
利用CRISPR-Cas9技术对鲍曼不动杆菌噬菌体进行改造的实验方案*
2020-06-21 06:51郭雨薇
生物学通报 2020年8期
和 渊 郭雨薇
(中国人民大学附属中学 北京 100080)
STEM课程侧重于现实世界的问题解决,有科学探究或工程设计过程,培养学生运用所学知识、创造性解决问题的能力。本次课程针对噬菌体疗法杀菌谱范围窄的问题,学生应用CRISPR-Cas9设计开发了尾丝蛋白序列基因改造的噬菌体,从而改变了噬菌体与宿主结合能力,或可对临床治疗提供一定的思路和操作路径。在此过程中,学生综合运用了生物学、数学、计算机等学科知识解决问题,并与同伴合作、沟通,提问、论证等环节培养了学生创造性解决问题的能力,使之成为适应性的问题解决者。
1 问题的提出
随着抗生素的不断使用,细菌的耐药性不断增强。其中,耐药性鲍曼不动杆菌导致的流行性疾病引发了世界各地的广泛关注[1]。鲍曼不动杆菌(Acinetobacterbaumannii)可导致呼吸道感染、泌尿系统感染、烧伤后感染、败血症、伤口感染等症状[2]。近年来针对鲍曼不动杆菌的治疗难度逐渐增高,目前尚无合适的抗生素能有效抑制多重耐药性鲍曼不动杆菌的生长。在此背景下,寻找针对耐药菌的治疗方法已成为了当前研究的重中之重。近年来,一种有前景的治疗方法——噬菌体疗法逐渐发展,相比于抗生素,噬菌体疗法具有高效性和特异性等优势[3-4]。但由于噬菌体对宿主菌的识别和感染具有高度的特异性,使噬菌体杀菌谱过窄,难以广泛投入应用。
在STEM课程教学中,教师向学生介绍CRISPR-Cas9技术可进行基因编辑,学生提出“是否可利用CRISPR-Cas9技术改造噬菌体的特异性以解决鲍曼不动杆菌的抗药性”的问题,笔者认为这是一个非常值得探究的问题。CRISPR-Cas9技术是近年来出现的基因编辑新技术,而噬菌体展示技术也获得了2018年诺贝尔化学奖,因此,该问题的提出非常接近前沿的科学问题。通过探究性学习和基于项目式的学习方式,开展对此问题的可行性分析和实验方案设计,也非常符合STEM教学中的“科学、技术和工程”结合的精神,可培养学生的学科核心素养和解决真实问题的能力。
2 实验目标的确立
根据已提出的问题,确立本实验目的是通过对噬菌体基因组进行编辑,改变噬菌体用于特异识别宿主菌的受体结合蛋白,从而改变其宿主特异性,拓宽噬菌体的杀菌谱[5-6]。为合理设计针对多重耐药鲍曼不动杆菌具有杀伤能力的噬菌体,并初步评估其临床应用潜力,学生需要解决如何设计可特异性结合泛耐药性鲍曼不动杆菌的噬菌体尾丝结构、如何利用大肠杆菌的CRISPR-Cas9系统合成并纯化噬菌体、如何筛选合成的噬菌体、如何评估噬菌体疗法的治疗效果这4个关键问题,实验目标和内容如图1所示,实验目标的确定符合STEM教学科学探究的要求。
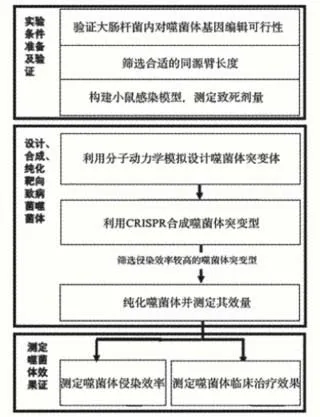
图1 实验目标
3 实验流程
本研究方案的设计旨在尝试应用CRISPRCas9系统实现对噬菌体基因组中编码尾丝蛋白序列的基因编辑,通过对鲍曼不动杆菌噬菌体改造和筛选,以获得针对泛耐药菌株的噬菌体制剂。CRISPR-Cas9是近年来迅速发展的一项对基因进行编辑的技术,学生在思考解决问题方案时,查阅了大量资料,确立了尝试采用这一手段进行研究。事实上,利用此技术在噬菌体上进行操作还鲜有报道,学生的尝试是一次开放性的探索,在理解的基础上运用多样化的学习策略深度加工知识信息,充分体现了STEM探索精神。实验流程如图2所示。
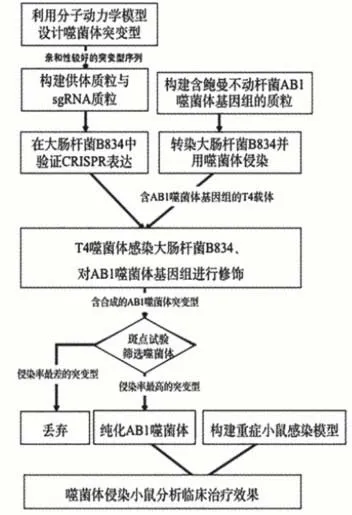
图2 实验内容流程图
3.1 针对泛耐药性鲍曼不动杆菌菌株的噬菌体设计 噬菌体的特异性决定于噬菌体尾丝蛋白底座与宿主菌表面脂多糖、蛋白质等分子的特异性结合。噬菌体的尾丝具有可伸缩性,当尾丝蛋白底座与宿主或环境信号结合时,尾丝会通过收缩穿透细菌外膜并注射遗传物质至细菌细胞质中。不同菌株的鲍曼不动杆菌其噬菌体对应的尾丝蛋白结构略有所不同,因此,在设计噬菌体时,首先需要对基于相对广谱的鲍曼不动杆菌噬菌体进行改造。使用分子动力学模拟(MD simulation)可分析不同突变型噬菌体的结构及不同噬菌体和泛耐药鲍曼不动杆菌的亲和性,通过模型计算给出的预测结果,可帮助下一步实验筛选出特异性结合泛耐药菌效果最好的噬菌体结构。
3.2 通过CRISPR-Cas9系统实现对噬菌体基因组的合成 除设计噬菌体外,本研究还需要能实现对噬菌体的合成和纯化,且合成机制应当便捷高效。利用同源重组修复机制,可实现对基因组上编码衣壳蛋白底座部分序列的任意替换。本研究需要注意的问题是,传统的Cas9蛋白以NGG为PAM位点,但是鲍曼不动杆菌噬菌体基因组中CG碱基对含量相对较低,可能存在脱靶效应,因此,构建供体质粒前,需在已经验证CRISPR表达的大肠杆菌中,转入具有不同长度同源臂的供体质粒,以筛选最合适的同源臂长度(图3)。图3绘制了通过遗传工程对噬菌体进行改造的具体模式图。
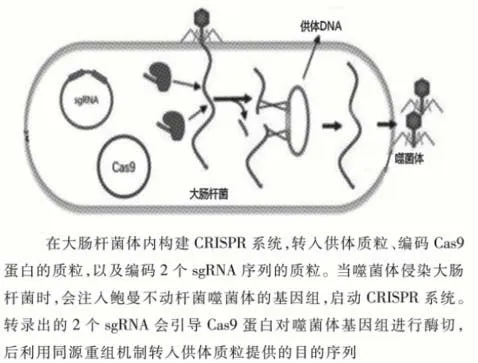
图3 利用CRISPR系统在大肠杆菌内实现对噬菌体的合成
3.3 对合成噬菌体的筛选和纯化 在合成噬菌体后,需筛选侵染效率最高的噬菌体用于临床治疗。此过程可采用斑点实验,测定特异性噬菌体的有无,并根据筛选结果对噬菌体进行进一步纯化。
3.4 对噬菌体侵染效果、临床治疗效果的评估为评估合成的噬菌体的治疗效果,首先需要测定噬菌体侵染泛耐药鲍曼不动杆菌的效率。这一过程可通过斑点实验,对于噬菌体的效价(pfu/mL)进行测定,以评估噬菌体特异于鲍曼不动杆菌的侵染效果。此外,还需要应用数学建模的知识,验证基因编辑的效率和准确度,以及噬菌体在小鼠感染模型中的临床治疗效果。
在整个探究流程中,运用了数学、遗传工程学、噬菌体侵染病理学、计算机科学等一系列的跨学科知识。
4 实验步骤的设计
4.1 验证CRISPR系统在大肠杆菌中能否进行多位点切割 向大肠杆菌B834感受态中转入编码dCas9的质粒(Addgene no.48645),表达sgRNA的质粒(Addgene NO.42229)。sgRNA应互补于编码鲍曼不动杆菌AB1噬菌体尾丝蛋白序列上下游的一段20 bp左右的DNA序列。dCas9质粒上具有绿色荧光蛋白用于检验转化是否成功,而表达sgRNA质粒上具有氨苄抗性用于检验转染是否成功。挑选转化成功的大肠杆菌菌落培养于1∶1 LB/M9CA培养基上并放置于37℃。向处于指数增长期的鲍曼不动杆菌Aba46(每毫升含有约2×108个细胞),取300μL接种含有鲍曼不动杆菌AB1噬菌体基因组的T4噬菌体(m.o.i为1时)100μL,并于37℃培养15 min(因鲍曼不动杆菌AB1噬菌体潜伏期为10 min),离心后取上清获得编辑后的噬菌体。将分别稀释为10、102、103体积的感染上清液,与未经过基因编辑的同浓度感染的上清液,涂在鲍曼不动杆菌Aba46培养皿上并过夜培养。统计感染效率并提取编辑后噬菌体基因组进行测序,计算公式如下:

4.2 设计同源臂长度梯度寻找最优长度 在已经验证CRISPR表达的大肠杆菌B834中,转入不同供体DNA质粒pET28b(含有长度为500 bp,700 bp,900 bp,1.1 kb,1.3 kb同源序列,转入序列为鲍曼不动杆菌AB1噬菌体基因组本身序列,即基因编辑前、后噬菌体基因组序列不应出现改变),挑选转化成功的大肠杆菌菌落,培养于1∶1 LB/M9CA培养基上并放置于37℃。向处于指数增长期的大肠杆菌B834(每毫升含有约2×108个细胞),取300μL接种含有鲍曼不动杆菌AB1噬菌体基因组的T4噬菌体(m.o.i为1时)100μL,并于37℃培养15 min,离心后取上清液。用分别稀释为10、102、103体积的上清液,与未经过基因编辑的同浓度鲍曼不动杆菌AB1噬菌体上清液,以及实验3.1中编辑后的同浓度鲍曼不动杆菌AB1噬菌体上清液,感染泛耐药鲍曼不动杆菌Aba33并过夜培养。统计感染效率,挑选噬菌斑进行基因测序验证基因编辑准确度。
4.3 利用分子动力学模拟对鲍曼不动杆菌AB1噬菌体突变型进行设计 在分子动力模拟中,以鲍曼不动杆菌AB1噬菌体的底盘结构信息,与泛耐药鲍曼不动杆菌Aba46菌株表面特异识别蛋白作为位置信息输入,改变底盘结构上亲水氨基酸种类,并利用分子动力学,模拟计算出2个结构间亲和性,计算方式如下:
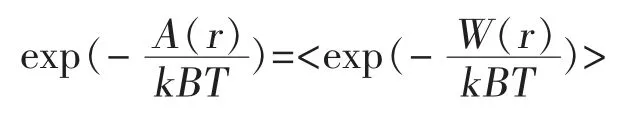
这一公式可计算结构的自由能,并以此侧面表现2个结构间亲和性。选择亲和性超出原本结构中最好的10种突变结构构建质粒。
4.4 利用CRISPR在大肠杆菌中替换噬菌体基因组中编码尾丝蛋白序列并进行斑点实验 在已经验证CRISPR表达的大肠杆菌B834中,转入不同供体DNA质粒pET28b(上含有同源序列以及编码目的噬菌体编码尾丝蛋白的DNA序列),挑选转化成功的大肠杆菌菌落,培养于1∶1 LB/M9CA培养基上并放置于37℃。其余方法同3.3,之后统计感染效率,挑选11种噬菌体侵染获得的噬菌斑。
4.5 鲍曼不动杆菌噬菌体的纯化 挑选实验3.5中形状规则、边缘光滑的单个噬菌斑,接种于2 mL鲍曼不动杆菌Aba46宿主菌液,于37℃以160 r/min振荡培养4~6 h,4°C,l0 000 r/min离心l0 min。最后得到的上清液,用0.22μm微孔滤膜过滤除菌。重复上述过程3~5次,即得到纯化的噬菌体。
4.6 构建重症小鼠感染模型 将泛耐药鲍曼不动杆菌Aba46加入少量肉汤培养基中,在36℃培养箱培养18 h,再将此菌液涂在斜面营养琼脂培养基上,于36℃培养16 h后,用灭菌处理过的生理盐水冲洗,取冲洗所得液体稀释建立不同梯度,利用比浊法确定浓度同时计数其中活菌数量。取0.5 mL不同稀释浓度的泛耐药鲍曼不动杆菌Aba46对5组小鼠进行腹腔注射,观察小鼠生存情况,记录小鼠全部死亡的最小剂量为绝对致死量。以此浓度腹腔注射小鼠建立重症感染模型。
4.7 编辑后噬菌体临床治疗效果分析 将模型小鼠分为5组,每组10只,每组内又随机选取5只作为对照组。向实验组小鼠中注射相同效价的噬菌体,而对于对照组小鼠注入等浓度等体积的生理盐水。不同实验组小鼠所注射的鲍曼不动杆菌浓度不一,浓度最低为注入等浓度生理盐水,最高为注入绝对致死量浓度鲍曼不动杆菌,注入体积均为0.5 mL。
5 实验结果的预期
5.1 验证CRISPR系统在大肠杆菌中能否进行多位点切割 如果CRISPR系统能实现在大肠杆菌中,对鲍曼不动杆菌AB1噬菌体的多位点切割,经过基因编辑的鲍曼不动杆菌AB1噬菌体感染效率,应显著低于对照组感染效率,因为若基因编辑成功,噬菌体基因组会在大肠杆菌内被切割降解。此处应注意,对照组噬菌体(即原AB1噬菌体)感染效率也会相对较低,因AB1噬菌体并不适用于泛耐药鲍曼不动杆菌菌株Aba46,故还需提取编辑后的噬菌体基因组进行测序。
5.2 设计同源臂长度梯度寻找最优长度 含有不同同源臂长度的供体质粒的大肠杆菌编辑后的噬菌体感染效率不同,选取基因编辑无误差突变,感染效率最高的噬菌体对应同源臂长度作为最优长度。
5.3 利用CRISPR在大肠杆菌中替换噬菌体基因组中编码尾丝蛋白序列并进行斑点实验 经过基因编辑的鲍曼不动杆菌AB1噬菌体对于泛耐药鲍曼不动杆菌感染效率相对较高,说明对噬菌体尾丝的修饰扩大了原本不适用于泛耐药鲍曼不动杆菌的AB1噬菌体的杀菌谱。
5.4 编辑后噬菌体临床治疗效果分析 向实验组小鼠中注射相同效价的噬菌体、对照组不注射,由于噬菌体能侵染鲍曼不动杆菌,所以各个实验组中的小鼠均不会患病死亡,而对照组会有不同程度的患病现象(其中最高浓度组小鼠死亡)。
本实验是学生在STEM课程中的一个有益尝试。随着抗生素发展的减缓和细菌耐药性的增加,噬菌体疗法成为一种新的疗法,学生通过提出问题、查阅资料、可行性分析、目标确定、内容展示和步骤详解,利用CRISPR-Cas9技术基因编辑解决噬菌体疗法不够广谱的问题。在这一过程中,学生通过高效地实现对噬菌体的设计、改造、生产和检验过程,自己分析问题、讨论问题、与同伴沟通、提问和论证,在解决问题的过程中应用了生物学、数学、计算机等学科的知识,发展了学生创造性解决问题的能力,有效实现了STEM教学中科学探究的教学目标,为培养学生的学科思维、学科能力奠定基础。
猜你喜欢
军事文摘(2022年16期)2022-08-24
传染病信息(2022年2期)2022-07-15
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年4期)2021-11-12
今日农业(2021年11期)2021-08-13
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
World Journal of Radiology(2020年4期)2020-07-17
科学24小时(2020年4期)2020-05-14
幼儿园(2016年4期)2016-03-14
