
澳系睡莲花粉离体萌发及低温保存研究
2020-09-02 06:54唐毓玮龙凌云黄秋伟苏群池昭锦卢家仕毛立彦*
热带作物学报 2020年7期
关键词:睡莲
唐毓玮 龙凌云 黄秋伟 苏群 池昭锦 卢家仕 毛立彦 *

摘 要:为研究适宜睡莲花粉离体萌发的培养基和低温保存的方法,以睡莲品种‘默笙和3份原种睡莲的花粉为试验材料,采用单因素、正交试验设计,筛选出最适宜的培养基,观测睡莲花粉在不同温度下保存的萌发率。结果表明,4种澳系睡莲花粉的最适培养基的各组分浓度不同,花粉的生活力存在差异,澳洲原生浅色睡莲与澳洲原生深色睡莲的花粉生活力较高,分别为43.90%和45.63%;‘默笙睡莲的花粉生活力次之,为31.84%;‘白巨睡莲的花粉生活力较低,为9.73%;4种澳系睡莲花粉均不耐贮藏,不同温度下保存72 h后萌发率均为0。本研究结果可为睡莲杂交育种的亲本选择、花粉的低温保存研究提供参考。
关键词:睡莲;花粉生活力;离体培养;低温保存中图分类号:S31 文献标识码:A
Pollen Germination in vitro and Cryopreservation Reserch of Anecphya Waterlily
TANG Yuwei1, LONG Lingyun1, HUANG Qiuwei1, SU Qun2, CHI Zhaojin1, LU Jiashi2, MAO Liyan1*
1. Guangxi Subtropical Crops Research Institute, Nanning, Guangxi 530001, China; 2. Flowers Research Institute, Guangxi Academy of Agricultural Sciences, Nanning, Guangxi 530007, China
Abstract: The pollens ofNymphaea‘Moshen and three orginalNymphaea accessions were used as the test material to study the optimal medium components for the pollen germinationin vitroofNymphaea and the method for prolonging pollen viability at low temperature. The single factor and orthogonal design were used to select the most suitable medium, and observe the germination rate of the pollens preserved at different temperatures. The optimum medium concentration for the pollens of the four accessions was different. The pollen viability varied among accessions. The pollen viability ofN. immutabilis subsp. Immutabilis blue formandN. immutabilis subsp. immutabilis purple form was 43.90% and 45.63% respectively, and that ofN.‘Moshen andN.‘Gigantea Albertde Lestangwas 31.84% and 9.73%, relatively. The pollen ofNymphaea was not tolerant to storage, and lost the viability after 72 hours of storage at different temperatures. The results of the study could provide references for parents selection and pollen cryopreservation in hybrid breeding ofNymphaea.
Keywords: Nymphaea; pollen viability;in vitro culture; cryopreservation
DOI: 10.3969/j.issn.1000-2561.2020.07.013
睡蓮是睡莲科(Nymphaeaceae)睡莲属(Ny- mphaea)植物的总称,属于多年生草本植物[1],花色丰富,花态多样,群体花期长,极具观赏价值,在水生园林景观中应用广泛。睡莲属植物主要以生态学为基础进行分类,可分为Anecphya、Brachyceras、Hydrocallis、Lotos、Nymphaea(orCastakua)5个亚属[2-5]。Anecphya亚属睡莲俗称为澳系睡莲,因原种主要分布于澳大利亚而得名,澳系睡莲适宜广西地区种植,群体花期长达8~9个月。澳系睡莲花态独特,花瓣形似荷花,与其他亚属的睡莲形态有明显差异,是重要的杂交育种材料[6]。
花粉生活力的强弱是影响杂交育种成败的重要因素[7],授粉前对植株的花粉生活力进行评价,选择花粉生活力较高的亲本可以提高育种效率。花粉离体萌发法评价花粉生活力是目前较为准确和直观的方法,在杜鹃、山茶、月季、海棠等植物的育种研究中均有应用[8-11]。关于睡莲花粉的研究较少,杨梦飞等[12]与毛立彦等[13]的研究均表明,睡莲花粉离体培养4 h后,萌发率趋于稳定;张慧会等[14]研究证明,30 ℃是睡莲花粉离体萌发的最佳温度。不同亞属的睡莲广泛分布在暖温带至热带地区[15],单朵花期约4~5 d,杂交育种经常受地域、花期等因素制约。延长睡莲花粉的生活力能有效解决花期不遇、异地授粉等问题,而低温保存是延长花粉生活力的主要手段[16],因此低温保存研究对于睡莲的杂交育种有重要意义。目前,关于睡莲花粉低温保存方面的研究还未见报道,本研究以澳系睡莲为试验材料,进行花粉离体萌发试验,筛选较佳的花粉离体萌发培养基,研究适宜的花粉保存温度,为杂交育种的亲本选择、种质资源的保存提供参考依据[17]。
1 材料与方法
1.1材料
试验地点位于广西壮族自治区南宁市兴宁区亚热带作物研究所睡莲种质资源圃(108.38° E,22.87°N),属于亚热带季风气候,年平均气温21.6 ℃,年平均降水量约1300 mm。试验地为睡莲种质资源圃,水体无浮萍、藻类等污染,水位平稳,水深约40 cm。
以4种澳系睡莲为试验材料,在花蕾时期做套袋处理,避免昆虫携带花粉干扰试验。随机选取长势良好、无病虫害的植株,在花药散粉时,采摘睡莲花朵带回实验室内,将成熟花药中的花粉均匀抖落在干燥培养皿中混匀(4种睡莲花粉分别置于4个培养皿中),再将花粉分装到若干支2 mL离心管中密封保存,用于花粉培养基筛选与花粉保存试验。
1.2方法
1.2.1 花粉离体萌发法 取出保存花粉的2 mL离心管,使用移液枪向离心管加入700 μL培养液,置于30 ℃培养室,500 lx的光照条件下自然培养,每隔1 h观察一次,直至花粉萌发率稳定为止。将离心管轻轻摇匀,用移液枪吸取50 μL花粉液滴在载玻片上,使用Leica DM 2500显微镜进行镜检,使用Image-Pro Plus 6.0软件统计视野内的花粉总数、萌发花粉数,每个玻片随机选取5个不重复的视野,每个视野花粉数量不少于40粒,以花粉管长度大于花粉粒直径视为萌发。培养基用水为双蒸馏水,试验重复3次。
花粉萌发率=(视野内萌发花粉数/视野内花粉总数)×100%
1.2.2 培养基筛选 以‘默笙睡莲的新鲜花粉为材料进行单因素试验设计,探讨蔗糖、MgSO4、H3BO3、KH2PO4、CaCl2·2H2O对花粉萌发的影响,其意义在于缩小正交试验因素水平的筛选范围。参照张瑞等[18]的方法,以BK培养基[7](100 g/L蔗糖、100 mg/L H3BO3、300 mg/L CaCl2·2H2O、200 mg/L MgSO4、100 mg/L KH2PO4)作为基本培养基,在探讨每个影响因素时,设置不同浓度梯度,其余因素均以BK培养基为假设最适值。
蔗糖设6个浓度梯度:50、100、150、200、250、300 g/L;MgSO4设6个浓度梯度:100、200、300、400、500、600 mg/L;H3BO3设6个浓度梯度:50、75、100、125、150、175 mg/L;KH2PO4设6个浓度梯度:100、200、300、400、500、600 mg/L;CaCl2·2H2O设6个浓度梯度:100、200、300、400、500、600 mg/L。以双蒸馏水为培养基作阴性对照,以‘默笙睡莲花露[19]作阳性对照(睡莲花朵第1天盛开时,柱头上会分泌花露,以接收昆虫所携带的花粉)。
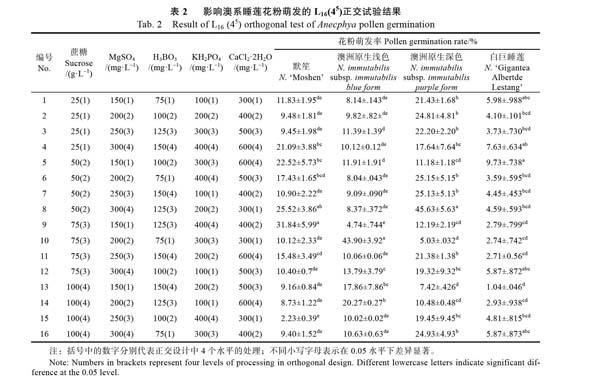
在单因素试验的基础上,以蔗糖(25、50、75、100 g/L)、MgSO4(150、200、250、300 mg/L)、H3BO3(75、100、125、150 mg/L)、KH2PO4(100、200、300、400 mg/L)、CaCl2·2H2O(300、400、500、600 mg/L)进行5因素4水平共16个组合L16(45)的正交试验。参试材料进一步扩增至4种,每种材料进行3次重复试验,依据花粉萌发率筛选出澳系睡莲花粉的最佳萌发培养基。
1.2.3 花粉保存 以上述干燥的花粉作为试验材料,每1 g分装于2 mL离心管内密封保存,分别置于常温(CK)、4、?20、?40、?70 ℃的密闭黑暗条件下保存。每隔24、48、72 h,取1支离心管的花粉进行离体培养萌发。分别选用4种澳系睡莲对应的最佳培养基,观测花粉的离体萌发率。试验操作步骤、培养时间与条件、统计方法与上述方法相同。每种澳系睡莲进行3次重复试验。
1.3数据处理
采用Image-Pro Plus 6.0軟件统计每个视野睡莲花粉的萌发率,采用Excel 2013 软件进行数据统计与制图,采用SPSS 18.0软件进行方差分析。
2 结果与分析
2.1不同培养基组分对花粉萌发率的影响
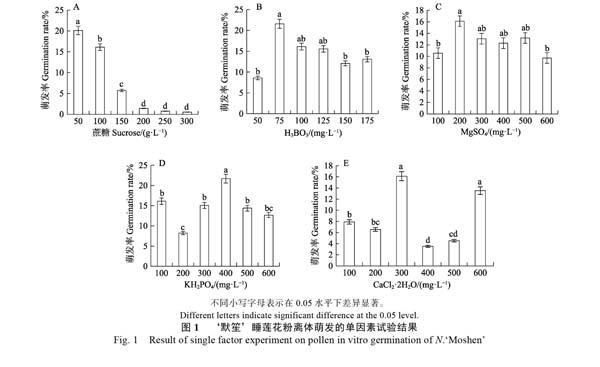
2.1.1 蔗糖浓度对花粉萌发的影响 阴性对照(双蒸馏水处理)的花粉萌发率为0;阳性对照(睡莲花露处理)的萌发率为14.33%。从图1A可以看出,低浓度的蔗糖更适宜‘默笙睡莲花粉的萌发,浓度在50 g/L时,花粉萌发率达到试验范围内最高值20.12%,高于阳性对照。随着蔗糖浓度的提高,花粉萌发率显著下降。方差分析结果表明,当蔗糖浓度达到200 g/L以上时,花粉萌发率较低,且无显著差异。依据单因素试验的结果,可将蔗糖浓度进一步精确调整至25、50、75、100 g/L进行正交试验。
2.1.2 其余因子的浓度对花粉萌发的影响 与不同浓度的蔗糖处理相比,H3BO3、MgSO4、KH2PO4、CaCl2·2H2O的浓度对‘默笙花粉萌发的影响规律不明显,不同浓度范围表现出不同的结果。
不同小写字母表示在0.05水平下差异显著。
Different letters indicate significant difference at the 0.05 level.
由图1B可知,H3BO3浓度为50 mg/L时,花粉萌发率最低(8.59%),当浓度上升至75 mg/L时,萌发率显著提升至最高(21.59%),之后随着H3BO3浓度的提高,萌发率缓慢下降。方差分析结果表明:当H3BO3浓度为75 mg/L时,花粉萌发率显著高于50、150、175 mg/L的H3BO3处理。
由图1C可知,MgSO4的浓度从100 mg/L提升至200 mg/L时,花粉萌发率显著提升,从10.57%提升至最高(16.11%)。当浓度超过200 mg/L以上时,‘默笙的花粉萌发率随着浓度的提高呈缓慢下降的趋势。方差分析表明:当MgSO4浓度为200 mg/L时,‘默笙花粉的萌发率显著高于100、600 mg/L MgSO4的处理。
由图1D可知,当KH2PO4的浓度在200~600 mg/L时,花粉萌发率呈现中间高、两头低的分布趋势。方差分析结果表明:当KH2PO4浓度为400 mg/L时,‘默笙睡莲花粉的萌发率显著高于其他处理,而100 mg/L处理的花粉萌发率与300、500、600 mg/L处理无显著差异。
由图1E可知,300、600 mg/L的CaCl2·2H2O适宜‘默笙花粉的萌发,萌发率分别为16.11%、13.52%。方差分析显示:300、600 mg/L CaCl2·2H2O的2个处理之间无显著差异,且花粉萌发率显著高于其他处理。
2.2正交试验各处理对澳系睡莲花粉萌发的影响
正交试验设计组合及结果如表2所示,各处理对4种澳系睡莲花粉萌发率的影响不同。其中,‘默笙的9号处理平均花粉萌发率最大,达到31.84%,显著高于除8号处理外的其他处理的花粉萌发率(P<0.05);澳洲原生浅色睡莲的10号处理与澳洲原生深色睡莲的8号处理的花粉萌发率均显著高于同类别中其他处理的平均花粉萌发率(P<0.05),平均花粉萌发率分别达到43.90%和45.63%;‘白巨睡莲的5号处理花粉萌发率相对较高,但与1号、4号、8号、16号处理的平均花粉萌发率差异不显著(图2)。
注:括号中的数字分别代表正交设计中4个水平的处理;不同小写字母表示在0.05水平下差异显著。
Note: Numbers in brackets represent four levels of processing in orthogonal design. Different lowercase letters indicate significant difference at the 0.05 level.
A:‘默笙花粉;B:澳洲原生浅色睡莲花粉;C:澳洲原生深色睡莲花粉;D:‘白巨睡莲花粉
A: The pollen ofN. ‘Moshen; B: The pollen ofN. immutabilis subsp. immutabilis blue form; C: The pollen ofN. immutabilis subsp. immutabilis purple form; D: The pollen ofN.‘Gigantea Albertde Lestang.
2.3不同保存条件对花粉离体萌发率的影响
由图3可知,随着花粉保存时间的增加,萌发率大幅度下降。?70 ℃保存的花粉24 h以后花粉萌发率下降至0。常温、4、?20、?40 ℃保存的花粉72 h以后萌发率下降至0。花粉在4 ℃条件下保存效果相对较好,24 h后的萌发率由31.84%下降至18.88%;48 h后,萌发率进一步降至4.32%。花粉在4 ℃条件下保存24 h和48 h时的萌发率均高于其他温度处理,表明‘默笙澳系睡莲的花粉相对适合在4 ℃条件下保存。
如图4所示,‘白巨睡莲花粉的初始萌发率较低,花粉随着保存时间的增加,萌发率呈下降趋势。?70 ℃条件下保存24 h,萌发率为0;常温条件下保存48 h,花粉无萌发;?4、?20、?40 ℃条件下保存72 h,萌发率降至0。花粉在4 ℃条件下保存24 h,萌发率小幅度下降,由9.73%降至9.05%;48 h后,萌发率大幅度降至2.61%。花粉在常温、?20、?40、?70 ℃下保存24 h,萌发率均大幅度下降,分别为2.71%、3.66%、2.08%、0。结果表明‘白巨睡莲在4 ℃条件下可保存24 h。
由图5可知,澳洲原生浅色睡莲的花粉随着保存时间的增加,萌发率大幅度下降,不同温度下保存24 h后,萌发率降至10%以下,48 h后均下降至2%以下,72 h后萌发率为0。花粉在4 ℃下处理效果相对略好,24 h后萌发率由43.90%下降至7.02%,略高于其他的温度处理;48 h后,萌发率降至1.65%。花粉在?20 ℃条件下保存24 h,萌发率由44.90%下降至7.02%,仅次于4℃的处理;48 h后,萌发率降至1.81%,略高于其他的温度处理。
如图6所示,澳洲原生深色睡莲花粉随着保存时间的增加,萌发率大幅下降。在常温、?20、?40、?70 ℃條件下保存48 h,萌发率下降至0。花粉在4 ℃下保存效果相对略好,萌发率略高于其他温度处理,但花粉生活力仍然大幅下降,保存至24 h,萌发率由45.63%下降至15.60%,48 h后萌发率进一步下降至1.52%,72 h后萌发率为0。
3讨论
蔗糖是影响植物花粉萌发的重要因素,为花粉萌发和花粉管生长提供能源物质,同时维持着花粉细胞外界环境的渗透压[20-22],H3BO3能增加花粉对糖的吸收、运转和代谢,形成糖硼酸复合体[23-24],Ca2+的动态平衡影响花粉的萌发与花粉管的顶端生长[25]。本研究中,蔗糖的单因素和正交试验结果表明,50、75 g/L的蔗糖配适宜的B3+、Ca2+、K+、Mg2+分别对4种澳系睡莲花粉萌发有较好的促进作用。但是B3+、Ca2+、K+、Mg2+的单因素试验与正交试验结果不完全吻合。例如,在单因素试验的BK培养基中,75 mg/L H3BO3处理的‘默笙花粉萌发率最高,而在正交试验的9号培养基中,125 mg/L H3BO3处理的‘默笙花粉萌发率更高,其原因可能是各因素存在交互作用,H3BO3与不同浓度的蔗糖、Ca2+、K+、Mg2+互作产生了不一样的效应。毛立彦等[13]对‘保罗兰睡莲花粉的研究发现,蔗糖、CaCl2、H3BO3对睡莲花粉萌发存在极显著的交互作用;常海龙等[26]研究表明,在硼酸、蔗糖、硝酸钙、硫酸镁4个因素中,蔗糖对甘蔗花粉萌发的影响最大,硼酸次之,它们对花粉管的生长最为关键。4种澳系睡莲花粉的最适培养基的各组分浓度不同,合理配比可以增加花粉的萌发率。通过花粉离体萌发法,可以直观、有效地评价花粉生活力,在睡莲的杂交育种过程中,对于父本材料的筛选有指导意义。在本研究中,‘白巨睡莲的花粉生活力较低,不适宜作为父本育种材料。
低温保存可以降低花粉呼吸作用、水分代谢、延长花粉生活力[27],是解决花期不育、异地授粉的主要手段。聂超仁等[28]研究表明,钟花樱花的花粉可在?20、?80 ℃条件下保存150 d仍保持20%以上的萌发率;吴月亮等[29]研究证明,?80 ℃超低温条件保存文冠果花粉1年,花粉萌发率接近50%,而常温条件下仅能保存15 d。本试验中,4种澳系睡莲的花粉在4 ℃下保存相对较好,但72 h后花粉均丧失生活力,仍然无法解决睡莲杂交育种中异地授粉、跨花期授粉等问题。
花粉含水量是影响花粉低温保存时间长短的因素,在低温冷冻过程中,含水量过高,在低温下细胞内结冰过多,造成细胞膜受损,影响保存后的生活力[30]。赵婵璞等[31]研究表明,未干燥的有斑百合花粉?80 ℃保存480 d后萌发率下降至2.3%,经过8 h干燥处理的花粉(含水量21.3%)?80 ℃贮藏720 d后,萌发率为20.7%。时婷婷等[32]发现,经干燥处理的‘索邦和‘西伯利亚百合新鲜花粉超低温保存420 d后的花粉萌发率分别为60.12%和58.02%,而CK(未干燥的花粉)萌发率仅为3.20%和2.21%。本试验没有考虑澳系睡莲花粉的含水量对花粉低温保存的影响,有待后续进一步研究。
参考文献
[1] 关克俭, 肖培根, 潘开玉, 等. 中国植物志: 第27卷[M]. 北京: 科学出版社, 1979: 8-11.
[2] 廖卫伟, 杨志娟, 朱天龙, 等. 睡莲的植物学性状及其分类[J]. 现代农业科技, 2016(24): 148, 154.
[3] 余翠薇, 陈煜初, 余东北, 等. 睡莲5个亚属花、叶、块茎和基因组大小比较[J]. 浙江农业科学, 2017, 58(8): 1353-1356, 1361.
[4] 李子俊. 我国首批跨亚属耐寒睡莲育成[J]. 中国花卉盆景, 2013(10): 4-7.
[5] 刘义满, Slearmlarp Wasuwat, 柯卫东, 等. 泰国睡莲考察报告[J]. 中国园艺文摘, 2009, 25(3): 120-124.
[6] 黄国振. 睡莲[M]. 北京: 中国林业出版社, 2009, 23-25, 45, 134-182.
[7] Preston R E. The intrafloral phenology ofStreptanthustortuosus (Brassicaceae)[J]. American Journal of Botany, 1991, 78(8): 1044-1053.
[8] 陆 琳, 彭绿春, 宋 杰, 等. 不同高山杜鹃品种花粉活力测定及贮藏方法研究[J]. 山西农业科学, 2016, 44(2): 175-178.
- 贾文庆, 王少平, 李纪元. 大花红山茶花粉形态特征和培养条件及其储藏过程的生理动态分析[J]. 西北植物学报, 2015, 35(4): 754-760.
- 王岚岚, 游 捷, 俞红强. 月季花粉离体萌发液体培养基组分的优化[J]. 河北农业大学学报, 2008(3): 42-45.
- 李千惠, 姜文龙, 徐同乐, 等. 日本海棠花粉最佳离体萌发培养基筛选及萌发过程研究[J]. 经济林研究, 2018, 36(2): 154-160.
- 杨梦飞, 李怡鹏, 楼宇涛, 等. 培养液组分及培养时间对睡莲花粉萌发和花粉管生长的影响[J]. 浙江农业科学, 2016, 57(10): 1689-1691.
- 毛立彦, 唐毓玮, 谢振兴, 等. ‘保罗兰睡莲花粉离体萌发及花粉管生长的研究[J]. 西南农业学报, 2019, 32(5): 1155-1161.
[14] 张慧会, 祝遵凌. 香水莲花花粉活力测定最适培养条件分析[J]. 分子植物育种, 2020, 18(2): 553-560.
- 李淑娟, 尉 倩, 陈 尘, 等. 中国睡莲属植物育种研究
进展[J]. 植物遗传资源学报, 2019, 20(4): 829-835.
[16] 王钦丽, 卢龙斗, 吴小琴, 等. 花粉的保存及其生活力测定[J]. 植物学通报, 2002(3): 365-373.
[17] 孙春丽, 潘延云. 拟南芥花粉活力的测定及其在花粉发育研究中的应用[J]. 植物学通报, 2008(3): 268-275.
[18] 张 瑞, 李 洋, 梁有旺, 等. 薄壳山核桃花粉离体萌发和花粉管生长特性研究[J]. 西北植物学报, 2013, 33(9): 1916-1922.
[19] 程 茵, 马光耀, 赵 莹, 等. 睡莲花露主要成分及抗氧化作用[J]. 食品研究与开发, 2019, 40(8): 108-114.
[20] 黄 静, 曹秋芬, 玉 平, 等. 培养基成分对南瓜离体花粉萌发的影响[J]. 中国西瓜甜瓜, 2004(3): 10-11.
[21] 杜纪红, 叶正文, 苏明申, 等. 桃花粉离体萌发和花粉管生长特性研究[J]. 西北植物学报, 2011, 31(1): 64-71.
[22] 邓衍明, 叶晓青, 梁丽建, 等. 茉莉花粉离体培养萌发及花粉管生长特性研究[J]. 华北农学报, 2014, 29(5): 107-113.
[23] Fang K F, Du B S, Zhang Q,et al. Boron deficiency alters cytosolic Ca2+concentration and affects the cell wall components of pollen tubes inMalus domestica[J]. Plant Biology, 2019, 21(2): 1-9.
[24] 朱江华, 吕芳德, 王 森, 等. 6个蓝莓品种花粉特性和花粉育性的比较[J]. 经济林研究, 2016, 34(2): 101- 108.
[25] 汤红明, 徐冬青, 徐根娣, 等. 植物花粉萌发的研究进展[J]. 安徽农业科学, 2006(24): 6436-6438, 6440.
[26] 常海龍, 张 伟, 陈俊吕, 等. 甘蔗花粉离体萌发研究[J]. 热带作物学报, 2019, 40(10): 2068-2075.
- 史锋厚, 范蓉蓉, 周 婷, 等. 垂丝海棠花粉贮藏特性研究[J]. 经济林研究, 2013, 31(4): 190-194.
- 聂超仁, 况红玲, 于静亚, 等. 钟花樱花粉离体萌发培养及低温保存技术研究[J]. 西南林业大学学报(自然科学), 2018, 38(1): 54-58.
- 吴月亮, 汤 鑫, 刘 迪, 等. 不同贮藏温度下文冠果花粉离体萌发研究[J]. 沈阳农业大学学报, 2018, 49(5): 600-604.
- 张亚利, 尚晓倩, 刘 燕. 花粉超低温保存研究进展[J]. 北京林业大学学报, 2006(4): 139-147.
- 赵婵璞, 张泉卫, 史宝胜, 等. 有斑百合花粉低温贮藏研究[J]. 河北农业大学学报, 2014, 37(1): 54-58.
[32] 时婷婷, 张桂玲, 温四民, 等. 百合花粉超低温保存方法研究[J]. 北方园艺, 2013(3): 64-66.
猜你喜欢
环球人物(2021年23期)2021-12-24
中国慈善家(2021年5期)2021-11-19
阅读(低年级)(2020年8期)2020-11-06
优雅(2020年8期)2020-09-16
散文诗(2020年2期)2020-07-20
小天使·一年级语数英综合(2017年11期)2017-12-05
作文周刊·小学三年级版(2016年15期)2017-05-26
诗潮(2017年2期)2017-03-16
幸福·悦读(2016年11期)2016-12-21
儿童故事画报(2016年4期)2016-06-24
