
竞争强度变化对针阔混交林红松和水曲柳径向生长的影响
2020-07-31 08:21宣海憧郭梦昭高露双范春雨
生态学报 2020年12期
宣海憧,郭梦昭,高露双,范春雨
北京林业大学林学院,北京林业大学森林资源和环境管理国家林业和草原局重点开放性实验室,国家林业和草原局森林经营工程技术研究中心,北京 100083
采伐活动作为主要的人为干扰因素,是调整森林结构和功能的重要手段[1- 3]一直受到国内外众多研究学者的关注[4- 5]。大量研究表明,保留木的生长量随着采伐强度的增加呈现上升趋势[6- 8],Deal和Tappeiner[9]对阿拉斯加西南部的北美铁杉林和Man等[10]对北方针叶林的研究结果进一步证明了采伐对不同径级保留木生长量均具有促进作用。但也有研究指出,采伐后的1—2年内保留木生长量并没有出现明显的增加趋势,甚至随着采伐强度增加,部分保留木生长量出现降低的现象[11-12]。保留木生长受到物种特性,采伐强度[13]等多种因素的作用。保留木生长与采伐后林内其所处竞争环境的改变程度密切相关,而不同树种对竞争环境的响应滞后性也将改变保留木生长特征[14],这就导致采伐活动对保留木生长的影响具有一定的复杂性[15]。
因此,有必要基于物种特性和间伐强度检验时间尺度上竞争强度变化对保留木生长的影响,该结果对于当前森林经营与管理有着重要意义[16- 17]。本研究基于吉林蛟河针阔混交林内2012年进行不同间伐强度处理的3块固定样地,以顶级树种红松和水曲柳为研究对象,试图回答以下问题:(1) 不同间伐强度下红松和水曲柳保留木竞争强度变化特征(2) 不同竞争强度变化下保留木生长是否存在差异。
1 研究区概况和研究方法
1.1 研究区概况
2011年7月,在蛟河实验区管理局大坡林场,按照CTFS(Center for Tropical Forest Science,CTFS)标准在地形和林分状况大体一致的近熟林内设置3块面积为1 hm2(100 m×100 m)的固定样地,分别为2号样地(43°57.9625′N,127°43.5727′E,海拔443 m)、3号样地(43°57.9087′N, 27°43.4510′E,海拔430 m)和4号样地(43°57.8784′N,127°43.5560′E海拔447 m),并对样地内DBH≥10 cm全部个体进行调查,调查项目包括:号牌编码、物种名称、胸径、树高、枝下高、冠幅(东西长、南北长),并记录生长状况(枯立、枯倒、断头等)等信息。2012 年12 月以胸高断面积为指标确定间伐强度,按照间密留匀,留优去劣的原则,对3块样地实施不同强度的抚育措施,以达到调整林分结构和促进保留木生长的目的。间伐强度分别为: 2号样地15%(轻度间伐)、3号样地30%(中度间伐)和4号样地50%(重度间伐)。该区域气候为温带大陆性季风气候,年均温在3.7 ℃以下,7月最热平均气温为 21.7 ℃,1月最冷平均气温为-18.6 ℃,年均降水量在700 mm左右,地带性土壤为暗棕色土壤,森林群落类型为典型的针阔混交林,主要乔木树种包括红松(Pinuskoraiensis)、千金榆(Carpinuscordata)、胡桃楸(Juglansmandshurica)、水曲柳(Fraxinusmandshurica)、紫椴(Tiliaamurensis)、色木槭(A.mono)、蒙古栎(Quercusmongolica)、春榆(Ulmusjaponica)等,主要灌木包括暴马丁香(Syringareticulata)、瘤枝卫矛(Euonymuspauciflorus)和东北鼠李(Rhmamnusdavurica)等。红松和水曲柳是针阔混交林的顶级树种。 红松属于浅根性树种,对土壤水分要求较严,对土壤的排水和通气状况反应敏感,不耐湿,不耐干旱,不耐盐碱。水曲柳喜光、耐寒,喜肥沃湿润土壤。鉴于两树种对生长环境的敏感度不同,因此,本研究选取典型针叶树种红松和阔叶树种水曲柳为研究对象。
1.2 研究方法
1.2.1 样芯采集与生长量数据获取
2013 年,2015 年和2017 年分别对间伐样地进行复测,复测项目包括:号牌编码、树种名称、胸径和生长状况,以确定实际采伐强度。经复测后计算得出样地实际采伐强度分别为17.24%、34.74%和51.87%(表1)。2017 年8 月在2号,3号和4号样地内采集DBH≥10 cm的红松和水曲柳的保留木胸径处宽度样芯(内径 5.15 mm)。将样芯带回实验室进行处理后,利用LINTAB 5 年轮宽度测定仪器获取红松和水曲柳的连年径向生长量(0.01 mm精度),通过COFECHA软件对数据的可靠性进行验证,交叉定年核定后,结合3次复测的胸径数据,重建各样地内红松和水曲柳历史胸径数据。
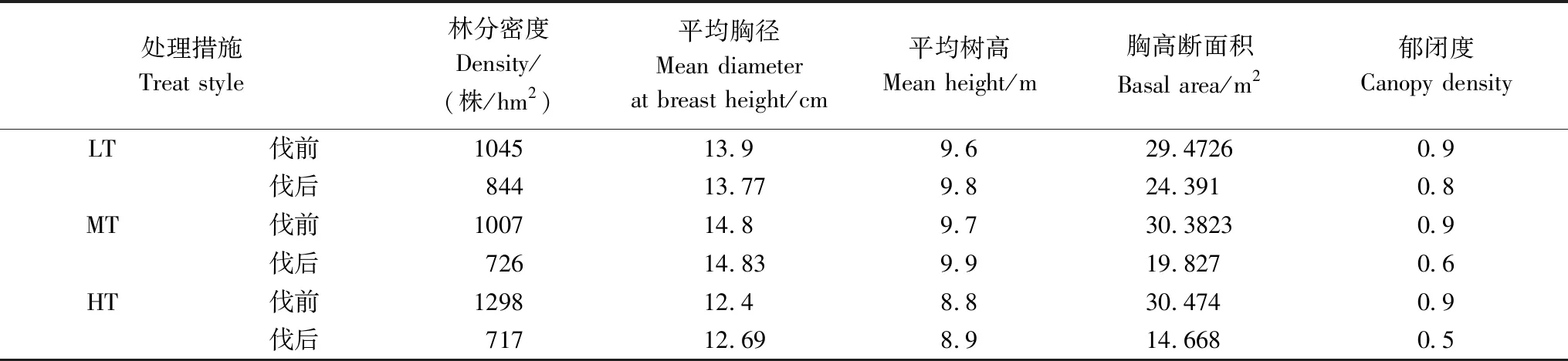
表1 采伐前后样地林分特征
1.2.2 竞争指标选取与量化
根据 Hegyi单木竞争指数模型公式计算出2012年(间伐当年)和2013年(伐后次年)每个竞争木对对象木的竞争指数,将2013年和2012年竞争指数相减即得该树种的竞争强度变化程度。本研究设置了10 m的缓冲区域排除边缘效应,10 m的缓冲区内未选取对象木,仅选取竞争木,从而可以保证距离对象木最近的相邻木均在样地内。具体计算公式为:
(1)
aba=Hegyi伐后-Hegyi伐前
(2)
式中,Dj代表相邻树胸径(cm),Di代表目标树胸径(cm),dij代表相邻树与目标树距离(m)。aba代表目标树间伐前后竞争指数的差值,Hegyi伐后为目标树伐后次年的竞争指数,Hegyi伐前为目标树间伐当年的竞争指数。
将树木竞争指数变化aba在5%以内的树木定义为竞争强度未发生变化,竞争指数变化大于5%的树木定义为竞争强度变大或变小,将aba为正数的目标树定义为竞争强度增强的树木,aba为负数的目标树定义为竞争强度减弱的树木。将竞争强度变化的保留木株数与各样地内该物种株树总数的比值定义为不同物种竞争强度发生变化的比例。通过配对T检验在0.05水平检验不同竞争强度变化下保留木径向生长是否存在差异,以上数据分析过程由R 3.6.1完成。
2 结果与分析
2.1 不同间伐强度下红松和水曲柳竞争强度变化特征
不同间伐强度下红松和水曲柳竞争强度变化有所不同(表2)。各间伐样地内水曲柳保留木竞争强度发生变化的单木所占比例基本一致,均达到65%以上,且有50%的水曲柳保留木的竞争强度均出现减弱的趋势。而红松保留木竞争强度发生变化的单木比例随着间伐强度增加而有所下降,但发生竞争减弱的保留木所占比例与间伐强度正相关。在轻度间伐样地内77%的红松保留木竞争强度均发生了变化,其中47%红松保留木竞争强度减弱,23%的红松保留木甚至出现竞争强度增加,在中度间伐样地内有64%红松保留木竞争强度发生变化,55%的红松保留木竞争强度减弱,在重度间伐样地内64%红松保留木竞争强度发生变化,63%为竞争强度减弱。
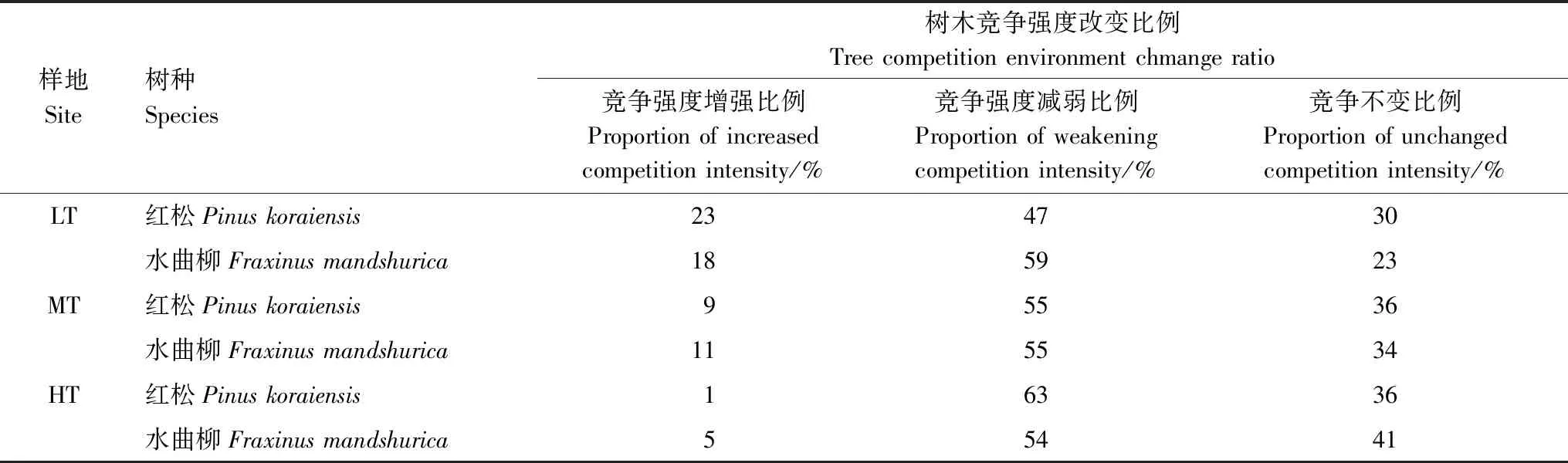
表2 不同间伐强度下林木竞争强度变化特征
2.2 不同竞争环境的红松和水曲柳保留木径向生长差异性分析
不同竞争环境下红松保留木径向生长变化趋势有所不同(图1)。轻度和重度间伐样地内竞争减弱的红松保留木径向生长在间伐后均呈上升趋势,而中度间伐样地内竞争减弱的红松保留木和各样地竞争不变的红松保留木径向生长则先出现下降(伐后2年内),而后出现上升。不同间伐样地内不同竞争环境红松保留木径向生长增加幅度也存在显著差异(图2)。竞争减弱的红松保留木径向生长在2015年(伐后第3年)随着间伐强度升高生长量增加,而竞争不变的红松保留木径向生长则未随着间伐强度增加而呈现任何规律性的变化。在轻度间伐样地内竞争不变的红松保留木径向生长在2013和2014年间(伐后2年内)径向生长下降幅度为0.2 mm,而竞争减弱的红松保留木径向生长增加幅度为0.35 mm,在2015年(伐后第3年)由伐前竞争不变的红松保留木径向生长显著高于竞争减弱的红松保留木(P<0.05)转变为二者不存在显著差异(P=0.92)。而在中度和重度间伐样地内竞争减弱的红松保留木径向生长年增量在2015年分别达到0.69 mm和0.79 mm,使得竞争减弱的红松保留木径向生长显著高于竞争不变的红松保留木(P<0.05)。
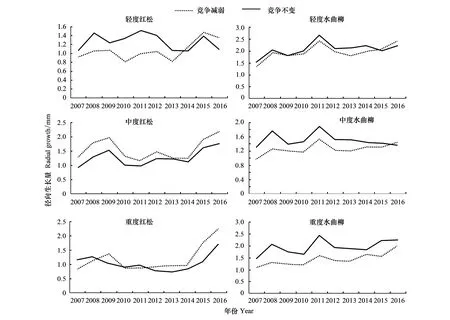
图1 不同竞争强度下红松和水曲柳径向生长量变化趋势Fig.1 Radial growth tendency of Pinus koraiensis and Fraxinus mandshurica under different competitive intensity
不同竞争环境下水曲柳保留木径向生长趋势基本一致(图1),均在2013年和2014年(间伐后2年内)出现生长抑制,而在2015年(伐后第3年)得到促进,但径向生长增加幅度有所不同,且竞争减弱的水曲柳保留木径向生长年增量与间伐强度密切相关(图2)。自2014年(伐后第2年),重度间伐样地内竞争减弱的水曲柳径向生长增加幅度最大,其次为轻度间伐样地,而在中度间伐样地内增加幅度最低。在轻度间伐样地内竞争减弱的水曲柳保留木径向生长在伐后第5年显著高于竞争不变的水曲柳(P<0.05)。在中度和重度间伐样地内,竞争减弱的水曲柳保留木径向生长由间伐前显著低于竞争不变的水曲柳保留木(P<0.05)转变为在伐后第5年与竞争不变的水曲柳保留木径向生长不存在显著差异(P=0.89)。竞争不变的水曲柳保留木径向生长的年增量与间伐强度没有明显的相关关系。

图2 不同间伐强度下红松和水曲柳保留木径向生长年增量变化Fig.2 Increment of Pinus koraiensis and Fraxinus mandshurica residual trees under different thinning density
3 讨论
不同间伐强度下树木竞争环境变化具有明显差异[19]。本研究三个样地林分水平采伐强度分别为17.24%、34.74%和51.87%,红松保留木竞争强度发生变化的单木比例随着间伐强度增加而有所下降,但发生竞争减弱的保留木所占比例与间伐强度正相关,而各样地内水曲柳竞争环境发生变化的单木所占比例基本一致。在物种丰富的林分中物种水平的间伐强度的影响可能会突显[20],本研究各样地内红松和水曲柳的物种采伐强度与林分水平采伐强度不同,其中红松的采伐强度分别为14%、11%和18%,而水曲柳的采伐强度为16%、17%和19%。因此,红松保留木竞争环境变化对林分水平间伐强度更敏感,而水曲柳保留木竞争环境可能受到物种水平间伐强度的影响。
采伐干扰下树木生长与竞争环境的变化密切相关[20],前人研究指出保留木径向生长量与采伐强度间呈正相关关系[7],随着采伐强度的升高保留木直径的生长也随之增加[8,21]。本文研究发现仅竞争减弱的红松和水曲柳保留木径向生长与间伐强度密切相关。竞争减弱的红松保留木径向生长在2015年(伐后第3年)呈现随间伐强度升高生长量增加的趋势,而水曲柳保留木随着间伐强度增加呈现先下降而后增加的趋势。在林分中伐后竞争强度下降的树种竞争能力增强进而生长得到促进[22- 23]。采伐活动可能增加森林内部的温度和空气流动能力,改变林内的光照和温度条件,显著增加部分树木进行光合作用的强度和时间,进而加速植物的蒸腾作用和土壤水分的蒸发,导致部分树木在伐后的1—2年内出现生长下降的现象[24- 25]。采伐后一般能够提高林内降雨量,但由于光照条件增强,林内空气湿度有降低趋势,对土壤温湿度及土壤理化性质产生显著影响[26]。红松和水曲柳对土壤水分条件的适应差异可能是导致间伐后红松和水曲柳径向生长变化存在不同的原因,针叶树和阔叶树对采伐强度的响应差异也有一定的影响[11,27]。竞争减弱的红松和水曲柳保留木的径向生长均在2015年(伐后第3年)呈现快速增加趋势则证实树木生长对采伐活动的响应可能存在一定的时间延迟[14]
综上,不同间伐处理下物种竞争强度变化明显不同,而不同竞争环境下保留木径向生长对间伐强度的响应也具有一定差异性,其响应关系受到间伐强度和物种特性的共同影响, 51.87%以内的间伐强度将有利于65%的竞争减弱红松保留木径向生长,而水曲柳保留木径向生长对间伐强度的响应更复杂,且可能对物种间伐强度更敏感,因此,利用间伐活动调整针阔混交林林分结构时应考虑间伐处理后竞争强度变化的单木所占比例,以及物种水平间伐强度对保留木生长的影响。
猜你喜欢
格言·校园版(2022年17期)2022-07-06
小哥白尼(野生动物)(2021年9期)2022-01-17
河北果树(2021年4期)2021-12-02
中南林业科技大学学报(2021年4期)2021-04-22
林业科技情报(2021年1期)2021-04-14
中南林业科技大学学报(2021年3期)2021-03-23
——以杉木抚育间伐为例
绿色科技(2020年21期)2021-01-08
新农民(2020年5期)2020-12-10
森林与环境学报(2020年4期)2020-07-27
农民致富之友(2019年11期)2019-05-23
