
植物高光效研究进展
2020-04-22 09:38刘虎虎陈永忠
生物学杂志 2020年2期
肖 璞, 刘虎虎, 王 翀, 陈永忠, 田 云
(1. 湖南农业大学 生物科学技术学院, 长沙 410128; 2. 国家油茶工程技术研究中心, 长沙 410004)
光合作用是植物生长发育的基础,培育对光能具有高效利用能力的植物品种,对于提升植物的生产水平将起到重要的作用。高光效植物是具有高光能利用效率的植物。在高等植物中,主要的光合碳同化途径有3种:景天酸代谢途径(CAM)、C3途径和C4途径。在一定条件下,理论上C3植物的光能转化效率为4.6%,C4植物可达到6%,而实际上田间主要C3作物,例如稻麦品种的光能利用效率仅为1%左右[1-3]。因此,提高植物光能利用效率尚有巨大的潜力。
绿色植物在进行光合作用时,首先通过捕光天线系统捕获太阳能,在光系统复合体的反应中心将其转化为电能,电子再进一步在光合膜的各个蛋白复合体和电子传递体上进行传递,将能量储存在ATP和NAD(P)H中。最终,植物将光能以有机物的形式储存。所以对于植物高光效的研究首先在于光合作用过程中光能的高效吸收、电子的快速传递与能量转化,其次在于如何有效地减少光能的损失,再是提高碳同化效率等3个方面[4-5]。目前,研究人员分别围绕这3个方面对植物高光效的影响开展了大量研究,并取得了重要的成果。但是,整合这3个方面综述其对植物高光效影响的报道少见。因此,本文主要综述光合作用过程中3个方面对植物高光效的影响(图1),同时围绕当前研究存在的不足进行讨论,并对未来发展进行展望,以期为提高作物产量提供理论支撑。
1 光能的高效吸收、快速传递与转化
光合器官上光能的捕获是光合作用的起始,是高光效的基础。在植物叶绿体中,类囊体上的蛋白复合体通过捕获光能并驱使电子传递来完成光合作用的第一步。参与光合作用的4种主要蛋白复合体包括:光系统II(photosystem II,PSII)、细胞色素b6f(cytochromeb6f,Cytb6f)、光系统I (photosystem I,PSI)以及ATP合酶(ATP synthase,ATPase)[6]。一般状态下,叶绿素含量的下降会影响叶片光捕获能力,使电子传递效率下降,从而引起光反应中对光能吸收的下降。然而,郭晶晶等在研究叶绿素含量降低的大豆时发现,幼叶时期的大豆具有较强的光捕获能力;并且随着叶绿素含量降低,光合电子向光呼吸传递的比例增大[7]。该研究表明植物体内叶绿素含量降低会使得更多的光穿过顶层叶片,通过增加低层叶片的光合作用效率,有利于提高群体的光合作用;同时含量降低还有利于光合系统内氮素的分配,提高光合氮素利用效率。此前,Kirst等研究也表明氮素在蓝细菌光合作用系统中起到重要作用[8]。此外,利用更高效的光收集系统改造植物将是提高植物高光效的一条新途径。现有作物光合作用利用波长400~700 nm,还不及太阳光谱一半。而研究发现光合作用的有效光可扩大至750 nm,使可利用光子数量增加19%[9]。Ho等通过研究蓝细菌(ChlorogloeopsisfritschiiPCC 9212)中编码叶绿素f(Chl f )psbA4的功能,证实了Chl f具有扩大远红外光谱吸收的能力,这为提升植物光合效率潜能提供了新思路[10]。在叶绿素效率较低的区域引入吸收更好的光收集复合物,如蓝细菌藻胆体组分或工程化的荧光蛋白也是一种可能[11-12]。
光能吸收之后,光合电子将通过PSII、PSI等光合膜复合物及一系列的电子传递体进行传递。Pesaresi等通过在拟南芥中过表达质体蓝素或藻类的细胞色素c6等电子传递体,发现植物的生物量显著增加,这表明优化与改造光合电子传递方式和速率可以增加光能利用效率[13]。此外,电子传递链所产生的ATP是固定CO2和为细胞提供能量以对抗非生物胁迫所必需的。每分子NADPH产生的ATP量在很大程度上取决于PSI周围的环式电子流与线式电子流的比例。然而,与循环电子流有关的复合体(PGR5 / PGRL1和NDH途径)的分子细节和调节仍然缺乏了解[14]。因此,加强对其研究将有助于提高ATP的产量。另外ATP合成酶本身也是一个重要目标。尤其,Fo复合体c环中c亚基的数目决定了通过复合体迁移的每个H+所产生的ATP化学计量比[15]。由于不同植物ATP合成酶亚基组成数不同,通过重新设计ATP合成酶,可增加NADPH在电子传递中产生ATP的能力。尤其在持续照明的条件下,这是提高植物光合效率非常有效的策略之一。
2 减少光合作用中光能的损失
研究人员发现在植物体光合作用中,光抑制通过非光化学淬灭(Non-photochemical quenching,NPQ)损失的能量占到总固碳能量的7.5%~30%[16]。此外由于光呼吸的存在,导致C3作物的光合效率降低20%~50%[17]。因此,通过研究光抑制和光呼吸的机理来减少能量的损失,这对提高植物光能利用效率尤为重要。
光抑制的机理主要有两个方面。一方面主要是热耗散的能量流失,其中非光化学淬灭是热耗散的一种主要形式。尽管在强光下,NPQ可用于保护植物本身,但是,当强光转变成弱光时,则会由于NPQ相对地处于高位状态而造成能量流失。此前,研究表明促进NPQ所需的叶黄素循环,能加快NPQ中能量依赖的热耗散组分的形成速率,从而使得植物从耗散的产热状态快速恢复原状态。而且,在田间试验中,植株叶片面积和株高均增加到20%以上[18]。因此,通过设计表达调控开关与优化NPQ模块成为提高作物光能利用效率和产量的一条有效途径。另一方面是光合系统结构的破坏。研究表明光抑制主要是在光合膜上产生的大量氧自由基对PSⅡ进行损伤和破坏,具体表现为荧光量与电子传递活性均下降[19]。实际上,PSⅠ与PSⅡ均容易受到光抑制伤害。通过改善植物的冠层光能分配的方式,不仅可以减少上层光能分配过量和下层光能分配过少,还可以避免对光合结构的破坏。如中科院上海植物生理生态研究所薛红卫研究员团队发现,通过适当的改变水叶倾角可提高水稻光合效率[20]。此外,植物体本身为了应对环境中的各种胁迫,进化出一系列由蛋白因子组成的应答机制,如水杨酸受体NPR1在植物中对低温胁迫的应答[21]。同样,植物对强光胁迫也有应答机制,这使得光系统可以快速修复[22]。因此,通过进一步解析光合结构破坏和修复机制,将为提高植物高光效的研究提供有利帮助。
光呼吸是植物在进行光合作用时所产生的一个损耗能量的副反应。但其在损耗能量的同时,还可以处理光合作用过程中所产生的毒性物质。光呼吸只有在光照条件下才发生,其呼吸底物为乙醇酸。最近,研究人员运用合成生物技术在烟草中成功构建出新型的光呼吸途径。这些新型代谢途径比天然途径更有效,在田间试验中可使烟草的生物量增加超过25%,使光能利用率提高17%。在重复试验中,通过利用RNAi抑制叶绿体中的乙醇酸转运蛋白使得烟草的生物量提高了40% 以上。这项工作表明,田间条件下,在抑制乙醇酸转运蛋白的同时,将新型光呼吸途径替代天然途径,可以有效提高C3作物的产量[17]。由于光呼吸是植物所共有的代谢反应,这为改造其他C3作物(如稻麦等)提供了理论与技术支撑。
3 提高碳同化效率
核酮糖-1,5-二磷酸羧化酶/加氧酶(Ribulose-1,5-bisphosphate carboxylase/oxygenase,Rubisco)是碳同化过程中的关键酶,长期以来一直是研究人员关注的主要目标。C4植物具有比C3植物更高的碳同化效率,原因之一是C4植物所具有的CO2浓缩机理(CO2-concentrating mechanisms,CCMs)使得其鞘细胞内含有较高浓度CO2含量。C4植物由于具有CCM,使得其Rubisco的羧化活性提高,又抑制了Rubisco的加氧能力,从而降低光呼吸,使其保持较高的碳同化效率[23]。所以,提高低光效C3植物碳同化效率关键在于以下两点:
1)提高Rubisco的活性。研究人员通过基因工程手段希望获得具有高CO2亲和力或高催化活性的 Rubisco,然而效果并不明显,主要是由于其转换速率慢且对底物的亲和力差[24]。此前,Lin等[25]研究发现部分藻类生物中Rubisco具有高催化活性,并首次成功地将烟草中的Rubisco替换为蓝藻的Rubisco酶。随后实验成功地提升了Rubisco酶的表达量,然而经过改造的烟草只能在具有高浓度CO2含量的条件下生长[26]。Salesse-Smith等对玉米Rubisco进行研究时发现,通过共表达RAF1和Rubisco亚基可提高其含量,同时还提高了碳同化效率。在该研究中,单独过表达Rubisco的亚基(Rubisco-SS,LS)对其含量影响甚微,在与RAF1共同表达后,提高的Rubisco含量超过30%。这表明RAF1可能与植物 Rubisco的含量有着重要关联。此外,研究还发现UBI-LSSS-RAF1(过表达Rubisco-SS,LS和RAF1)转基因植物在光饱和条件下,其增加了15% 的CO2同化量[27]。该研究为创建高催化活性的Rubisco酶的研究提供了新的思路。
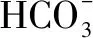
4 研究存在的不足
1)植物在生长发育过程中,受自身及外界环境因素的影响,不同时期的生理机能、空间位置具有差别,使得其光合特性也存在差别。目前对于植物高光效的研究主要集中在特定条件下对植物进行单一的研究,而对于植物不同生长发育时期光合特性动态的研究较少。2)缺少群体光合性能研究。植物群体光合性能受到多种因素的综合影响,研究群体光合性能更能体现实际生产中植物的光合能力。目前的研究对植物个体光合特性比较多,而群体与个体之间对光能利用能力存在较大的差异。3)缺少“源-库”关系的研究。植物高产的基础是高光合效率,但高光合效率不一定高产。植物产量形成的本质是源与库相互作用的过程。应进一步加强对植物产物形成机理的研究,通过调整产物成分的形成,减少有害物质的含量,从而提高植物产品的品质和综合效益。
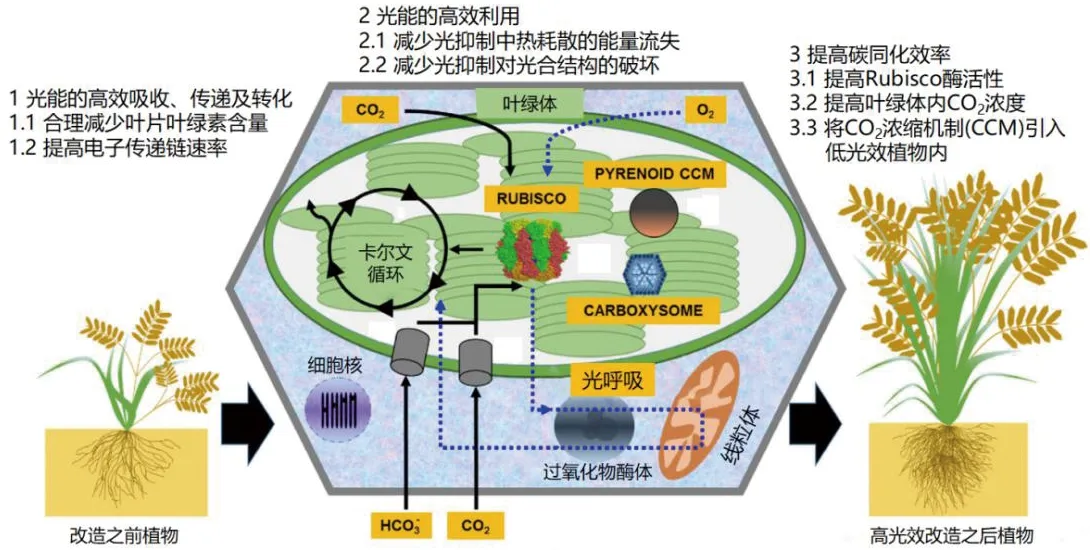
Rubisco:核酮糖-1,5-二磷酸羧化酶/加氧酶;Pyrenoid CCM:衣藻蛋白核的CO2浓缩机制;Carboxysome:叶绿体羧酶体
图1 提高植物光合效率的研究策略(根据文献[20]修改)
Figure 1 Research strategies on improving the photosynthesis in plants (modified according to reference[20])
5 展望
开展植物高光效研究在理论和实践上具有重要意义,一直以来受到科研人员的关注。我国在植物光合作用机理研究方面已取得重要成果。在此之上,围绕如何进一步提高植物高光效,提出3点展望:1)随着模式植物基因组研究的不断深入,一些关键的光合基因不断发现,鉴定这些基因的功能将为其他植物高光效研究提供更多有效的途径,从分子层面进行全基因组的关联分析将会成为未来研究的重点。2)随着合成生物技术的日渐发展,其对农业、生物制造、资源环境以及生物制造等很多领域具有深远的影响。通过合成生物技术来提高植物光能利用效率将是未来的关键途径之一[5]。3)近年来,科研人员对于植物根际微生物的研究取得了重要的成果,但对于具有丰富微生物多样性的叶际生态系统研究报道有限。叶际的范畴包括叶、茎、花、果等,是一个富含微生物的生态环境,每克新鲜叶片表面包含约几十万个细菌、真菌、藻类等微生物。这些叶际微生物对于现代植物产业生产以及对保护、修复生态环境有显著作用。研究这些叶际微生物并阐明相互关系及作用机理,将对提高植物产量具有重要的理论指导。未来,结合全基因组关联分析、合成生物技术以及叶际微生物研究提供的策略与技术,将为进一步提高植物高光效,解决粮食问题提供强大的推动力。
猜你喜欢
今日农业(2019年14期)2019-01-04
中国病理生理杂志(2018年6期)2018-01-22
中华老年口腔医学杂志(2016年3期)2017-01-15
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
原子与分子物理学报(2015年1期)2015-11-24
少儿科学周刊·儿童版(2015年10期)2015-11-07
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2012年1期)2012-10-26
中国医学科学院学报(2012年3期)2012-03-25
