
日本沼虾原肌球蛋白线性表位预测研究
2020-09-23 12:15华希玮谢彦海陈红兵
食品工业科技 2020年17期
华希玮,谢彦海,陈红兵
(1.南昌大学食品科学与技术国家重点实验室,江西南昌 330047;2.南昌大学食品学院,江西南昌 330047;3.南昌大学实验动物科学中心,江西南昌 330006;4.南昌大学中德联合研究院,江西南昌 330047)
水产种类繁多,因其营养价值高而在全球被广泛食用,但其体内的过敏原蛋白能够引起严重的过敏反应。水产品过敏中由甲壳类引起的过敏最为普遍[1]。而虾类由于营养丰富、口感极佳,深受喜爱,故在甲壳类过敏中也有显著贡献[2]。常见的虾类过敏症状有荨麻疹、血管性水肿,也会影响到胃肠道及呼吸系统,严重者甚至发生过敏性休克[3]。源自甲壳类的过敏原有多种,有原肌球蛋白(Tropomyosin)、精氨酸激酶(Arginine kinase)、肌球蛋白轻链(Myosin light chain)和肌钙蛋白(Troponin)等,其中原肌球蛋白是主要的过敏原,通常能引起甲壳类动物之间以及甲壳类与其他无脊椎动物之间的交叉反应[4]。日本沼虾(Macrobrachiumnipponense),又称河虾,广泛分布于中国等亚洲国家淡水低盐度河口地区,它被认为是中国重要的渔业资源[5]。然而,目前国内外鲜有对日本沼虾原肌球蛋白抗原表位的分析研究。
抗原表位是致敏蛋白中决定抗原特异性的特殊化学基团,是过敏原与抗体结合的物质基础。表位可根据结构分为线性表位和构象表位;也可根据其结合受体细胞分为T细胞表位与B细胞表位[6]。B细胞表位可以是线性表位或构象表位,而T细胞表位一般为线性表位。在加工过程中蛋白质的结构变化,会导致抗原表位被破坏或者被掩盖,从而致敏性降低。一些加工方法例如酶解、超高压等会使部分食物致敏性降低甚至消失[7]。因此,针对过敏原表位的研究对于过敏原消减技术以及开发低致敏食品尤为重要。
研究表位的方法有多种,许多传统的表位鉴定方法经费成本较高,时间也较长。生物信息学的兴起拓展了对食物过敏原的研究方法,通过计算机技术与生物学数据库的结合,对过敏原蛋白的表位、同源性、过敏原之间的交叉反应性以及过敏原刺激机体产生免疫应答的能力进行预测[8]。
原肌球蛋白的空间结构较简单,多数研究认为在原肌球蛋白引起的免疫反应中起主导作用的是线性表位[9],对其抗原表位的报道也多为线性表位。因此,本研究利用生物信息学技术对日本沼虾原肌球蛋白进行分析并预测其抗原表位,以期为基于过敏原表位靶点的检测提供依据,为针对抗原表位的过敏原性消减提供新靶标。
1 材料与方法
1.1 材料
通过NCBI蛋白数据库(https://www. ncbi. nlm. nih. gov/protein/)检索日本沼虾原肌球蛋白的氨基酸序列(登录号为AHJ10946.1)。
1.2 实验方法
1.2.1 日本沼虾B细胞线性表位的预测 利用SOPMA[10](https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)对日本沼虾原肌球蛋白的二级结构进行预测。利用DNAStar软件中Protean模块,导入氨基酸序列,分别采用Chou-Fasman[11]法及Garnier-Robson法分析日本沼虾原肌球蛋白的二级结构,Karplus-Schulz[12]法分析柔韧性(Flexibility),Kyte-Doolittle[13]法分析亲水性(Hydrophilicity)、Emini[14]法分析表面可及性(Surface probability)、Jameson-Wolf[15]法分析抗原性指数(Antigenic index),从蛋白质多方面性质综合分析预测表位。分别在BepiPred 1.0 Server[16](http://www. cbs. dtu. dk/services/BepiPred-1.0/)和ABCpred[17](http://crdd.osdd.net/raghava/abcpred/ABC_submission. html)两个网站中输入蛋白序列来预测B细胞表位。综合分析不同方法的预测结果筛选出可能的B细胞线性表位。
1.2.2 日本沼虾T细胞表位的预测 在SYFPEITHI[18](http://www.syfpeithi.de/)网站的表位预测界面选择合适的MHC类型及肽段长度,输入氨基酸序列提交后根据得分预测T细胞表位。应用NetMHCII 2.3 Server[19](http://www.cbs.dtu.dk/services/NetMHCII/)与NetMHCIIpan 3.2 Server[20](http://www.cbs.dtu.dk/services/NetMHCIIpan/)两个服务器,输入氨基酸序列并选择相应基因型及氨基酸长度,预测蛋白质与MHC-Ⅱ类分子的结合能力,从而预测出T细胞表位。综合分析几个服务器的预测结果筛选出可能的T细胞表位。
1.2.3 日本沼虾与其它种类原肌球蛋白序列比对 通过NCBI蛋白数据库(https://www. ncbi. nlm. nih.gov/protein/)检索8种水产类原肌球蛋白的氨基酸序列,并利用ClustalX软件对这些不同种间的序列进行比对。褐美对虾(Penaeusaztecus),登录号为AAZ76743.1;中国对虾(Penaeuschinensis),登录号为ADA70137.1;斑节对虾(Penaeusmonodon),登录号为AAX37288.1;南美白对虾(Penaeusvannamei),登录号为ACB38288.1;刀额新对虾(Metapenaeusensis),登录号为AAA60330.1;锯缘青蟹(Scyllaserrata),登录号为ABS12233.1;章鱼(Octopusvulgaris),登录号为BAE54433.1;太平洋牡蛎(Crassostreagigas),登录号为BAH10152.1。
2 结果与分析
2.1 日本沼虾原肌球蛋白B细胞线性表位预测
2.1.1 二级结构的预测结果 从SOPMA对日本沼虾原肌球蛋白的二级结构预测中发现,该蛋白质以α-螺旋(Alpha helix)结构为主,伴随少许无规则卷曲(Random coil)结构(图1A)。通过DNAStar软件中Protean模块中的两种方法对蛋白质的二级结构预测(图1B),发现Garnier-Robson法得到的结果与前者几乎一致,而Chou-Fasman法结果表明还存在一小部分β-转角(Beta turn)结构。有研究表明原肌球蛋白是两个α-螺旋多肽链相互缠绕形成的超螺旋结构[21]。由此可见,预测得到的二级结构与文献报道基本一致。在蛋白质二级结构中,α-螺旋一般位于蛋白质内部,不易与抗体结合,β-转角与无规则卷曲多位于蛋白质表面,更容易与抗体结合,有较大可能成为表位[22]。因此,不能通过日本沼虾原肌球蛋白的二级结构准确地预测出表位,而需要结合亲水性、抗原指数等其他性质综合分析。

图1 通过SOPMA与Protean预测的日本沼虾原肌球蛋白的二级结构
2.1.2 基于Protean的预测结果 应用DNAStar的Protean预测蛋白质结果如图2所示,图2中显示了蛋白质的亲水性、柔韧性、抗原指数和表面可及性。从图2中可以发现该蛋白质亲水性非常高,几乎整个蛋白质的亲水指数>0,蛋白质的亲水基团多位于蛋白质表面,更容易形成与抗体等结合的抗原表位;柔韧性分析结果显示该蛋白质柔韧性区域较多且范围较大,由于具有柔韧性的区域发生改变的几率较高,容易与抗体契合,形成表位的可能性较大;从抗原指数结果而言,该蛋白质大部分区域抗原指数>0,抗原指数较高更容易形成表位;对表面可及性进行分析,以表面可及性指数>1作为选择标准,这些区域呈现于蛋白质表面,具有与抗体结合的可能性,更易形成表位[23]。综合考虑,预测出的日本沼虾原肌球蛋白B细胞线性表位如表1所示。

图2 通过Protean预测的日本沼虾原肌球蛋白的结果
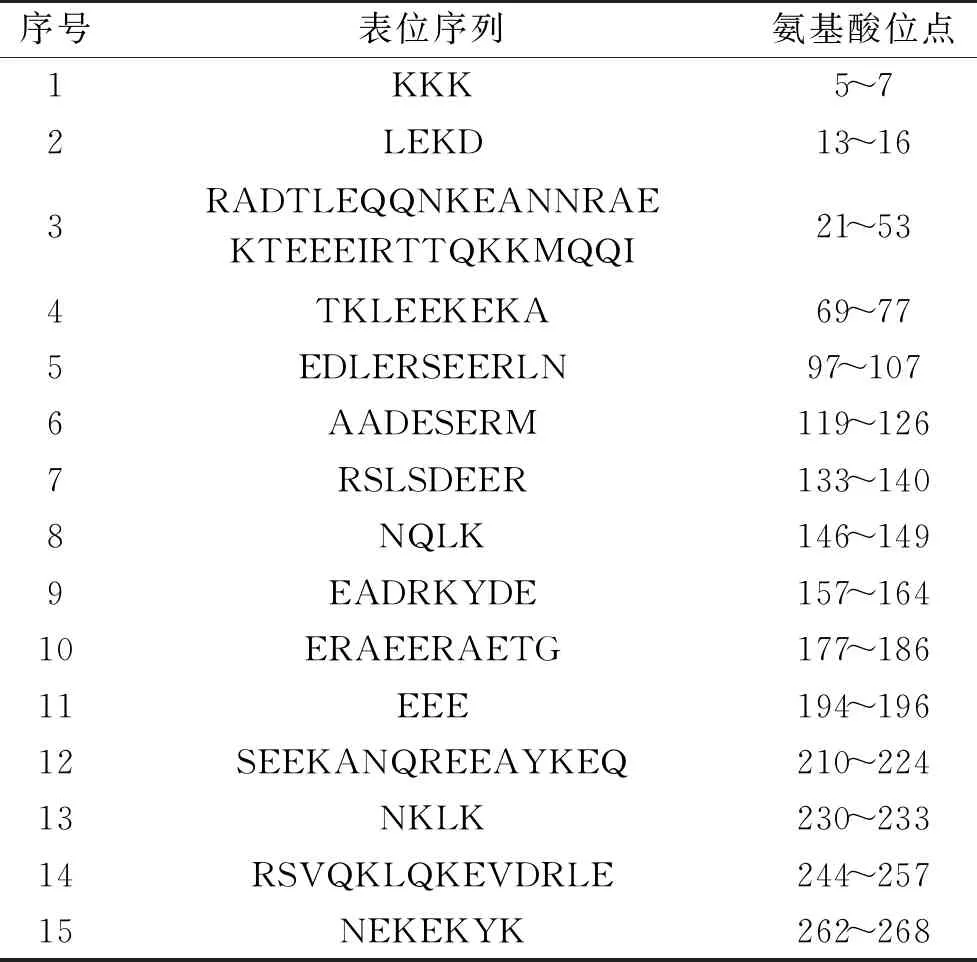
表1 通过Protean预测的日本沼虾原肌球蛋白表位
2.1.3 基于BepiPred的预测结果 BepiPred 1.0 server采用了隐马尔可夫模型和倾向标度法,结合氨基酸性质进行表位预测,表位分配的分数阈值设置为默认值(0.35),得分高于0.35则被认为是可能的B细胞线性表位[22,24]。BepiPred预测日本沼虾原肌球蛋白中所有得分高于0.35的区域序列如表2,可以发现预测区域平均分布在整个蛋白序列上。

表2 通过BepiPred预测的日本沼虾原肌球蛋白表位
2.1.4 基于ABCpred的预测结果 ABCpred基于标准前馈与递归神经网络开发[24]。运用700个 B细胞表位和700个最大长度为20的非B细胞表位的随机肽的数据库,在不同的输入窗口长度和隐藏单元下对网络进行了训练和测试,经交叉验证,结果显示递归神经网络及窗口长度为16时准确度达到65.93%[17]。选择16个氨基酸的长度以及阈值0.7作为标准来筛选表位。预测的B细胞表位根据训练后的递归神经网络得到的分数进行排序,肽得分越高,作为表位的概率越高。所有得分高于阈值的肽排列如表3。

表3 通过ABCpred预测的日本沼虾原肌球蛋白表位
2.1.5 综合分析B细胞线性表位 B细胞线性表位预测通常是依据氨基酸序列信息,分析二级结构、亲水性、柔韧性、表面可及性、序列保守性、残基失序等,利用机器学习方法对Bcipep、AntiJen等数据集进行训练,将蛋白质的氨基酸序列输入模型中进行鉴定[24-25]。综合Protean、BepiPred以及ABCpred的预测结果,将重复区域作为潜在抗原表位,得到了共有的10个潜在B细胞线性抗原表位,分别是21RADTLEQQNKEANN34、37EKTEEEIRTTQKKMQQ52、71LEEKEKA77、99LERSEERLN107、119AADESER125、134SLSDEER140、158ADRKYDE164、177ERAEERAETG186、210SEEKANQREEAYKE223、262NEKEKYK268。此前已有一些研究利用生物信息学预测得到相应的B细胞线性表位并进行了验证。李雪娇等[26]利用生物信息学软件DNAStar中的Protean预测出4个CM16的B细胞线性表位,与来自CM16的3条抗消化肽段的序列进行比对分析后,发现与其中2条肽段存在部分重合。Fu等[27]利用多种免疫信息学工具综合预测得到中国对虾原肌球蛋白与精氨酸激酶的线性表位后用Fmoc法固相合成多肽,经间接竞争ELISA验证,准确率分别高达83%与70%。由此可见,利用生物信息学预测B细胞线性表位具有一定的可行性与可靠性。不过,这些方法也存在一定的局限性。表位预测工具使用的数据集不够完善导致预测结果不够精准,不同工具使用的算法与模型也不尽相同,致使预测结果存在一定差异,因此均不能得出标准的预测结果[24]。虽然综合分析不同方法的预测结果可以提高预测的准确率,但还是需要实验进一步验证。目前较为常见的方法是将预测的表位合成多肽后与过敏患者血清采用Dot-blot等方法进行初步鉴定[27-28]。
2.2 日本沼虾原肌球蛋白T细胞表位的预测
2.2.1 基于SYFPEITHI的预测结果 SYFPEITHI为定性分析,它采用结合基序的方法对多肽与MHC-Ⅱ类分子的结合能力进行评分,用于筛选可能的T细胞表位[29]。在页面选择HLA-DRB1基因型以及15mers的氨基酸残基长度,输入序列后可以得到预测结果。若肽段得分超过25分,则认为其与MHC-Ⅱ类分子结合力较强,是潜在T细胞表位[30]。预测结果如表4所示。该蛋白对表中几种基因型都表现出较强的结合力,其中HLA-DRB1*0401基因型预测发现较多的表位,因此该基因型人群对该蛋白序列表现出更高的敏感性。

表4 通过SYFPEITHI预测的日本沼虾原肌球蛋白表位
2.2.2 基于NetMHCII与NetMHCIIpan的预测结果 NetMHCII和NetMHCIIpan已被证明是预测多肽与MHC-II类分子结合亲和力的准确率较高的方法[31]。这两种方法都是基于来自IEDB的数据及人工神经网络,使用NNAlign框架进行训练的[32-33]。它们的主要区别之一是,NetMHCII是每个MHC分子的独立网络的集合,而NetMHCIIpan则包含一个单一的通用网络,可以预测已知蛋白质序列的所有MHC分子的肽结合亲和力[32]。研究者开发的NetMHCIIpan-3.2与NetMHCⅡ-2.3服务器,则将NetMHCII和NetMHCIIpan预测范围扩大并可以显著地提高预测准确性。MHC-Ⅱ类分子能结合长度达10~28个氨基酸残基的表位,最佳长度为12~16个氨基酸残基[34]。因此,应用这两个服务器,选择中国人群中常见的HLA等位基因[35-36]以及15的肽段长度进行T细胞表位预测。Affinity(nM)<50即该肽段与MHC-II类分子结合呈强亲和力,50 表5 通过NetMHCII与NetMHCIIpan预测的日本沼虾原肌球蛋白表位 图4 不同物种间原肌球蛋白序列比对及T细胞表位对比 2.2.3 综合分析T细胞表位 外源性蛋白质抗原一般经抗原递呈细胞处理后由MHC-Ⅱ类分子递呈给T细胞[38]。因此,预测蛋白质的T细胞表位可通过评估MHC-Ⅱ类分子与多肽的结合能力来实现。在人体中负责编码相关分子的HLA-Ⅱ类基因主要包括DP、DQ、DR三个亚区[39]。因HLA基因组高度多态性,故选择中国人群中常见的HLA等位基因型进行预测。综合SYFPEITHI、NetMHCII和NetMHCIIpan的预测结果,选取重复区域作为潜在表位,得到了共有的5个潜在T细胞抗原表位,分别是82EGEVAALNRRIQLL95、105RLNTATTKLAEAS117、165VARKLAMVEADLE177、195EELRVVGNNLKSLE208、222KEQIKTLTNKLKAA235。T细胞表位预测在基础研究中可以减少工作量,降低成本,同时,一些准确性较高的预测工具的预测结果也能得到验证。伍慧妍等[30]利用NetMHCII预测得到牡蛎原肌球蛋白的T细胞表位,发现与前人经实验鉴定的表位几乎一致。在目前的T细胞表位预测工具中,预测多肽与MHC-Ⅱ类分子的结合位点和结合亲和力是预测与MHC-Ⅱ类分子结合的表位的主要方法。然而大多数方法仅考虑表位的核心结合序列,而忽略了两侧氨基酸残基及TCR对结合力的影响[29]。因此,表位预测结果可以通过合成多肽后进行淋巴细胞增殖实验、免疫保护实验等来进一步验证[38]。 2.3.1 日本沼虾原肌球蛋白B细胞表位对比 日本沼虾与不同物种序列比对结果如图3。从图3中可以发现这几种原肌球蛋白序列相似度很高,虾蟹类之间序列相似度均高于80%,而虾蟹类与软体动物之间相似度为60%~80%,由此可见,原肌球蛋白具有较高的保守性。一些学者已经鉴定出几种水产原肌球蛋白的表位[27-28,40-45]。这些表位多位于保守区内,且不同种间表位有部分或完全重叠。此次预测的日本沼虾原肌球蛋白B细胞表位也基本位于保守区内,且与已鉴定的表位均有重叠,其中预测的表位4与表位6重叠部分最多,因此该区域更有可能发生交叉反应。 图3 不同物种间原肌球蛋白序列比对及B细胞表位对比 2.3.2 日本沼虾原肌球蛋白T细胞表位对比 原肌球蛋白相关的T细胞表位研究不多,然而T细胞在免疫过程中也起着重要作用。Wai等[46]合成18个重叠肽,利用肽段刺激致敏小鼠中分离出的脾细胞,通过其增殖与细胞因子的反应鉴定T细胞表位。Ravkov等[47]综合体外的MHC-肽结合实验及体内的增殖及细胞因子释放实验鉴定了17个T细胞表位。此次预测的日本沼虾原肌球蛋白T细胞表位与已鉴定的表位均有重叠,其中表位5与鉴定表位完全一致,其他4个表位也均位于保守区。 本研究在NCBI蛋白数据库中找到日本沼虾原肌球蛋白的氨基酸序列,运用DNAStar、BepiPred和ABCpred对日本沼虾原肌球蛋白进行B细胞线性表位预测,最终定位出潜在的B细胞线性表位有10个肽段。同时,应用SYFPEITHI、NetMHCII和NetMHCIIpan分别对序列与MHC-Ⅱ类分子结合力评估,最终筛选出可能的T细胞表位有5个肽段。预测的表位与已经鉴定的表位均有部分重叠,这可能一定程度上会导致交叉反应。总之,本研究利用生物信息学技术预测出了日本沼虾原肌球蛋白的B细胞线性表位及T细胞表位,将有助于对日本沼虾过敏原的认识,为未来基于表位的相关研究提供了新的研究靶标。另外,日本沼虾原肌球蛋白的准确的表位以及优势表位还有待实验进一步确证。

2.3 日本沼虾与不同种间原肌球蛋白序列比对

3 结论
猜你喜欢
当代水产(2022年7期)2022-09-20
当代水产(2022年6期)2022-06-29
食品科学(2022年7期)2022-05-13
现代食品科技(2022年4期)2022-04-28
当代水产(2021年8期)2021-11-04
生命科学研究(2021年4期)2021-09-08
食品工业科技(2021年3期)2021-06-16
温州医科大学学报(2019年4期)2019-04-28
海洋与渔业(2017年11期)2018-01-26
中国免疫学杂志(2017年1期)2017-01-17
