
鲤(Cyprinus carpio)对稻田水体浮游植物及环境因子的影响研究
2020-04-11 04:15彭辉辉张凯陈成勋谢骏李志斐郁二蒙刘龙飞龚望宝田晶晶王广军通信作者
天津农学院学报 2020年1期
彭辉辉,张凯,陈成勋,谢骏,李志斐,郁二蒙,刘龙飞,龚望宝,田晶晶,王广军,3,通信作者
(1.天津农学院 水产学院,天津 300384;2.中国水产科学研究院 珠江水产研究所 农业农村部热带亚热带水产资源利用与养殖重点实验室,广州 510380;3.广东省水产养殖污染修复生态工程技术研究中心,广州 510380)
稻田养鱼指在种植水稻的同时利用稻田水环境养殖水产品,是一种兼顾经济效益和生态效益的劳作方式。相比水稻单种模式,稻田养鱼可通过鱼的代谢将N、P等元素转化为水稻易吸收的形式,可减少化肥使用量,进而降低施肥所带来的农业面源污染[1]。同时,由于鱼类的活动及摄食,减少了水稻虫害和病害的发生,降低了杀虫剂等农药的施放量[2]。此外,稻田养鱼在增加或保持水稻产量的前提下,收获了优质水产品,更立体地利用了稻田资源[3],在淡水和耕地资源日益紧张的今天,稻田养鱼有着广阔的发展前景。
浮游植物(Phytoplankton)作为水体生态系统的主要生物群落之一,在物质循环和能量流动中起着重要作用[4]。浮游植物作为系统的生产者,为其他生物提供能量、氧,吸收固持营养元素,增加土壤有机质含量,改善土壤物理性状且提高土壤肥力,促进水稻生长[5]。浮游植物还是水体富营养化的指示生物,其种类丰度、群落结构能反映水体富营养化程度,其生长繁殖也受外界环境因素如营养盐、有机物浓度、其他生物和人为活动的影响[6]。
作为水产动物的饵料,浮游植物与水体环境因子也有着密切的关系[7]。对养鱼稻田水体浮游植物进行研究,有利于了解养鱼稻田的水质和饵料情况,在控制稻田水质、提高稻田养鱼技术方面有一定的指导意义。目前,对稻鱼系统浮游植物的研究已逐步展开,有学者对稻鲫[8]、稻虾[9]、稻蟹[10]模式的浮游植物进行过研究,但对稻鲤模式浮游植物及其与环境因子关系的研究尚较少。
华南地区养鱼稻田多分布于北部山区,由于地形及历史原因,其稻田养鱼基础数据较为薄弱[11]。本研究通过田间试验对华南地区养鱼稻田和单种稻稻田的浮游植物群落结构及生物量进行分析,并对二者水体环境因子进行测定,旨在明确华南地区养鱼稻田与单种稻稻田浮游植物群落结构的差异,为稻田养鱼提供基础数据。
1 材料与方法
1.1 试验地点选择
试验在广东省清远市连南瑶族自治县进行,试验稻田数量6块,面积均为0.20 hm2,长宽比约为4∶3。6块稻田水源一致,其中试验组(RF)稻田3块,种稻并放养当地鲤鱼苗,对照组(RM)稻田3块,单种稻不养鱼。鲤鱼苗为当地的土著鲤鱼,俗称“禾花鱼”,当地传统养鱼密度为 112.5 kg/hm2,每块试验田放鱼苗900尾。试验期间,稻田水体深度保持在15~20 cm,养鱼稻田有人工开挖对鱼起保护作用的鱼涵,鱼涵呈长方形,深1 m,位于稻田正中央,面积约占稻田总面积的5%,以防水位降低对鱼造成影响。整个试验期不施肥、不喷农药、不投饵,鱼主要摄食稻田中的天然饵料。
1.2 采样方法与指标测定
采用YSI多功能水质分析仪现场测定水体温度、溶解氧、电导率、pH值、溶解性固体总量等指标,按照国标法测定水体亚硝酸盐、硝酸盐、磷酸盐、总氮、总磷和总有机碳含量[12],底泥环境因子指标采用郑必昭[13]的方法进行测定。
样品采集方式为五点采样法,即在稻田的四角和中心采样。浮游植物样品的定量采集方法为:取0.5 L水样,加入5 mL鲁哥氏液固定后,倒入有刻度的沉淀器定容,静置24~48 h后,用虹吸管吸取上清液,浓缩定容至30 mL时把沉淀液倒入塑料瓶再进行鉴定。浮游植物的定性和定量测定参考韩茂森等[14]、胡鸿钧等[15]、赵文[4]的方法,在光学显微镜下对浮游植物的种类和密度进行鉴定。
根据浮游植物不同物种的出现频率(fi)和个体数量(Ni)与总数量(N)的比例来计算优势度Y,当Y≥ 0.02时,确定为优势种,计算公式为:

采用Shannon-Weiner 多样性指数(H)描述两种模式浮游植物的多样性,计算公式为:

用Simpson指数(D1)描述样本群体的均匀度,即连续两次抽取样本,其中两次抽到不同种的概率,其计算公式为:

用 Margalef指数(D2)描述浮游植物物种的多寡性,计算公式为:

式中,s为浮游植物的总物种数,N为样本总个体数,Pi为第i种浮游植物所占总数的比例,即为抽取1次抽到i物种的概率。
1.3 数据分析
采用Excel 2013对试验数据进行整理作图,用SPSS 21.0软件进行统计分析,以独立样本T检验分析两组数据的差异显著性,数据结果以“平均值±标准差”表示,以P<0.05作为差异显著性水平,P<0.01作为差异极显著水平。采用Canoco 5.0软件,对两组稻田具有显著性差异的水体和底泥的理化因子与浮游植物相关性进行冗余分析(RDA)。
2 结果与分析
2.1 试验组与对照组环境因子的比较
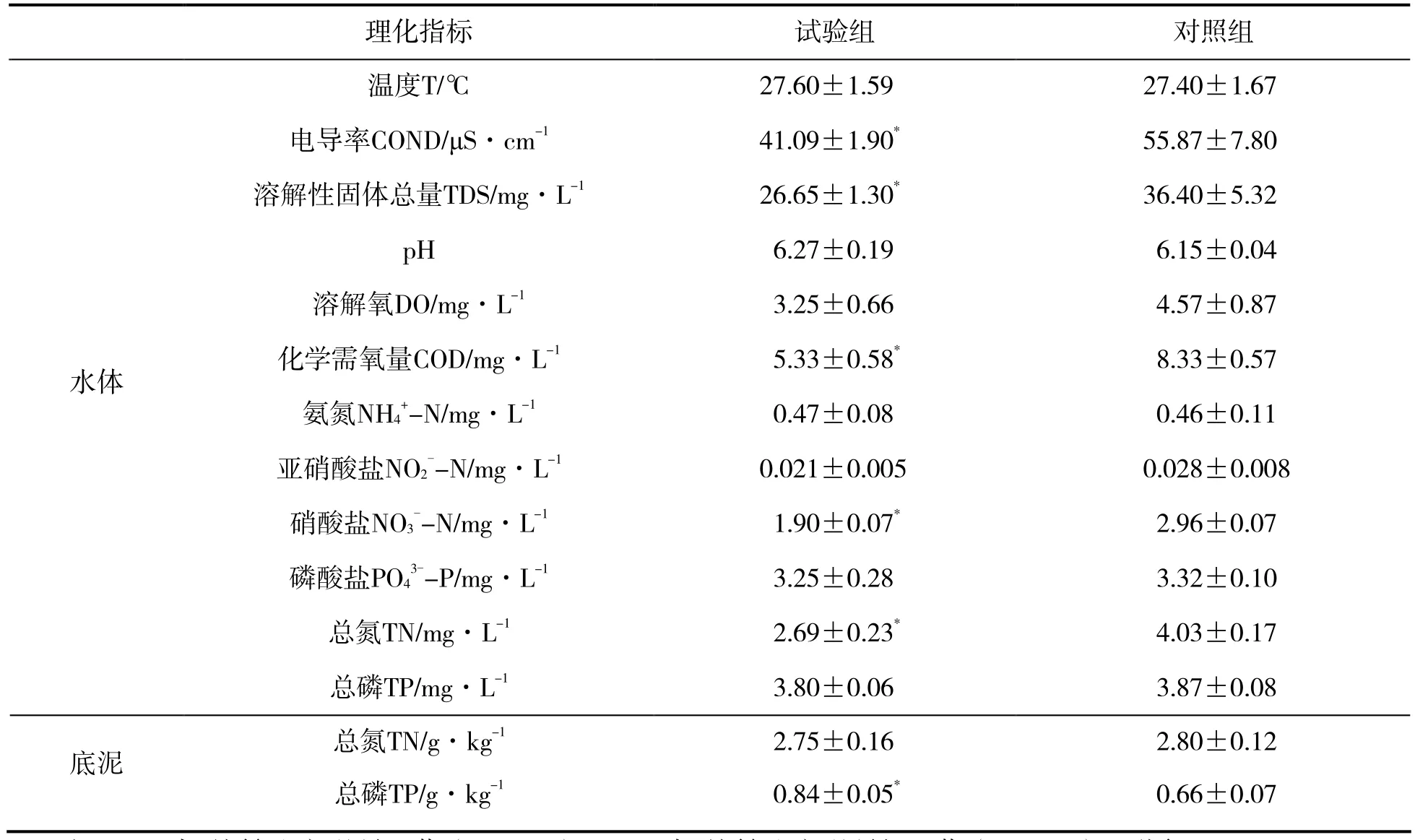
表1 两组稻田环境因子比较
2.2 浮游植物的密度和生物量
两种养殖模式浮游植物的密度和生物量如图1所示,试验组浮游植物密度和生物量为(66.95±7.50)×105ind/L和(5.98±0.63)mg/L,对照组则为(35.82±4.30)×105ind/L和(3.32±0.45)mg/L,试验组的浮游植物生物量和密度均极显著高于对照组(P<0.01)。

图1 试验组(RF)和对照组(RM)浮游植物密度(A)和生物量(B)
两种模式浮游植物各门类密度如图2所示。对照组藻类主要隶属于 5个门类,包括绿藻门(Chlorophyta)、蓝藻门(Cyanophyta)、硅藻门(Bacillariophyta)、裸藻门(Euglenophyta)和隐藻门(Cryptophyta),试验组藻类主要隶属于绿藻门、蓝藻门、硅藻门、裸藻门、隐藻门、金藻门(Chrysophyta)和甲藻门(Pyrrophyta)7个门,试验组丰度以绿藻门和硅藻门为主,其密度分别为(26.00±3.47)×105ind/L和(13.56±2.13)×105ind/L;对照组以硅藻门和蓝藻门为主,其密度分别为(15.12±2.84)×105ind/L和(10.70±1.84)×105ind/L。不同模式浮游植物各门密度差异显著,其中,试验组的蓝藻门密度显著低于对照组(P<0.05),试验组的绿藻门、裸藻门、隐藻门密度均显著高于对照组(P<0.05),两种模式硅藻门密度无显著差异(P>0.05)。
大多数民办高校教工党支部的组织生活仍然停留于传达上级的有关精神内容和完成上级的有关布置任务,组织生活活动方法陈旧或形式化,内容单一,缺乏创新意识,在民主生活会上的批评和自我批评时常流于形式而难出实际效果,这些都在一定程度上削弱了教工党支部的政治影响力。

图2 试验组(RF)和对照组(RM)浮游植物各门丰度对比
2.3 两种模式浮游植物密度和生物量百分比
两种模式浮游植物密度如图3(A)所示,试验组中浮游植物以绿藻门、硅藻门和隐藻门为主,其中绿藻门 6种,密度占浮游植物总密度的(39.33±1.25)%,硅藻门5种,占(19.89±1.19)%,隐藻门共1种,占(14.46±0.70)%;对照组中浮游植物以硅藻门、蓝藻门和绿藻门为主,其中硅藻门7种,占比为(42.09±1.75)%,蓝藻门6种,占比为(29.91±1.59)%,绿藻门 9种,占比为(25.35±0.95)%。两种模式浮游藻类生物量如图3(B)所示,试验组中浮游植物生物量以绿藻门、裸藻门为主,绿藻门生物量占浮游植物总生物量比例为(45.75±2.30)%,裸藻门占(34.67±2.37)%;对照组生物量以绿藻门和硅藻门为主,其中绿藻门生物量占比为(51.74±5.53)%,硅藻门占比为(41.33±6.44)%。
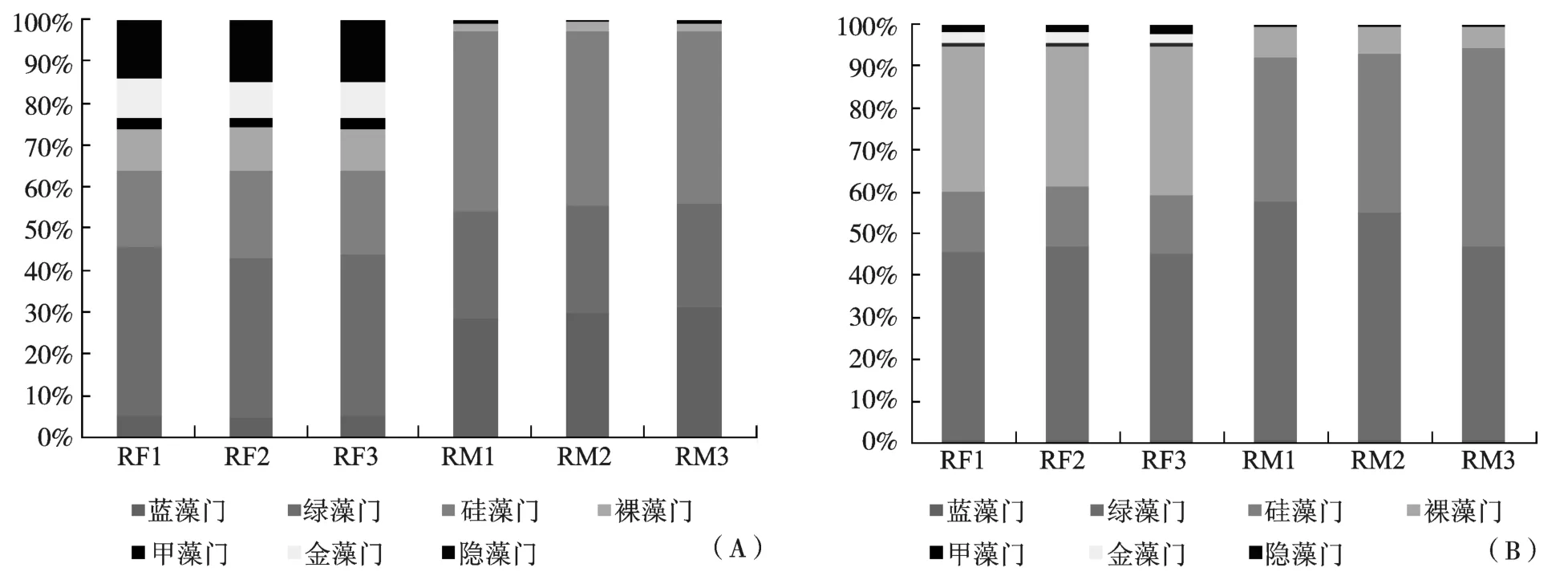
图3 两组稻田浮游植物各门丰度(A)和生物量(B)百分比占比
2.4 两组稻田养殖期间浮游植物优势种比较
如表2所示,以优势度Y≥0.02作为优势种,其中,试验组共记录到11种,分别为蓝藻门的阿氏浮丝藻(Planktothrix agardhii),绿藻门的卷曲纤维藻(Ankistrodesmus convolutus)、双对栅藻(Scenedesmus bijuga)、球衣藻(Chlamydomonas globosa),硅藻门的钝脆杆藻(F.capucina)、大羽纹藻(Pinnularia major)、缘花舟形藻(Navicula radiosa),裸藻门的绿裸藻(Euglena viridis),甲藻门的裸甲藻(Gymnodinium uberrimum),金藻门的锥囊藻(Dinobryon divergens)和隐藻门的卵形隐藻(C.ovata);对照组有蓝藻门的假鱼腥藻(Pseudanabaena schmidlei)、优美平裂藻(Merismopedia elegans)、微小隐球藻(Aphanocapsa delicatissima)、铜绿微囊藻(M.aeruginosa)、泽丝藻(Limnothrix redekei)、齿牙栅藻(Scenedesmus denticulatus),绿藻门的单针藻(Monoraphidium komarkovae)、双对栅藻,硅藻门的弯曲菱形藻(Nitzschia sigmoide)、矮小曲壳藻(Achnanthes exigua)、大羽纹藻、缘花舟形藻。相对于对照组,试验组优势种分布门类更广,遍及金藻门、裸藻门、甲藻门和隐藻门,集中于绿藻门和硅藻门,而单种稻系统主要集中在蓝藻门。

表2 试验组与对照组水体浮游植物的优势种优势度
2.5 试验组和对照组浮游植物多样性指数
如表 3所示,试验组水体浮游植物的Shannon-Weiner多样性指数和Margalef指数显著低于对照组(P<0.05),二者的辛普森多样性指数无显著差异(P>0.05)。

表3 两组稻田浮游植物多样性指数
2.6 水体和底泥理化因子与浮游植物丰度的相关性分析
环境因子与浮游生物的RDA分析见图4,水体硝酸盐浓度(NO3--N)、总氮(W-TN)和COD与蓝藻门的丰度正相关,与绿藻门、金藻门、隐藻门的丰度呈显著负相关关系,底泥总磷(S-TP)与金藻门、隐藻门、裸藻门、甲藻门和绿藻门丰度呈正相关关系。

图4 环境因子与浮游植物各门丰度的RDA冗余分析
3 讨论
3.1 水体和底泥环境因子
水产养殖活动一般会对养殖系统及周边环境造成一定的影响。本研究中,试验组水体的浓度与对照组均无显著差异(P>0.05),这与丁伟华等对稻鱼系统水环境的研究结果一致[16],说明稻田养鲤并未对水体环境造成不良影响,其原因可能与稻田养鱼密度较低,且未人工投饵有关[17]。本研究中,试验组水体的总氮浓度显著低于对照组(P<0.05),这说明鲤的加入对水体氮素去除具有一定作用。溶解性固体为水体中的细微悬浮颗粒,电导率可反映水体的离子强度,COD是水体还原性物质的含量,三者皆为判断水质的重要指标[18]。本研究试验组水体的溶解性固体总量、电导率、COD均低于对照组,说明稻田养鲤可显著改善水质,其原因可能为稻田鲤主要摄食天然饵料,促进了环境氮素向生物体转化,故能净化水质,减少稻田水体污染[19]。磷是水稻生长过程中必需的营养元素,本研究试验组底泥的总磷(STP)含量显著高于对照组(P<0.05),表明养鲤稻田底质得到了一定的改善,原因可能是鲤的排泄物沉积到底泥,促进了底泥磷元素的富集。
本研究中,鲤鱼初始均重为(25.34±3.12)g/条,养殖3个月,收获时均重为(158.05±28.85)g/条,可见,鲤提高了稻田系统物质及能量利用率。植物的生长和产量与其环境条件有密切的关系[20-21],有研究表明,稻田养鱼对水稻的产量有一定的保持或促进作用[19],本研究中,试验组与对照组环境因子存在较大差异,试验组稻田开挖了沟渠及占地5%的鱼涵,其实际种植面积小于对照稻田面积的 95%。试验期间,两组稻田管理基本一致。试验结束时,养鲤稻田和单种稻稻田干谷产量分别为(452.73±28.15)kg和(462.92± 35.47)kg,二者无显著差异(P>0.05),说明尽管稻田养鲤减少了水稻种植面积,但可能由于其环境条件更有利于水稻生长,其水稻产量并未降低。
3.2 浮游植物群落结构特征
浮游植物作为生态系统的初级生产者,其丰度和生物量与水体初级生产力正相关[22]。本研究中,试验组浮游植物丰度和生物量均高于对照组,可为稻田系统其他生物提供更多饵料,造成二者差异的原因可能是鲤的活动及代谢使稻田水体环境更利于藻类的繁殖,从而使浮游植物生物量增加,这一结果与袁伟玲等[8]的研究结果一致。本研究中,试验组的蓝藻门密度及生物量显著低于对照组(P<0.05),绿藻门和隐藻门显著高于对照组(P<0.05),结果与宋庆洋等[9]对稻虾系统的研究结果相一致。有研究表明,浮游植物的生长条件受其所处环境的温度、pH、氮磷含量和有机质的影响[23],其群落结构也常常受到其他生物摄食等活动的影响[24],在水温一定的条件下,水体营养盐含量越高,蓝藻生长优势越大,同时,蓝藻可通过竞争作用抑制其他藻类生长[25-26]。试验组鲤的加入降低了水体营养盐含量,降低了蓝藻的优势度,说明稻田养鲤可改善浮游植物群落组成。此外,本研究中试验组含有金藻门的锥囊藻,而对照组未检测到。锥囊藻富含 β-胡萝卜素、白糖素和油滴,可作为生物饵料,多分布于贫营养淡水中,水体营养盐升高时消失[27],这反映了稻田养鲤改善了浮游植物群落组成,并降低了水体营养盐含量。
本研究用 Margalef、Simpson和 Shannon-Weiner 3个指数来描述浮游植物的多样性状况,Margalef丰富度指数是反映物种多寡的指标,Simpson指数直接反映物种的均匀程度,Shannon-Weiner指数则是概括了丰富度和均匀度的综合性指标[28]。本研究中,试验组水体鉴定出浮游植物7门18种,对照组5门24种,导致试验组Shannon-Weiner多样性指数和Margalef丰富度指数低于对照组。试验组物种数低于对照组的原因在于蓝藻门物种的减少(试验组蓝藻门藻类1种,对照组7种),这进一步反映了稻田养鲤对浮游植物群落组成的改善效应。
3.3 环境因子对浮游植物群落的影响
水体环境因子与浮游植物群落结构存在较为密切的关系[23]。本研究中,试验组与对照组环境因子和浮游植物群落结构存在较大差异,通过冗余分析(RDA)对二者的相关性分析表明,WTN、与蓝藻门丰度正相关,与绿藻门、金藻门、隐藻门、裸藻门丰度负相关,STP与绿藻门、金藻门、隐藻门、裸藻门丰度正相关。有研究表明,TN、TP、NO3--N、COD、TDS均是影响浮游植物种类丰度的重要环境因子,其中是浮游植物吸收利用的无机营养盐,COD、TDS则通过影响水体透明度、氧化还原电位以及水质等间接影响浮游植物的光合作用[29]。不同浮游植物各指标最适范围不同[30],这是本研究中同一环境因子对不同藻类的影响效应存在差异的原因。以蓝藻门为例,当水温高于22 ℃时,其营养盐浓度的最适范围高于其他浮游植物,此时营养盐浓度越高,其竞争优势越大[26]。贾海燕等[31]研究指出,电导率是影响丹江口水库浮游植物的重要环境因子,本研究中电导率对浮游生物的相关性低于其他因素,原因可能是试验组和对照组的电导率均较低,差异较小,影响效果不及其他因素。
4 结论
本研究中,试验组水体的化学需氧量、溶解性固体总量、电导率、硝酸盐和总氮浓度均低于对照组,氨氮、亚硝、磷酸盐浓度与对照组无差异,说明稻田养鲤降低了水体离子和氮营养盐含量,对水体环境具有一定的净化作用。此外,试验组水体浮游植物的密度和生物量均高于对照组,且水体绿藻密度升高,蓝藻密度降低,说明稻田养鲤改善了水体浮游植物群落组成。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
发明与创新·中学生(2022年3期)2022-03-14
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
农业与技术(2019年11期)2019-07-12
小品文选刊(2016年21期)2016-11-27
海洋世界(2016年2期)2016-03-11
科学24小时(2015年3期)2015-09-10
幼儿智力世界(2015年5期)2015-08-20
