
天山林区土壤总氮矿化过程对季节性冻融的响应
2020-07-31 08:20常顺利张毓涛张云云
生态学报 2020年12期
陈 磊,常顺利,*,张毓涛,张云云
1 新疆大学资源与环境科学学院绿洲生态教育部重点实验室, 乌鲁木齐 830046 2 新疆林科院森林生态研究所, 乌鲁木齐 830063
土壤冻融通常是因季节或昼夜气温变化而出现土壤冻结和融化的现象[1],其实质为土壤水的相态变化过程[2],冻融以多种方式影响着高纬度和高海拔区域的土壤氮素转化过程[3]。氮矿化是森林土壤矿物氮的主要来源[4],其多受土壤pH值[5]、微生物量氮(Microbial biomass nitrogen, MBN)[6]、群落结构[5]和土壤酶活性[6]等因子的影响。国内外研究学者认为冻融过程会显著影响净氮矿化、反硝化和硝化等作用[3,7],另外水、热等环境因子及微生物量氮对氮矿化过程有重要的影响[8]。
天山林区是干旱区山地的主要组成部分,对全球气候变化十分敏感[20]。林区所处纬度与海拔较高,冻融时期长达4—6个月,在秋冬与冬春交替期间温度变化较大[20]。由于冬季雪被覆盖,野外调查与取样较为困难,该区冬季土壤氮矿化动态及影响因素的研究仍相对缺乏。因此,本文在天山北坡林区设置不同类型植被样地,根据测定数据分析土壤氮矿化动态及与水、热环境因子的关系,拟回答以下科学问题:(1)冻融期间天山林区不同群落类型土壤总氮矿化速率、累积量及季节贡献率;(2)各氮素之间的相关关系以及温度、含水率和MBN等因素对土壤总氮矿化速率的影响。以便深入了解冻融期天山林区土壤氮循环过程及冻融期土壤氮素转化和迁移,以期为天山林区可持续经营提供理论指导,同时为该地区氮循环提供参考。
1 材料与方法
1.1 研究区概况
以中国森林生态系统研究网络(China Forest Ecology Research Net, CFERN)天山森林生态系统定位研究站(43°24′48″—43°26′18″N, 87°27′29″—87°28′48″E)为依托进行样地布设及采样,定位站海拔为1908 m。属温带大陆性气候,最热月平均气温为 10—16℃,最冷月平均气温-11— -9℃,年降水量350—600 mm,雨季集中在6—8月,年蒸发量980—1150 mm,年均相对湿度65%,平均无霜期89 d,≥10℃积温为1170.5℃,最大积雪深度为65 cm。土壤冻结期为11月初至次年4月底,持续6个月。天山云杉温带针叶林为该地植被类型的优势种,郁闭度约为0.4—0.8,林下土壤为山地灰褐色森林土[21]。
1.2 研究方法
1.2.1 试验设计与采样方法
于2017年9月植物生长季末期在定位站附近的乔木林地(woodland)、灌丛(bushland)和草地(grassland)3个群落结构中,分别随机布设3个20 m20 m的样方(图1)。在每个样地内布设40根不锈钢管(钢管直径为11 cm、长度为20 cm、管壁厚度为2 mm),平行每隔约7 m设4排,每排10根土柱。因钢管较为坚固、管壁较薄,取用厚木板置于其上截面部分,垂直敲击钢管于土壤中(为防止部分根系被切断后植物死亡,需要为每个土柱补充200 mL水)。为冻结期时方便取样,在每个钢管的外管壁套塑料薄膜,距上截面1.5 cm处对称开孔(孔径为0.5 mm)。其中每个样地20根用于测定矿质氮含量和净氮矿化速率,另外20根用于测定总氮矿化速率,共布设360根。
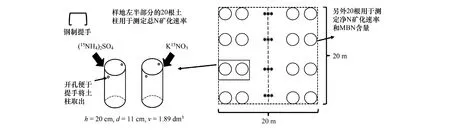
图1 样地布设图Fig.1 Sampling plot layout
为测定总氨化和总硝化速率,在每个样地中随机取出2个土柱,分别对土柱进行K15NO3和(15NH4)2SO4同位素溶液标记(选取30%丰度,上海化工院),置于装有冰袋的保温箱中。另外,在每个样地中随机选取1个土柱(20个土柱按相邻位置被分为10对),另一个土柱留待两周的原位反应后采取,用于测定净氮矿化速率和MBN含量。
为测定表层土壤(-5 cm)的温度和含水率连续变化数据,在每种群落类型选取1个样地布设便携性温度计(MicroLite 5032P-RH, Fourtec-Fourier Technologies, Israel)和FD探针(ECH2O- 5, Decagon Devices, Pullman, USA),设置每30 min采样一次,每日分别产生48个温、湿度数据,便于形成连续数据(图2)。在样地中心位置挖取1个5 cm浅坑(坑底需平整),将温度计和FD探针放入后,复原土壤环境。每两个月将温湿度探测器的数据导入笔记本电脑中,并为探测器更换电池。日均降水、日均气温等数据由定位站提供。分别在秋季冻融期(AFP, Autumn Freeze-thaw Period, 2017.11.01—2017.11.17),冻结期(CFP, Constantly Frozen Period, 2017.11.17—2018.3.12)和次年春季冻融期(SFP, Spring Freeze-thaw Period, 2018.3.12—2018.4.15)进行采样(每月的中旬或下旬各1次),共采10次。
1.2.2 实验室分析方法
利用15N稀释技术确定总氨化和总硝化速率[22],将土柱置于恒温恒湿培养箱中培养(温度设为样地前三日平均土壤温度)。标记后的18小时(t1)和42小时后(t2),分别取出样品置于冰箱冷冻室中(-20℃)存放,以便送往检测机构(ICAS英格尔检测认证集团, 上海)测得不同培养时间的15N丰度。另外,将采集的土壤样品经过自然风干后过0.149 mm筛(100目)。土壤硝态N含量使用酚二磺酸比色法测定,铵态氮利用靛酚蓝比色法测定,MBN用氯仿熏蒸-提取(FE)法测定。另外土壤含水率用称重法测定(105℃, 12 h)。
1.2.3 氮矿化速率及氮转化累积量计算方法
在查阅文献的基础上,结合本实验研究现状,采用Kirkham和Bartholomew给定的公式确定总氮矿化速率[23]:
(1)
(2)

净氮矿化速率采用以下公式[24]计算:
(3)
(4)
Rm=Ramm+Rnit
(5)
Ramm、Rnit、Rm分别代表土壤氨化速率、硝化速率和矿化速率(mg N kg-1d-1)。
N0=∑N1
(6)
(7)
N0为整个时期(2017.11.01—2018.04.15)的氮素累积量(kg N hm2a-1),N1为单次氮素转化量(kg N hm2a-1),N2为AFP、CFP和SFP不同时期的氮素转化量占比(%),如公式(7)所示。
1.2.4 数据处理与分析方法
采用Excel 2016和SPSS 19.0软件对数据进行统计分析,并利用Origin 8.5和Prism 7软件作图。通过双因素方差分析(Two-Way ANOVA)来确定冻融时期及3种群落类型对总氮矿化、净氮矿化和MBN的影响。用Pearson相关分析法分析土壤总氮矿化、净氮矿化和微生物量氮之间相关性。用线性回归模型来分析土壤温度和含水率对总氮矿化、净氮矿化和MBN的影响。
2 结果分析
2.1 冻融期土壤温度、含水率的变化
为研究冻融期间土壤总氮矿化过程,本文对土壤表层日均含水率与日均温度进行了监测(图2)。2017年秋、冬季,天山林区土壤初次冻结于11月20日夜间形成,于白天土壤温度高于0℃时结束。从2017年11月下旬至2018年3月下旬,≥20 cm深度的土壤处于冻结状态。另外从2017年11月上旬至2018年4月末,土壤含水率增加了10.2%(图2)。次年3月中旬至4月末为春季冻融期,这期间土壤频繁发生冻结-融化现象。
2.2 冻融期总氮矿化速率及MBN含量变化
在整个时期间的总氨化速率(图2)特征表现为随着土壤温度下降而持续下降,与总硝化速率和MBN含量相比,它以相反方向变化,即伴随最小的氨化速率,土壤MBN含量处于上升位置。各个冻融时期,3种群落类型土壤总氨化速率较为接近。表层土壤总氨化速率于11月中旬达到峰值(6.75 mg N kg-1d-1),随土壤含水率和土壤温度的下降而至低点(1.3 mg N kg-1d-1),冻结期土壤总氨化速率变化不明显。春季冻融期间,随着土壤温度和含水率的上升,总氨化速率缓慢上升(图2)。
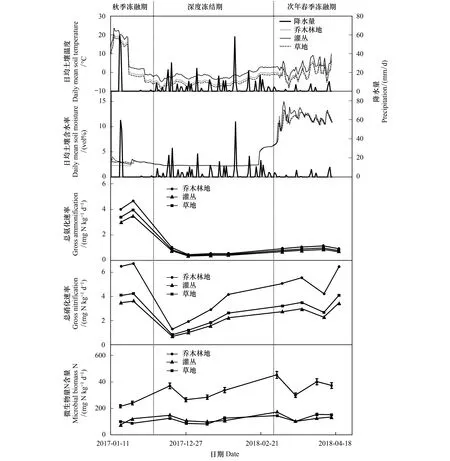
图2 土壤总氮矿化速率、微生物量氮含量、温度及含水率变化Fig.2 Dynamic changes in gross nitrification rate, gross ammonification rate, MBN content, temperature and moistureAFP:秋季冻融期Autumn freeze-thaw period;CFP:冻结期Constantly frozen period;SFP:次年春季冻融期Spring freeze-thaw period
与总氨化速率呈持续下降趋势相比,总硝化速率在整个时期呈较为强烈的震荡变化,数值在1.3—6.75 mg N kg-1d-1之间(12月中旬和4月中旬,以乔木林地土壤为例)。总硝化速率在秋季冻融过程中先增加至3.5—4.7 mg N kg-1d-1,随着土壤温度和含水率下降呈减小的趋势(图2)。从群落类型来看,在秋季冻融期林地土壤总硝化速率相较另外两种类型土壤较高,并且上升速率提升更快。
土壤MBN含量随着表层土壤冻结后下降,但12月中旬的一场降水致使土壤含水率上升,MBN含量随之而增加。春季冻融期间温度相较于冻结期有所回升,MBN含量随之增加,但增幅不明显(图2)。整个冻融期间,林地土壤MBN含量始终大于灌丛和草地。
2.3 冻融期对总氮矿化累积量、季节贡献率的影响
为了获得氮转化累积量,利用计算可以说明秋季和春季冻融期对整个冻融时期总氮矿化的重要性(公式 6, 公式7)。整个期间乔木林地、灌丛和草地表层土壤总氨化累积量分别为163.93 kg N hm2a-1、88.27 kg N hm2a-1和103.41 kg N hm2a-1,相同样地类型在不同冻融时期有显著差异(P<0.05);而总硝化累积量为55.43 kg N hm2a-1、47.12 kg N hm2a-1和41.58 kg N hm2a-1,对总硝化作用来说,相同样地类型在不同冻融时期也有显著差异(P<0.05),并且乔木林地的总氨化和总硝化累积量最多。对于净矿化作用来说,在相同时期不同群落类型土壤净氨化和净硝化累积量没有显著性差异(表1)。
秋季冻融期(17 d)的总氨化累积量均值占整个时期的29.5%,秋季冻融期总硝化量均值占比为57.1%;春季冻融期(32 d)总氨化占比为36%,总硝化占比为20.3%。总氨化在秋、春冻融时期与全部冻融时期占比为66%,总硝化转化量占比为77.4%(表1)。

表1 总氮矿化累积量、净氮矿化累积量在3种土地类型的不同时期所占比例
整体上冻融时期及3种群落类型对总氮矿化速率、净氮矿化速率和MBN均有极显著(P<0.01)的影响。群落结构对以上所有指标均有极显著的影响(P<0.01),冻融期对除净硝化速率以外所有指标都有极显著(P<0.01)影响(表2)。
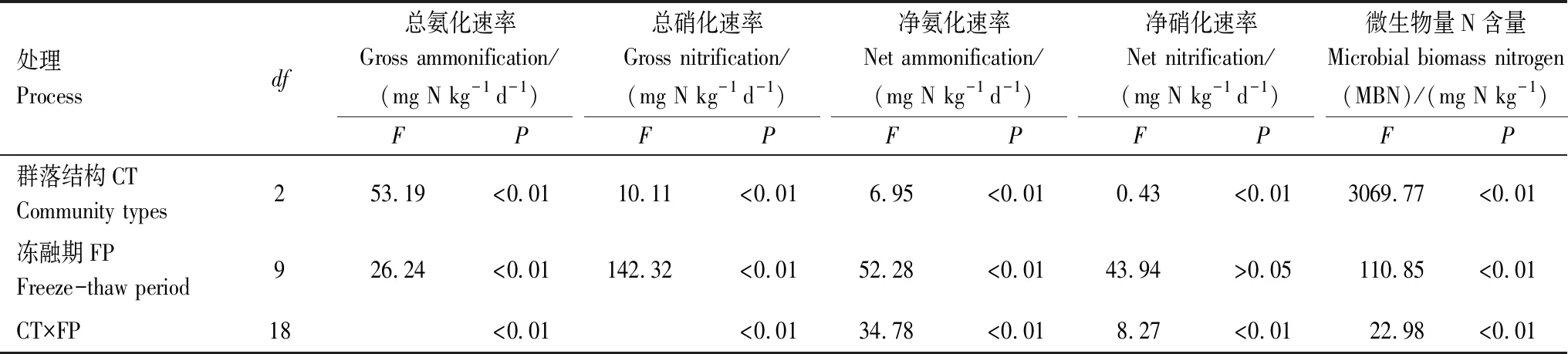
表2 冻融期及群落类型对总氮矿化速率、净氮矿化速率和微生物量氮含量的影响
总氨化速率与总硝化速率、净硝化速率、MBN呈极显著的正相关关系(P<0.01),说明总氨化速率与总硝化速率之间有一定的相互影响,同时总氨化速率还受MBN的驱动作用;总硝化速率与净硝化速率、净氨化速率、净硝化速率呈极显著的正相关关系(P<0.01),其中净氨化速率和净硝化速率相关性较小,同时总硝化速率也受MBN的影响,总硝化速率和MBN、净氨化速率和MBN速率为负相关关系,其中净氨化速率和MBN速率之间呈极显著负相关关系(P<0.01, 表3)。
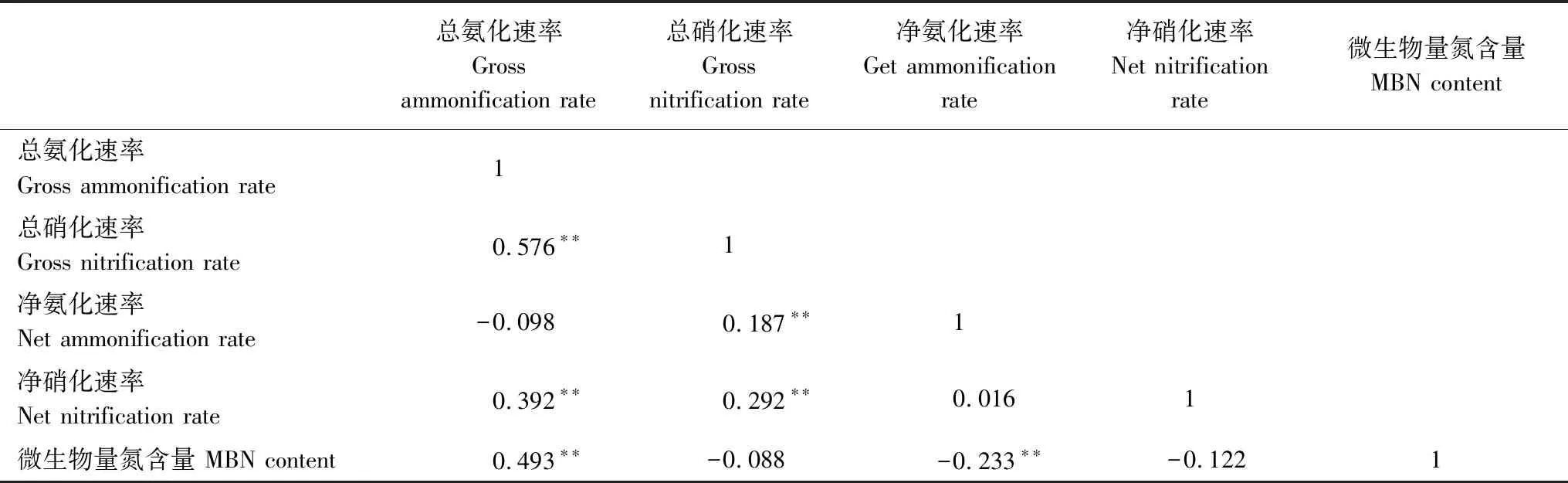
表3 总氮矿化速率、净氮矿化速率和微生物量氮含量之间相关性
2.4 土壤温度及含水率对氮素动态的影响
在整个冻融时期,总氨化速率在整体上呈现出与土壤温度相关关系一致的规律,但是3种群落类型的相关程度有所差异。随着土壤温度的升高,总氨化速率(林地和灌丛)显著增加(P<0.05);但MBN含量却与土壤温度表现相反,当土壤温度升高时MBN(林地和草地)含量显著降低(P<0.05, 图3)。

图3 土壤温度对总氮矿化速率、净氮矿化速率及微生物氮含量的影响Fig.3 Effects of daily mean soil temperature on gross N mineralization rate, net N mineralization rate and microbial biomass N content
当土壤含水率上升时,净氨化速率(灌丛)、净硝化速率(灌丛)和净矿化速率(灌丛)显著下降(P<0.05),而总硝化速率和土壤含水率之间没有显著相关关系(图4)。
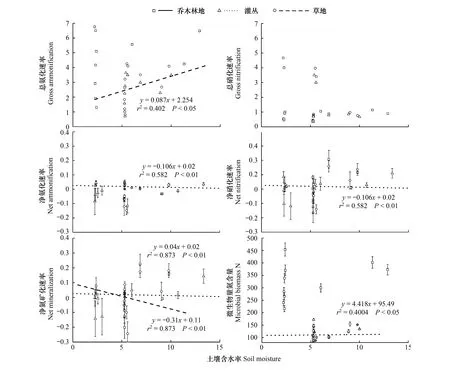
图4 土壤温度对总氮矿化速率、净氮矿化速率及微生物氮含量的影响Fig.4 Effects of daily mean soil moisture on gross N mineralization rate, net N mineralization rate and microbial biomass N content
3 讨论
3.1 冻融对氮矿化速率动态的影响
3.1.1 冻融对总氮矿化速率的影响
总氮矿速率能够反应土壤氮素周转情况和供氮能力,对认识植物获得可利用无机氮及生态系统氮素流失的途径起到重要作用[22]。冻融时期氮矿化速率动态变化得知,冻融过程中土壤温度对总氨化速率和总硝化速率影响显著(P<0.05, 图3),冻融期的总氮矿化速率比冻结期更高(图2)。这一观点得到了几项在草原、农业上研究的支持[25-26],这些研究认为在冻融时期极端低温改变了土壤结构,影响土壤团粒大小和稳定性,继而影响氮素的循环和流失,使得冻融条件下总氨化和总硝化速率较高,从而促进了土壤的氮矿化过程[25-26](图2)。对比冻结期,秋季、春季冻融期的总氨化量和总硝化量占比为一半以上(表1),与内蒙古草原土壤总氮转换量的研究结果相符[2],这一时期土壤温度和含水率有所上升,微生物也更加活跃,土壤中氮素矿化过程增长更为明显[27]。
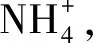
3.1.2 冻融条件下总氮矿化速率与净氮矿化速率的关系
本研究通过对比总氮矿化和净氮矿化累积量后发现,总氮矿化累积量为83.29 kg N hm2a-1,净氮矿化累积量为3.53 kg N hm2a-1,前者是后者的23.59倍(表1)。这一结果与前人研究相近,表明总矿化累积量和净矿化累积量的比值约为23倍左右[9]。当森林土壤小型无机氮库的产生和固定速率较快时,仅考虑净矿化速率可能会低估氮的累积量[30]。对内华达山脉森林土壤的研究中,净氨化年累积量为16—37 kg N hm2a-1,而总氨化量约为此的7倍[31]。
3.1.3 冻融条件下不同群落类型对总氮矿化速率的影响
3.2 冻融条件下土壤温度、含水率及MBN对总氮矿化的影响
3.2.1 冻融条件下土壤温度、含水率对总氮矿化的影响
3.2.2 冻融条件下土壤MBN含量对总氮矿化速率的影响
土壤温度和含水率对土壤氮矿化速率的影响主要是通过改变微生物的生理特征而进行[37]。冻融过程使微生物群落数量及多样性显著降低,当土壤微生物死亡后,残余微生物利用其营养物质的恢复力直接决定土壤氮矿化周转能力[37]。随着土壤MBN含量增加,总氨化速率显著增加(P<0.01, 表3)。相关研究表明,冻融对土壤微生物区系、数量和酶活性上均有影响[38]。当土壤温度下降时,土壤中冰晶体的膨胀和扩充使得微生物细胞膜和细胞壁有机械损伤,从而导致细胞形态发生变化[38];另外当土壤水冻结时,水溶液发生浓缩,微生物细胞渗透压失去平衡,细胞膜流动性减少,导致细胞受到伤害[38]。
在本研究中,群落类型和冻融过程对MBN有极显著的影响(P<0.01, 表2)。这与奥地利温带山毛榉林土壤观察的现象相似,微生物生物量的周转和演替是土壤中可利用氮的主要来源[39],冻融期土壤中MBN的累积可能会改善不同生态系统的氮利用效率(表2, 表3)。因为3种群落类型土壤的结构和孔隙度不同,当冻融事件发生时土壤孔隙中储存的水分和氧气均会有所变化,当氧气扩散到达一定限度时,微生物的有氧活动减少,氮矿化速率受到影响[40]。因此土壤温度、含水率和MBN是控制氮循环和微生物活性的主要因素。土壤氮素可利用性及其对气候变化的响应是预测气候变化背景下陆地生态系统碳、氮收支的关键之处[40]。由于全新世时期天山林区的湿度值一直在增加[41],春、秋季冻融循环过程也受到影响,总氮矿化过程对冻融循环响应的量化工作能够在一定程度上反映陆地生态系统的氮循环过程。
4 结论
综上所述,在天山林区,冻融期内土壤以总氨化过程为主;秋、春冻融期总氮矿化累积量占全年的一半以上,并且冻融期的总氮矿化速率相比冻结期更高;对不同的群落类型而言,冻融过程对总氨化速率和总硝化速率的影响差异明显。未来应进一步探究冻融条件下土壤氮矿化与碳矿化的协同效应及其氮素转化的关键微生物过程与机制,进而了解天山云杉森林中各类氮转化过程之间的关联性。
猜你喜欢
河南畜牧兽医(2020年11期)2020-01-11
水土保持研究(2019年6期)2019-10-19
中国粮油学报(2018年12期)2018-03-19
中国公路(2017年10期)2017-07-21
中国水土保持科学(2017年3期)2017-07-05
中国计划生育学杂志(2017年3期)2017-06-01
浙江大学学报(工学版)(2016年2期)2016-06-05
中国房地产业(2016年24期)2016-02-16
农村农业农民·B版(2015年9期)2015-10-16
养殖与饲料(2015年3期)2015-01-03
