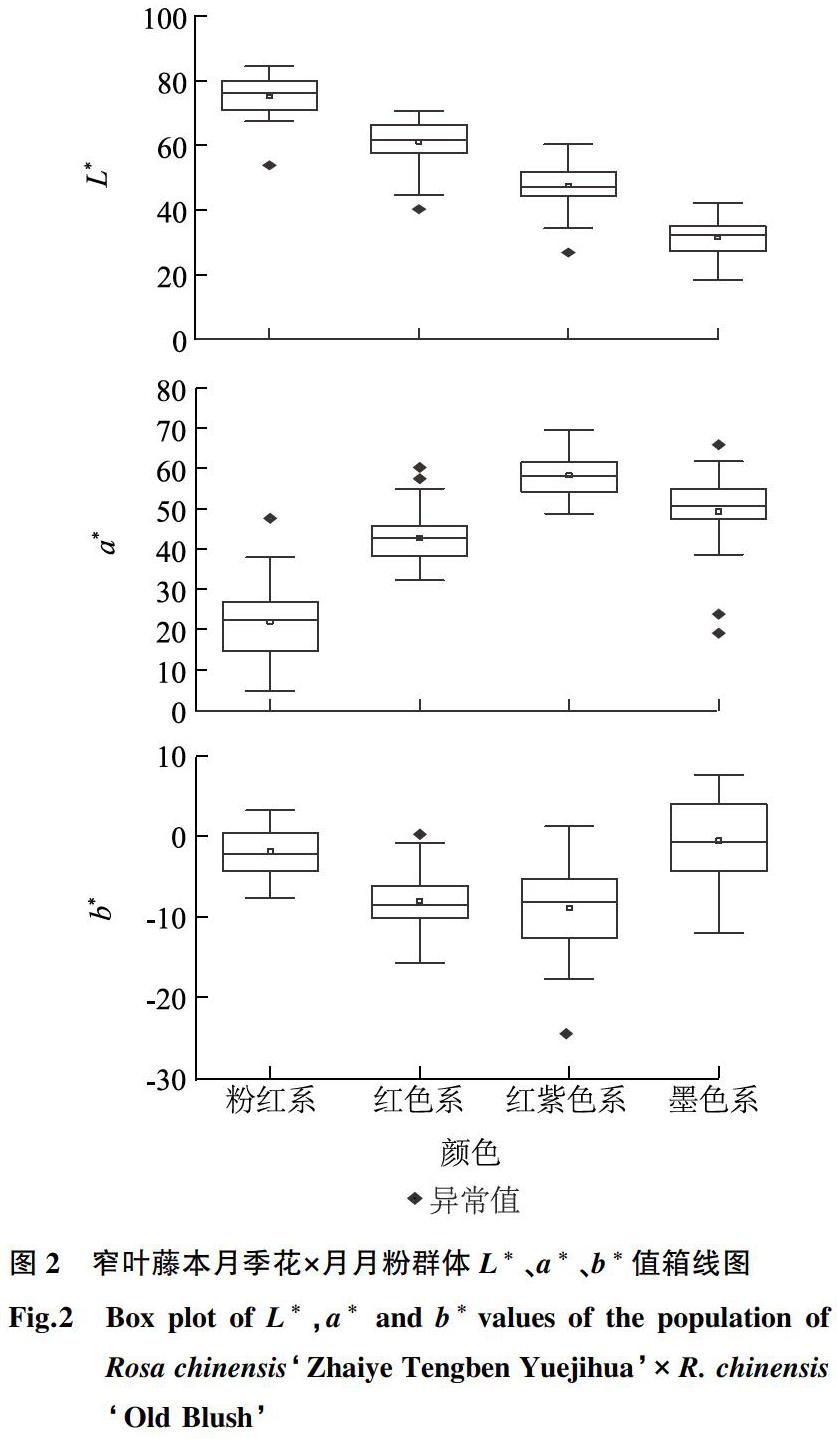
对每个株系的9次测量数据取平均值,并计算主要反映物质表面颜色的色光值[28],色光值=2 000×a/L×(a2+b2)1/2。根据色差仪测得的孟塞尔颜色系统参数色调(H)、明度(V)和彩度(C),采用 ISCC-NBS 命名法,先在第3等级给所有样本命名颜色,然后简化明度和饱和度的描述,归类为第1等级的色系[29-30]。利用 Origin 2019软件绘制各个色系的 L*、a*和b*箱线图及二维、三维分布图。
1.3主基因+多基因遗传分析
采用盖钧镒等[23]提出的主基因+多基因混合遗传模型,并参考马杰等[31]的方法,采用F2群体的模式分析月季群体花色性状的遗传方式。利用SEA软件统计11种模型的AIC值(Akaikes information criterion),选择最优遗传模型[24,32],并进行遗传参数估算。
2结果与分析
2.1窄叶藤本月季花×月月粉群体花色表型遗传变异
对杂交亲本以及杂交获得的完成开花过程且处于同一生长状态的F1代163個株系进行花色分析。结果显示,父本月月粉(RHS:68-C)和母本窄叶藤本月季花(RHS:67-D)均为粉色系品种,但其杂交后代出现从白色到暗红色的花色分离(图1)。
2.1.1基于ISCC-NBS色名表示方法对群体花色表型进行命名采用 ISCC-NBS色名表示法对163个杂交后代株系的花色进行命名和分类,将群体花色命名为22 种颜色类型(表1),杂交后代花色以粉色、红色、红紫色居多,其中鲜红偏紫色(Vivid purplish red)个体最多,占总数的21.47%,其次为强红偏紫(Strong purplish red),占总数的12.27%。
2.1.2窄叶藤本月季花×月月粉群体花色表型变异特点参考洪艳等[30]的分类方法,将H值较低的一类归为墨色系(Dark red group),同时考虑到实际应用习惯与经验,将唯一的白偏黄(Yellowish white)后代单独归为白色系。最终,将163个后代划分为白色系、粉色系(Pink group)、红色系(Red group)、红紫色系(Red purple group)、墨色系5个色系。通过绘制箱线图,对所划分的5个色系进行验证和评价。对比L*、a*、b*值的分布特点,发现各色系参数分布具有一定的规律(图2),能科学地将不同色系进行区分。L*值由高到低依次是粉色系、红色系、红紫色系和墨色系,4个色系基本无重叠。其中墨色系的L*值分布集中,红色系、红紫色系和墨色系的a*值基本集中在30~60,其中红紫色系的a*值最大,粉色系的a*值分布最广泛。红紫色系的b*值最小,红色系和红紫色系的b*值均匀分布在负值区间内,粉色系和墨色系的 b*值在正负区间内都有分布。墨色系的 b*值分布范围较广,但是L*值和a*值分布集中。粉色系的a*值低于其他色系,而 L*值和 b*值均高于其他色系。白色系仅1株,未达到箱线图作图要求,不放在图中。
通过分析和比较各个色系L*、a*和 b*值的分布,发现群体后代花色分离广泛。在a*、b*值二维色相坐标系中(图3A),5个色系集中在Ⅱ、Ⅳ象限,Ⅰ、Ⅲ象限基本无分布,其中在Ⅳ象限的分布最为集中、数量最多;白色系和粉色系分布位置较接近原点,红色系和红紫色系主要分布在Ⅳ象限,墨色系广泛分布于Ⅱ和Ⅳ两象限。在 L*、a*、b*值三维坐标系(图3B)中,各色系呈三维带状分布,L*值分布范围为18.43~84.99,a*值分布范围为2.27~69.52,b*值分布范围为-24.71~7.69。
通过采用箱线图和 L*、a*、b*参数分布图的分类、分析及验证,表明ISCC-NBS色名表示法对花色的定义和分类结果准确。由此,基本建立了划定群体色系与色差仪测色空间CIELab表色系统 L*、a*、b*值的对应关系,每种花色类型都有其对应的参数分布范围(表2、表3),能够定量描述和分析群体花色的分离特点。
2.2采用主基因+多基因模型对窄叶藤本月季花×月月粉杂交群体花色的遗传分析
2.2.1最优模型的选择和检验参考前人对果蔬颜色进行主基因+多基因遗传分析的方法[33-34],将本研究分类所得5个月季色系定为不同级别的数值,用花色分级数值和色光值2个参数对窄叶藤本月季花×月月粉群体的花色进行分析。分别选出AIC值最小的2个模型作为候选模型,色光值的2个候选模型(1MG-AD、2MG-A)均未达到显著水平,根据AIC值最小原则选择1MG-AD作为最佳模型。花色分级数值的2个候选模型(1MG-AD、2MG-AD)达到显著水平的参数个数相同(表4、表5),通过比较AIC值确定最优模型为1MG-AD,因此本研究得出月季花色最优遗传模型为1MG-AD,受1对加性-显性主基因控制。
2.2.2遗传效应值的估算采用最小二乘法估算群体花色的一阶和二阶参数(表6),有|da|>|ha|的关系,说明月季花色主基因的加性效应大于显性效应,色光值和花色分级值的主基因遗传率分别为81.92%和96.86%,花色主基因遗传率较大,属于高等遗传率,该性状不易受环境影响。
3讨论
3.1粉色系月季杂交群体花色的分离
月季种类丰富,色彩缤纷,对研究花色遗传具有重要价值,是不可或缺的良好材料。本研究使用的月季杂交亲本均为粉色,但杂交群体花色分离广泛,出现了不同于亲本的花色,表明亲本杂合度高、遗传信息丰富。通过ISCC-NBS色名表示法将群体花色分成白色、粉色、红色、红紫色、墨色系5类,笔者整理了各色系的参数分布范围,建立了该群体色系与色差仪CIELab表色系统 L*、a*、b*值的对应关系,实现了对花色分离特点的定量描述,便于区分每个个体之间的差异。与前人研究结果[14,35-36]相比,本研究群体花色分离更广泛,为群体遗传学、花色分类、花色形成机理、新品种培育等研究提供了重要依据。5个色系的 L*、a*、b*值分布区间与大小关系,与前人的研究结果相似[10,37]。在a*、b*二维分布图中,Ⅰ象限无后代分布,是因为该群体中无黄色系、橙色系个体,推测群体花瓣色素种类中没有类胡萝卜素;而Ⅲ象限没有个体分布,是因为月季缺少类黄酮 3,5-羟化酶,缺少合成蓝色的飞燕草色素[38]。汪有良等[39]通过分析月季品种与其亲本花色的相关性发现,粉色类型的月季品种花色受亲本影响较小,与本研究中亲本均为粉色而杂交后代出现从白色到暗紅色的花色分离现象的结果一致。杂交后代出现不同程度的“粉色”,说明粉色是分析花色数量遗传的优良性状,后续可通过测定杂种后代的色素含量来深入研究花色遗传变异机制。
3.2月季花色的主基因+多基因混合遗传模型分析
基因组高度杂合的植物,杂交获得的F1代相当于作物纯合亲本杂交获得的F2代,因此常用F1代进行遗传模型分析[31,40]。利用F2群体主基因+多基因混合遗传模型,杨信程等[40]分析了切花小菊(Chrysanthemum morifolium)F1代的分枝性状;马杰等[31]分析了菊花F1代舌状花的半致死温度。在对颜色的遗传分析中,王建科等[41]利用色差仪测得的L*值和C*值对黄瓜(Cucumis sativus)嫩果皮颜色进行主基因+多基因遗传分析;申晓青等[33]根据色差仪测定值,对黄瓜嫩果皮色进行分类和分级,再进行果色遗传分析。色差仪所测得的每个颜色的数据都有对应的L*、a*和 b*等参数值,颜色的差异无法用其中任何一个单一数值表示,而由 L*、a*和 b*值计算而来的色光值可以综合体现物质表面颜色和明亮度,在花青素含量分析及花色遗传分析中应用广泛[28,34]。本研究通过综合选用色光值和颜色分级数值进行花色遗传模型分析,提高了试验结果的准确性及可靠性。经分析,确定月季花色的遗传模型为1MG-AD,受1对加性-显性主基因控制,且控制花色的主基因遗传率高,受环境影响小[40,42],与前人研究得出的月季花色变异受单基因和数量性状位点控制、环境产生的效应相对较小的结论类似[14,16,35-36]。月季花色主基因的加性效应大于显性效应,在栽培早期适当提高选择标准,遗传改良较易取得效果。本研究鉴定了月季花色的遗传模式,并分析了主基因的遗传效应,为月季花色选育提供了有益信息。
[18]CONESA A , MANERA F C , BROTONS J M , et al. Changes in the content of chlorophylls and carotenoids in the rind of Fino 49 lemons during maturation and their relationship with parameters from the CIELAB color space[J]. Scientia Horticulturae, 2019, 243:252-260.
[19]WAN H H, YU C, HAN Y,et al. Determination of flavonoids and carotenoids and their contributions to various colors of rose cultivars (Rosa spp.)[J]. Frontiers in Plant Science, 2019,10:123.
[20]YE Y J, WU J Y, FENG L, et al. Heritability and gene effects for plant architecture traits of crape myrtle using major gene plus polygene inheritance analysis[J]. Scientia Horticulturae, 2017, 225:335-342.
[21]SUN X R, LIU L, ZHI X N, et al. Genetic analysis of tomato internode length via mixed major gene plus polygene inheritance model[J]. Scientia Horticulturae, 2019, 246:759-764.
[22]FAN Z P, GAO Y K, LIU R, et al.The major gene and polygene effects of ornamental traits in bearded iris ( Iris germanica ) using joint segregation analysis[J]. Scientia Horticulturae,2020,260:108882.
[23]蓋钧镒,章元明,王建康. 植物数量性状遗传体系[M]. 北京:科学出版社,2003:26-56.
[24]曹锡文,刘兵,章元明. 植物数量性状分离分析Windows软件包SEA的研制[J]. 南京农业大学学报,2013,36(6):1-6.
[25]章元明. 植物数量遗传学的建立、发展与展望[J]. 南京农业大学学报,2012,35(5):19-24.
[26]王国良.中国古老月季[M]. 北京:科学出版社,2005:160-178.
[27]TAN J R,WANG J,LUO L,et al. Genetic relationships and evolution of old Chinese garden roses based on SSRs and chromosome diversity[J]. Scientific Reports,2017,7(1):15437.
[28]周蓉,蒋芳玲,梁梅,等.用色差仪法定量分析番茄果实番茄红素的含量[J]. 江西农业学报,2012,24(9):45-48,50.
[29]KELLY K L,JUDD D B. Color:universal language and dictionary of names[M]. New York:US Department of Commerce,National Bureau of Standards,1976:37-158.
[30]洪艳,白新祥,孙卫,等. 菊花品种花色表型数量分类研究[J]. 园艺学报,2012,39 (7):1330-1340.
[31]马杰,徐婷婷,苏江硕,等. 菊花F1代舌状花耐寒性遗传变异与QTL定位[J]. 园艺学报,2018,45(4):717-724.
[32]CAI C C,TU J X,FU T D. The genetic basis of flowering time and photoperiod sensitivity in rapeseed Brassica napus L.[J]. Russian Journal of Genetics,2008,44(3):326-333.
[33]申晓青,陈书霞,潘玉朋,等. 黄瓜嫩果果皮颜色的遗传研究[J]. 农业生物技术学报,2014,22(1):37-46.
[34]吴浪,刘婧仪,梁燕. 番茄绿果与红果颜色性状遗传的研究[J]. 园艺学报,2016,43(4):674-682.
[35]YAN Z,DENNEBOOM C,HATTENDORF A,et al. Construction of an integrated map of rose with AFLP,SSR,PK,RGA,RFLP,SCAR and morphological markers[J]. Theoretical and Applied Genetics,2005,110(4):766-777.
[36]SPILLER M,LINDE M,HIBRAND S O L,et al. Towards a unified genetic map for diploid roses[J]. Theoretical and Applied Genetics,2011,122 (3):489-500.
[37]于超. 四倍体月季遗传连锁图谱的构建及部分观赏性状的QTLs分析[D]. 北京:北京林业大学,2015.
[38]HOLTON T A,TANAKA Y. Blue roses - a pigment of our imagination? [J]. Trends in Biotechnology,1994,12(2):40-42.
[39]汪有良,孙海楠. 月季品种与其亲本的花色相关性分析[J]. 江苏林业科技,2016,43(5):32-36.
[40]杨信程,苏江硕,吴洋洋,等. 切花小菊主要分枝性状的混合遗传分析[J]. 南京农业大学报,2018,41(3):440-446.
[41]王建科,方小雪,李雪红,等. 黄瓜嫩果皮颜色的遗传研究[J]. 园艺学报,2013,40(3):479-486.
[42]顾万春. 统计遗传学[M]. 北京:科学出版社,2004:241-321.
[43]金亮,包劲松. 植物性状-标记关联分析研究进展[J]. 分子植物育种,2009,7 (6):1048-1063.
[44]徐剑文,孔杰,赵君,等. 盐胁迫下棉花萌发、成苗和产量相关性状的QTL定位[J].江苏农业学报,2018,34(5):972-977.
[45]俎峰,赵凯琴,张云云,等. 甘蓝型油菜的花期与生育期QTL定位[J].南方农业学报,2019,50(3):500-505.
(责任编辑:陈海霞)
猜你喜欢
作文大王·笑话大王(2021年4期)2021-04-26
课堂内外(高中版)(2021年7期)2021-01-17
初中生学习指导·提升版(2020年10期)2020-09-10
阅读与作文(初中版)(2020年3期)2020-03-02
环球时报(2020-02-19)2020-02-19
孩子(2020年12期)2020-01-06
东坡赤壁诗词(2019年5期)2019-11-14
学生天地·初中(2018年6期)2018-07-08
饮食与健康·下旬刊(2018年3期)2018-04-11
食品界(2017年10期)2017-11-16
