
Relationship between the stoichiometric characteristics and leaf functional traits of Castanopsis hystrix in different ages*
2020-05-07 11:24ZhangZhongruiQiYeDingXiaogangYangHaiyanWeiDanZhangGengZhuHangyongChenLi
林业与环境科学 2020年2期
Zhang Zhongrui Qi Ye Ding Xiaogang Yang Haiyan Wei Dan Zhang Geng Zhu Hangyong Chen Li
(Guangdong Provincial Key Laboratory of Silviculture, Protection and Utilization / Guangdong Academy of Forestry, Guangzhou, Guangdong 510520, China)
Abstract Both the stoichiometric characteristics and leaf functional traits can reflect the adaptability of plants to changes in the external living environment. In particular, for varying aged forest plants, the study of stoichiometric characteristics and leaf functional traits can reflect the plant’s life history strategy and its resource investment and allocation methods. In this paper, the 1.5-year-old, 5-year-old, 15-year-old and 20-year-old Castanopsis hystrix were selected as the research objects to investigate the trends of stoichiometry and leaf functional traits, and their synergistic changes were verified. The results showed that with the increase of age, the stability of C. hystrix stoichiometry gradually increased, and the functional traits also tended to invest in security. However, there was no synergistic change between the two, which might be attributed to the different sensitivity of the C. hystrix leaf to the environment at the life history level of 1.5-year-old to 20-year-old.
Key words Castanopsis hystrix;different age;stoichiometry;leaf functional traits
Stoichiometry is a science that studies the balance of multiple chemical substances in biological systems and ecological processes in the context of ecology (Reed et al., 2012). In recent years, the Stoichiometry has been widely used in the study of nutrient characteristics of forest vegetation, and plays a unique role in identifying the nutrient limitations of regional forests and the relationship between nutrients and plant functional traits (Zeng et al., 2017). The study of the Stoichiometry measurement ratio of vegetation carbon(C), nitrogen(N), phosphorus (P) and the level of community nutrient supply can reveal the relationship between nutrient supply characteristics and vegetation development in various regions. In particular, N and P are two major elements and main limiting factors for forest growth in southern China (Sistla & Schimel, 2012). In particular, the ratio of nitrogen and phosphorus is a very important indicator of the absorption ability of N and P elements by plants. Therefore, it is necessary to study the dynamic characteristics of nutrient up-take indicators to reflect the supply of nutrients to forest growth in southern China (Yu et al., 2010).
Plant functional traits are the adaptive performance of plants to changes in the external living environment (Violle et al., 2007). These characteristics (such as morphology, physiology, etc.) reflect the resultant impacts of plants on the environment during growth and of the strategy of plant life history and its ecological indication function in the ecosystem (Wright et al., 2006). Because of a rapid growth cycle, fast regeneration (renewal), a large number of plant leaves, and the most sensitive to the change of habitat conditions, the plant leaf and their functional traits have become an important indicator of plant resource utilization, energy allocation, and balance of growth strategies (Ordoñez et al., 2009). The leaf functional traits at different age can reflect the strategy of plant life history and reflect the material investment and distribution mode of plant nutrients (Lavorel, 2013).
For species growing in a particular habitat, the leaves are senescent with age, and the resulting redistribution of resources is the main reason for the difference in leaf traits. During the aging process, the content of the main nutrient elements (C, N, P) of leaves decreased (Scalon et al., 2015; Valliere & Allen, 2016). In the process of plant evolution, the functional traits of each leaf are co-evolutionary, so there is a strong interaction between them (Wright & Sutton-Grier, 2012; Funk & Cornwell 2013), so the functional traits of other leaves may also change. Castanopsis hystrix is one of the main forest species in the subtropical regions of China, which is the excellent materials and has a wide range of uses. It can be renewed normally and naturally in the forest ecosystem, as well as widely plantation. Taking C. hystrix, a typical native tree species in South China as an example, this paper attempts to answer the following two questions by studying the relationship between the stoichiometric characteristics and leaf functional traits of C. hystrix stand at different age: 1) with the increase of age, the change trend of the stoichiometric characteristics of C. hystrix and the functional characteristics of leaves; 2) whether the stoichiometric characteristics can coordinate with leaf functional traits to reflect the response of C. hystrix to changes in the external environment at different ages.
Materials and methods
Research area
The study area was located in three forest foarms in Guangdong Province. The test material of 1.5-year-old and 20-year-old C. hystrix are located in Jinji Forest Farm in Guangdong Province with geographical location of (24°8′N, 113°0′E); the test material of 5-year-old C. hystrix is located in Longyandong Forest Farm in Guangdong Province, and its geographical location is 23°12′N, 113°24′E; the test material of 15-year-old C. hystrix is located in Jiulianshan Forest Farm in Guangdong Province, and its geographical location is 24°21′N, 114°30′E. The three areas all have a tropical marine climate of South Asia, the average elevation varied from 250 to 300 m for the afforestation sites; the slope is 10 to 20 degrees, the southeast slope dominated. The soils at three site are acidic mountain red soil. The time of this survey is November 2018.
Sampling and measurement
Five mean trees were selected from the 1.5-year-old, 5-year-old, 15-year-old and 20-year-old C. hystrix pure forests. From each tree one healthy leaf was collected from the upper, middle, and lower parts of the plant canopy with four directions from the east, south, west, and north respectively. A total of 12 leaf samples were collected from each plant per age group.
The collected leaves samples were brought back to the laboratory, washed with deionized water, and then (1) characterizing each leaf as stated in the below section (Characterizing each leaf sample); (2) crushed each sample freshly with the mortar; and (3) determining nutrient index based on conventional analysis methods; Nitrogen was measured by the Kjeldahl method; the determination of Phosphorus is based on the Mo-Sb Antispetrophotography method, and the Potassium is measured by the method of the Flame Atomic Absorption Spectrometry.
Leaves character calculation
Characterizing each leaf sample:
(a) fresh weight, dry weight, length, width, the ration of leave length and width, leaf area and specific leaf area of each leaf sample were quantified individullyas follows: flattening fresh leaves, washing the surface with Deionized Water and drying, measuring the fresh weight(mf) of a single leaves with an analytical balance, scanning the leaves image with a scanner for determining leaf length L, width D, aspect ratio L/D, area S of a single leaf with the Adobe Photoshop CC Software;
The calculation formula for the specific leaves area (SLA) is as follows:

(b) The relative water content and dry matter content of leaves drying are determined as follows. Fresh leaves were taken to weigh their fresh weight(mf) and were completely immersed in deionized water for 24 hours according to the number of treatment group. The saturated wet weight was measured, and the dry weight(m) was measured after drying (all sampled component samples were dried at 70 °C to constant weight for determining dry weight(m)) . The calculation formula for dry matter content and saturated moisture content of leaves is as follows:
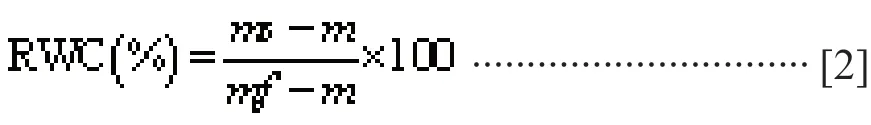
Data Processing
Excel 2019 was used to analyze the obtained data and produce the resulted graphs. This analysis also used the One-way ANOVA. An analysis of variance (ANOVA) was performed using version 21 of SPSS software (SPSS Inc., Chicago, IL, USA), followed by multiple-comparison tests (least-significant-difference test) to determine the leaf functional traits and the stoichiometric characteristics difference among different forest age groups. A significant level of 0.05 was used in all statistical analyses.
Results
Nutrients and stoichiometric characteristics of C. hystrix in different forest ages
Compared the nutrients in the forest tree leaves at different age, it was found that the contents of C, N, and P in the leaves all increased with the increase of age, and the contents of C, N, and P in the leaves of 20-year-old were significantly higher than those in other ages (fig. 1a, fig. 1b and fig. 1c). Different from this, only 1.5-year-o ld and 15-year-old have significantly higher C:N than 20-year-old, and no significant differences in C:P and N: P were found among age groups of forest tree leaves (fig. 1d, fig. 1e, fig. 1f).
With the increase of age, there is no clear pattern identified in the length-width (ratioRspect ratio) of C. hystrix (Fig. 2a). LDMC of 1.5-year-old was significantly smaller than other forest ages and there was a slightly increasing linear trend when the forest age increased from 1.5 years to 20 years. RWC and SLA of 1.5-year-old were significantly larger than other forest ages (Fig. 2b, Fig. 2c, Fig. 2d).
The stoichiometric characteristics and leaf functional traits of C. hystrix in different forest ages were analyzed using the Principal Component Analysis (Fig 3). The first two principal components accounted for 87.20% of the total variance, of which the first principal component accounted for 56.47% and the second principal component accounted for 30.72%. The first principal component could account for more than half of the variation in leaf traits. According to the ranking chart of each trait, it can be seen that the stoichiometric characteristics of leaves (i.e., N, P, C: N, and C: P) converged on the first ranking axis, and the leaf functional traits (i.e., SLA, RWC, and LDMC) converged on the second ranking axis. It showed that the first ranking axis mainly represents the variation of the stoichiometric characteristics of leaves, and the second ranking axis mainly represents the variation of the leaf functional traits. Moreover, in the overall variation of leaf functional traits of C. hystrix of different forest ages, the variation contribution of the stoichiometric characteristics of leaves is greater than that of the leaf functional traits.
Discussions
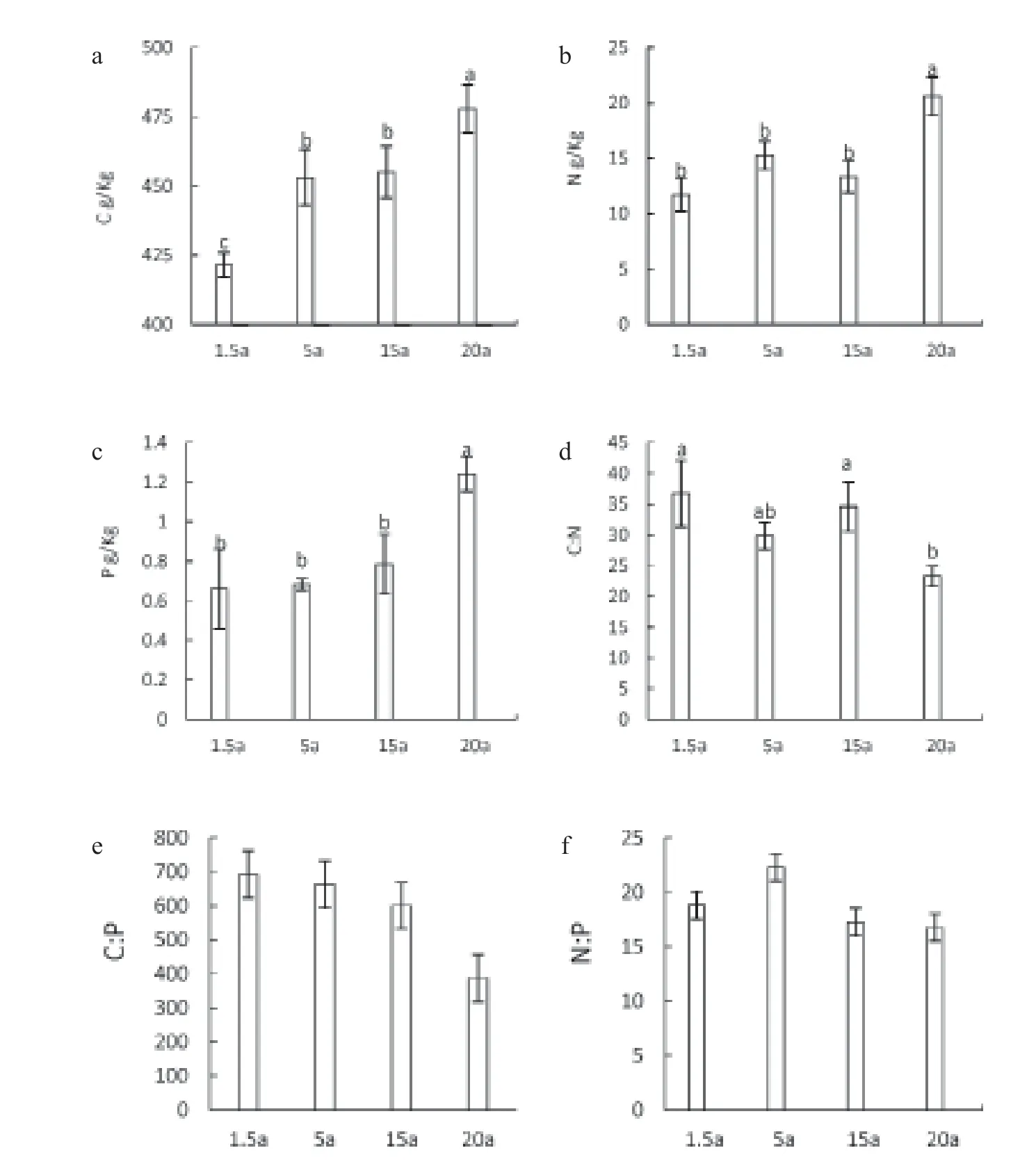
Fig. 1 Variation in of leaf C、N、P content and stoichiometric ratio of C. hystrix forest with age
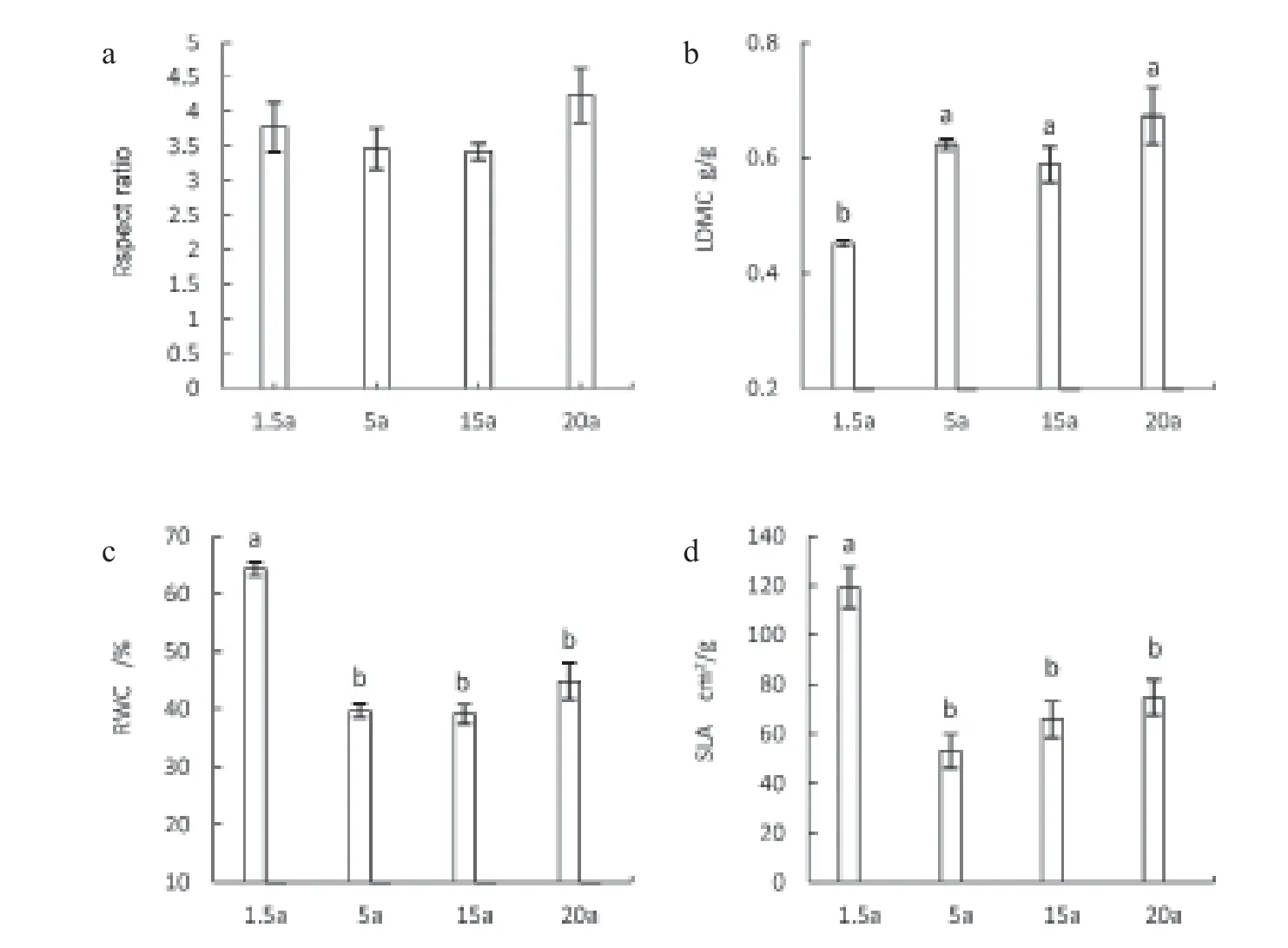
Fig. 2 Leaf functional traits of C. hystrix in different forest ages
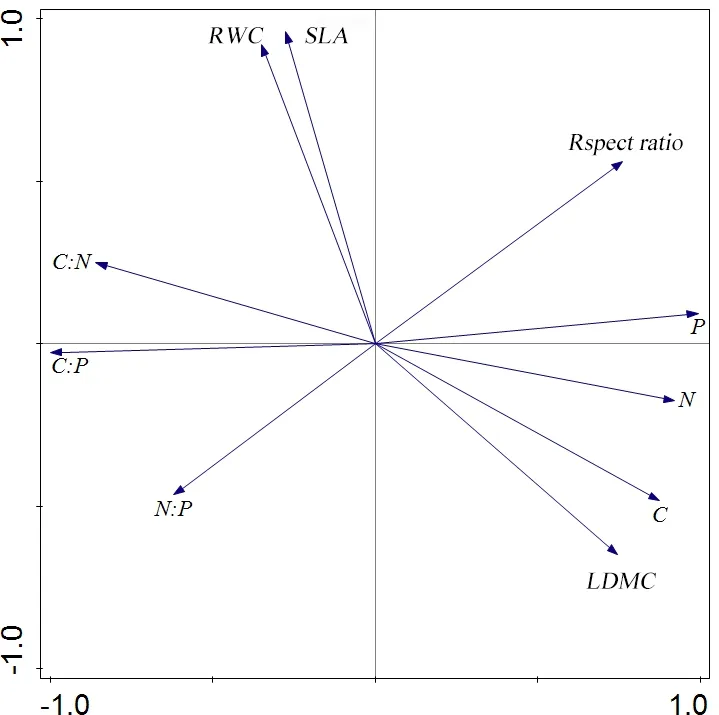
Fig. 3 Two dimensional sequence diagram of stoichiometric and functional properties of C. hystrix based on Principal Component Analysis (PCR)
C, N, and P play an important role in the growth, development, and behavior of organisms (Zeng et al., 2017). C is the most important element that makes up the dry matter in the plant; as a key element, N plays an important role in the biosynthesis of amino acids, proteins, nucleic acids, and other substances, as well as in improving photosynthesis of plants; P is a basic element of the tissues and is a component of nucleic acids and enzymes (See et al., 2015). In this study, from age of 1.5 to 20, the contents of C, N, and P elements in the leaves of C. hystrix tree increased. However, C: N showed a gradually decreasing trend. As a result of basic physiological and biochemical adjustments, C: N can reflect the response of the organism to the environment. The combination of leaves life and the optimal element ratio for growth results in higher stoichiometric stability of leaves C:N (Zhang et al., 2013). In this study, the C: N of 1.5-year-old and 15-year-old C. hystrix were significantly higher than those of 20-year-old, indicating when the C. hystrix forest is young and the leaves life was relatively short, the stability of C:N ratios were relatively weak (Su et al., 2000).
Plant functional traits are the adaptive performance of plants to changes in the external living environment (Violle et al., 2007). These characteristics reflect the environmental impact of plants during growth and their response to environmental change. Among them, SLA represents the light capture capacity of leaves dry mass per unit area of a plant (Adler et al., 2014). LDMC and RWC characterize the water content of plants under natural conditions, and the aspect ratio is the adaptability of plants to changes in the light environment (Brodribb et al., 2013). In this study, the LDMC of the 1.5-year-old young forest was significantly lower than other forest ages, and its RWC and SLA were significantly higher than other forest ages. It reflects the adaptation of C. hystrix plant in different forest ages to optimize the resource and nutrient environment by adjusting the basic structure of their leaves (Niinemets, 2014). With the increase of tree height, the increased water gravity potential energy leads to water transmission difficulties, in response to greater water pressure and lower RWC, the plant adapted the changes by improving LDMC, to ensure hydraulic safety. On the other hand, as the forest ages increases, the C. hystrix canopy is more at risk of being exposed to physical environments, and the leaves need to be more robust to protect against the risk, the crown layer reduced SLA tomake the leaf blades stronger against the risk. (Brodribb et al., 2013).
The change of plant leaf functional traits with leaf age can reflect the living strategy of plants, and also reflect the material investment of plant nutrients and distribution of plant leaves. Especially the leaves of the typical evergreen tree species in South China, because of their long life history, there are different resource investment, distribution patterns and traits characteristics of plants at different ages, growing environment and development stages (Medlyn et al., 2011). The growth of forest ages and the redistribution of resources resulting from changes in system structure are the main causes of leaf traits differences, which may also change due to the co-evolution of the various leaf traits during plant evolution (Huang et al., 2018). In this study, the stoichiometric characteristics and leaf functional traits of C. hystrix at different forest-age were ranked in a non-constrained manner, which was belonging to two sorting axes respectivelyhttps://cn.bing.com/dict/search?q=respectively&FORM=BDVSP6&mkt=zh-cn. It is shown that there is no coordinated variation between the characteristics of C. hystrix and the leaf functional traits between different forest ages. This may be due to the different environmental sensitivities of the two types of leaf traits. The stoichiometric characteristics of plant leaves reflect the adaptability of plants to soil nutrient effectiveness, especially when mature individuals are often able to effectively regulate plant nutrient utilization between old and new leaves, resulting in greater stability in the context of changes in the resource environment (Donovan et al., 2014).
