
湘西喀斯特地区人工林树种的生长规律及生产力
2020-10-20 05:58刘立玲朱宁华
江苏农业科学 2020年18期
刘立玲 朱宁华

摘要:石漠化地区人工林培育与经营技术是当今林业亟需解决的问题之一。以湖南省湘西自治州青坪国有林场的35年南酸枣、40年台湾杉及40年香椿3个树种的试验林为研究对象,分别设置标准地展开系统调查研究,对标准地标准木进行树干解析,分析其生长规律,推算林分生物量。结果表明,南酸枣、台湾杉、香椿单木胸径、树高及材积的总生长量均随树龄的增加而增加;3个树种的林分生产量以乔木层占比最大,乔木层林分生产力从大到小排列为南酸枣(5194 t/hm2)>香椿(49.48 t/hm2)>台湾杉(49.33 t/hm2),3个树种各器官的生产力均表现为树干>树根>树枝>树叶。通过对这3个树种生长过程的量化研究,对各树种生长的速生期及数量成熟期有所了解,可为提高林木生产力提供理论依据,同时也为湘西喀斯特地区南酸枣、台湾杉及香椿人工林的科学经营与管理提供一定的理论基础,从而推动喀斯特地区人工林的发展。
关键词:南酸枣;台湾杉;香椿;生长规律;生产力;喀斯特地区;胸径;树高;材积
中图分类号: S718.43文献标志码: A
文章编号:1002-1302(2020)18-0165-06
收稿日期:2019-11-28
基金项目:中南林业科技大学研究生科技创新基金(编号:CX20192026)。
作者简介:刘立玲(1994—),女,陕西安康人,硕士研究生,主要从事石漠化植被恢复相关研究。E-mail:1041280341@qq.com。
通信作者:朱宁华,博士,副教授,主要从事森林培育相关研究。E-mail:225429@qqq.com。
南酸枣(Choerospondias axillaris)为漆树科南酸枣属落叶阔叶乔木,高度可达20 m,具有落叶量大、生长快的特点,是较好的速生造林树种,具有较高的生态价值和经济价值。台湾杉(Taiwania cryptomerioides)为杉科台湾杉属高大常绿乔木,分布在中亚热带季风气候区,属于国家一级保护植物,为优良珍贵的用材树种和庭园绿化树种[1]。香椿(Toona sinensis)为楝科香椿属高大落叶乔木,雌雄异株,其嫩叶可供食用,树干通直,素有“中国桃花心木”之称,不但是珍贵的用材树种,也是园林绿化的优选树种,具有很高的经济价值和开发前景[2]。南酸枣、台湾杉和香椿是湖南省湘西自治州森林生态次生林的主要造林树种。
目前,学者已经对南酸枣地理种源苗期性状变异[3-4]、次生林幼林配方施肥[5]、南酸枣家系叶及生理性状[6]等方面进行了研究;对于台湾杉的研究内容主要偏重于种群遗传结构[7-8]、次生代谢产物的鉴定[9]、叶挥发油的成分及其生物活性[10]等方面;而对香椿的研究主要集中在香椿花粉、种子的萌发及活力[11]、香椿种子在特定贮藏条件下活力的变化[12]等方面。目前,关于以上3个树种次生林生长规律及生物量的研究均较少。因此,本研究根据3个树种各自的胸径、树高、材积生长量,获得它们的生长规律,通过对这3个树种单木生长规律及生物量的分析来估测它们的林分生物量[13-14],旨在为培育高品质林分提供理论基础。
湖南省湘西自治州青坪镇土壤属于典型的岩溶性土壤,生态系统脆弱,植被种类单一,生态系统结构和功能稳定性差,土地生产力低下,基岩大面积裸露,因此该地区属于生态造林困难地区。要想培育品质较好的林分,就需要围绕森林的生物量和生产力进行研究。有研究者通过对湘西自治州永顺县青坪国有林场南酸枣、台湾杉及香椿人工林测树因子各年间的变化动态进行测定,拟合了相应的生长曲线[15-16]。了解和掌握南酸枣、台湾杉、香椿的树高、胸径及材积生长规律,合理调整林分结构,促进林木生长,可为林木快速生长期提供科学的抚育管理,从而为进一步深入我国亚热带岩溶地区造林树种的研究提供参考。
1 材料与方法
1.1 试验地概况
试验地位于湖南省湘西自治州青坪国有林场,地理位置为27°44.5′N,109°10′E。试验地大陆性季风气候特征明显,境内年降水量为1 300~1 500 mm,年均温度为16.3 ℃,年无霜期为269~292 d,年均日照时数为1 340 h。该地的母岩主要以石灰岩为主,土壤主要是红壤和黄棕壤,土层厚度不一。各样地人工林林下的植被比较丰富,其中草本层以天葵、兰草、蕨类为优势种,此外還分布着少量沿阶草、紫菀、金银花等,平均高度为0.17 m;灌木层以蔷薇、悬钩子、油茶为优势种,还分布着少量珊瑚樱等,平均高度为0.85 m;林下植被覆盖度约为30%,枯落物层厚度约为2~3 cm。
1.2 样地设置与数据测量
于2019年1月对湘西自治州青坪国有林场的南酸枣、台湾杉及香椿人工林进行全面的林分踏查,根据典型选样原则,在3个树种林分内各设置1个最具代表性的标准地,大小为20 m×20 m,对标准地进行每木检尺,同时记录相关测树因子及每个样地的海拔、坡向、坡位及林下植被。
根据实地调查结果,选取样方内能够代表林分生长水平的平均木作为解析木,3个树种各选1株,共计3株。用树干解析法、林木分层收获法、样方收获法获得各林分的林木生物量及生产力实物量[17]。标准木伐倒后,在0、1.3 m,以后按2.0 m间隔的区分段分别取1个约5 cm厚的圆盘,带回室内进行树干解析,分析解析木胸径、树高、材积的生长规律[18]。标准地的林分结构特征见表1。
1.3 生物量调查
地上部分样本采用分层切割收获法采集,树干以2 m为区分段,测定各区分段树干、树皮鲜质量,地下部分采用全挖法[19]采集。各部分均采集样本500 g。
林下植被采用样方收获法采集,在标准地内各设置5个面积为2 m×2 m的灌木小样方,5个面积为1 m×1 m的草本小样方,调查并记录小样方中的物种类别、数量、高度、盖度和枯落物厚度等因素,分别测定灌木层、草本层、凋落物层的鲜质量。将样品带回实验室后,于105 ℃杀青处理30 min后,再于85 ℃恒温烘干至恒质量,测定各样品的含水率,以推算各部分的干物质量[20-21]。
1.4 数据统计与分析
用Excel 2013、SPSS 20.0等软件整理解析木数据,统计各器官生物量,计算单株和林分生产力,并绘制林木生长规律图。
2 结果与分析
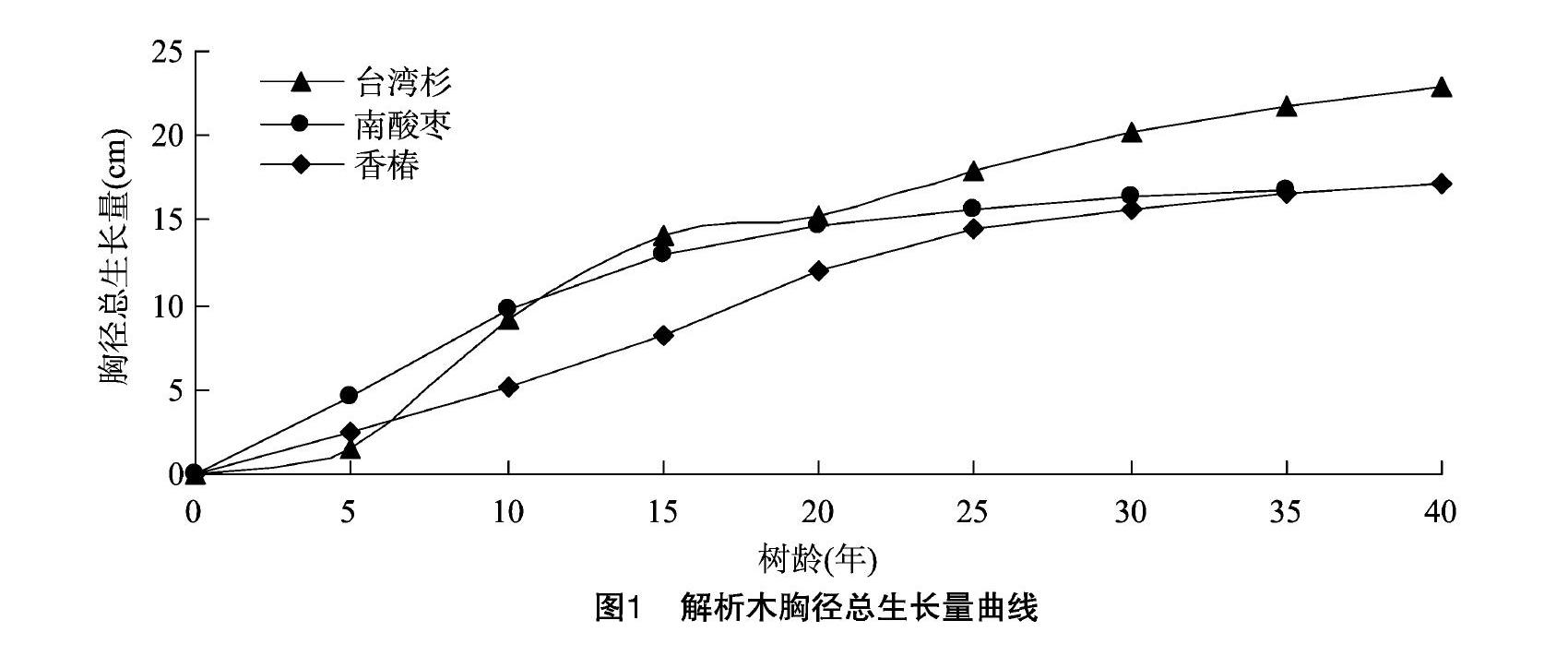
2.1 解析木胸径生长过程分析
解析木的胸径生长过程如图1、图2所示。南酸枣在树龄达到35年时,胸径总生长量达到 16.8 cm,此时平均胸径生长量、连年胸径生长量分别为048、0.08 cm。树龄为10年时,南酸枣胸径连年生长量达最大值,为1.06 cm,连年胸径生长曲线与平均胸径生长曲线在树龄为10~15年时出现相交。40年生台湾杉的胸径总生长量达到 22.95 cm,此时平均胸径生长量、连年胸径生长量分别为0.57、0.23 cm,其连年胸径生长量分别在树龄为10、25年时出现峰值;连年生长曲线与平均生长曲线在树龄在16年时出现相交。40年树龄的香椿胸径总生长量为17.15 cm,树龄为20年时胸径连年生长量达到峰值,为0.76 cm。
2.2 解析木树高生长规律
由图3、图4可知,3个树种的树高总生长量均随着树龄的增长而增加。35年树龄的南酸枣树高总生长量达到15.52 m,树高于树龄为5~10年时生长得最快,处于快速生长期,此后进入稳定生长期,其树高的连年生长量范围为0.05~1.23 m,波动较大;树龄为10年的南酸枣树高连年生长量、平均生长量均达到峰值,分别为1.33、128 m。树龄为40年的台湾杉树高总生长量达2122 m,台湾杉的树高连年生长量出现3次峰值,分别出现在10、25、35年树龄,最大值为10年树龄的 1.14 m;15年树龄的台湾杉树高平均生长量达到峰值,为 0.91 m,15年以后台湾杉树高的生长速度减缓。树龄为40年的香椿树高总生长量达23.50 m,其树高连年生长量出现2个生长高峰,分别在树龄5、20年时,25年树龄的树高平均生长量与连年生长量第2次相交,此后香椿树高平均生长量保持在0.72 m左右。
2.3 解析木材积生长规律
如图5、图6所示,3个树种单株材积总生长量随着树龄的增长而增加。35年生的南酸枣的单株材积总生长量为0.184 6 m3,材积的平均生长量和连年生长量均随着树龄的增加呈现先增加后下降的趋势,材积连年生长量在树龄为15年时达到峰值,为0.009 67 m3;在树龄为25~30年时,材积的平均生长量和连年生长量相交,此时林分达到主伐年龄。树龄为40年的台湾杉材积总生长量达最大值,为0.396 1 m3,材积连年生长量在树龄为5~25年间不断增加,在树龄为25年时达到峰值,为0194 2 m3。台湾杉材积连年生长量和平均生长量曲线一直未相交,材积生长成熟期至少在树龄为40年之后。40年生香椿的材积总生长量为 0.253 9 m3,材积连年生长量随树龄增加呈现先增后减的趋势,香椿材积的平均生长量一直随着树龄的增加而缓慢增加,当树龄为25年时,材积的连年生长量达最大值,为0.012 0 m3;树龄达30年之后,材积平均生长量保持在 0.007 6 m3,处于平稳增长状态。香椿材积的生长高峰比树高、胸径的生长高峰出现得晚;在树龄为0~25年范围内,材积连年生长处于一个快速增长的状态,在树龄为25~40年范围内,材积的连年生长量增加速度明显减缓。材积的连年生长量和平均生长量相交于树龄为38年时,此时林分达到数量成熟,已达主伐年龄。
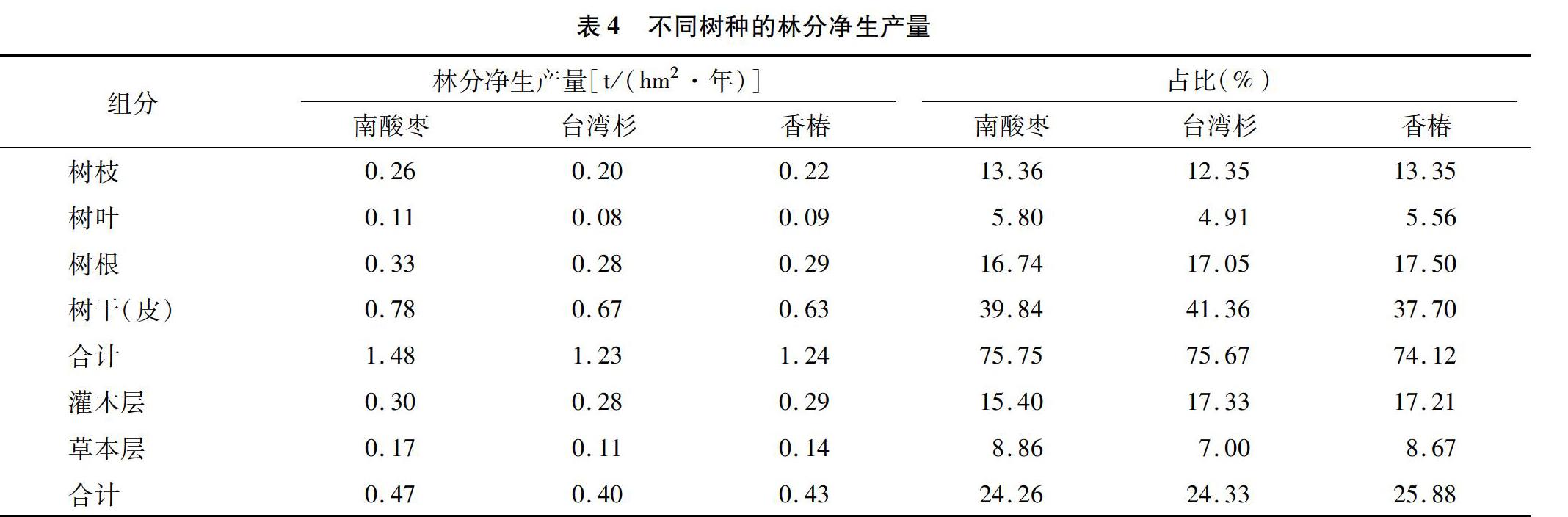
2.4 林分生物量分析
由表2可知,解析木单株各器官生物量的分配比例在不同树种间差异明显。3个树种的单株生物产量排序为台湾杉(515.11 kg/株)>香椿(297.50 kg/株)>南酸枣(205.06 kg/株)。3个树种中地上部生物量占单木生物量比例最大的是树干,均大于50%,其次是树枝生物量,约占单木生物量的17%,而树叶生物量只占单木生物量的极小部分。3个树种地下部分根生物量占单木生物量的比例约为23%。
将实测得到的标准地的标准木生物量乘以该样地的林木数,即得到林分生物量[22]。由表3可见,虽然树种不同,但是各树种林分生物产量情况却相同。3个林分乔木层的生物量排序均为干(皮)生物量>根生物量>枝生物量>叶生物量。在3种林分林下,灌木生长良好,均以蔷薇、悬钩子、茶树
为优势种,灌木层的林分生物量分別为10.56、1130、11.49 t/hm2,均占全林分的17%左右。与乔木层、灌木层相比,草本层生物量只占林分生物量的很少一部分,占比在0.3%以下,虽然占比较小,但是草本层也是森林生态系统的重要组成部分,在生物多样性及涵养水源方面发挥着巨大的生态作用。
每年林木通过光合作用生产有机物质,除去呼吸消耗的部分,剩余部分称为净生产量[23]。由表4可知,在3个树种的林分中,乔木层的年平均净生产量排列为南酸枣[1.48 t/(hm2·年)]>香椿[124 t/(hm2·年)]>台湾杉[1.23 t/(hm2·年)]。3个树种各器官年平均净生产量排序均为树干>树根>树枝>树叶;林分净生产量占比中乔木层最高,其次是灌木层,3个树种林分对应的灌木层年平均净生产量占比为16%左右,草本层占比最小,均在10%以下。
3 结论
本研究结果表明:(1)3个树种胸径、树高及材积生长量均随着树龄的增长而增加,35年生南酸枣的胸径总生长量为16.8 cm,40年生台湾杉、香椿的胸径总生长量分别为22.95、17.15 cm。由此可见,在石漠化地区,3个树种的胸径生长状况以台湾杉最优。35年生南酸枣树高总生长量为15.52 m,40年生的台湾杉、香椿树高总生长量分别为21.22、23.50 m,相比较而言,香椿的树高生长得较快。(2)3个树种林分生产量以乔木层占比最大,林分乔木层生产力由大到小排列如下:南酸枣(51.94 t/hm2)>香椿(49.48 t/hm2)>台湾杉(4933 t/hm2)。乔木层部分以树干的生产力最高,均占总生产力的40%以上,3个树种各器官生产力排序均为树干>树根>树枝>树叶。
4 讨论
本研究分3个方面对湘西自治州石漠化地区南酸枣、台湾杉和香椿3个树种的生长规律进行分析得出:(1)由树干解析的数据可知,3个树种的胸径总生长量均随年龄的增长而增加,35年生南酸枣人工林胸径的总生长量为16.8 cm,40年生台湾杉、香椿胸径的总生长量分别为22.95、17.15 cm。相比之下,该地区3个树种胸径生长量较其他地区生长量稍有逊色,可能是由于该石漠化地区人工林群落结构比较单一、土壤贫瘠、土层厚薄不均匀,缺乏促进植物生长的氮、磷、钾等元素,根系生长受阻,导致胸径生长量偏小,生产力较低。(2)35年生南酸枣树高总生长量为15.52 m,40年生台湾杉、香椿的树高总生长量分别为21.22、23.50 m,相比较而言,香椿的树高生长得较快,可能是随着林木的生长发育,林分郁闭度逐渐增大,个体之间竞争明显,生长空间不足,连年生长量增加速度明显减缓,导致其生长出现差异。(3)3个树种的材积总生长量均随年龄的增长而增加,35年生南酸枣材积连年生长量在树龄为15年时达到峰值,为0.009 67 m3,连年生长量与平均生长量在树龄为28年时相交,此时已达南酸枣人工林材积生长成熟期。40年生的台湾杉材积连年生长量在树龄为25年时达到峰值,为0194 2 m3,材积连年生长量与平均生长量曲线一直未相交,说明台湾杉材积生长成熟期至少在40年之后。40年生香椿材积连年生长量在树龄为25年时达到最大值,为 0.012 0 m3,其材积的连年生长量和平均生长量相交于树龄为38年,此时林分达到数量成熟,林分达主伐年龄。因此可见,在3种林分的快速生长期应加强水肥管理和适当间伐,控制好林分密度[18],充分利用其快速生长期,有效促进其树高和胸径的快速生长,从而提高生产力。
根据3个树种表现出的林分生产力可以看出,南酸枣在喀斯特地区林分生产量最大。由于南酸枣和香椿为阔叶树,台湾杉为针叶树,阔叶树枝干叶比针叶树发达,加上南酸枣本身的生物学特性,材质柔韧,收缩率小,所以尽管南酸枣树龄较台湾杉小,但是其林分的生产量反而大于台湾杉。因此,在进行喀斯特地区植被恢复时,可以考虑优先选择南酸枣、香椿等阔叶树或针阔多形式混合造林,根据该地区小生境,进行乔灌草搭配,合理地配置密度,充分利用林地内小气候环境的有利条件,促进其生物量的积累[24]。本研究选取标准木法测算乔木生物量、反向推演林分生长过程,今后可扩大径阶分布范围,通过建立不同的生长模型,开展年度追踪观测,为乔木层生物量估测提供依据,更深入地去研究喀斯特地区主要树种的生長规律及生物量分配,加上人工林的科学经营,最终实现湘西自治州石漠化地区的生态环境恢复与生态经济可持续发展。
参考文献:
[1]李江伟. 基于NSSR和CPSSR标记的台湾杉遗传多样性研究[D]. 武汉:华中农业大学,2014:4-5.
[2]彭方仁,梁有旺. 香椿的生物学特性及开发利用前景[J]. 林业科技开发,2005,19(3):3-6.
[3]骆文坚,何贵平,陈益泰,等. 南酸枣地理种源幼林生长性状变异和种源选择[J]. 江西农业大学学报,2007,29(3):365-371.
[4]何贵平,陈益泰,孙银祥,等. 南酸枣地理种源苗期性状变异研究[J]. 林业科学研究,2003,16(2):177-182.
[5]童 琪,李志辉,童方平,等. 南酸枣幼林配方施肥及叶片氮磷钾元素分布规律[J]. 中南林业科技大学学报,2017,37(5):50-54.
[6]区锦玮,李光友,徐建民,等. 南酸枣家系叶及生理性状的研究[J]. 中南林业科技大学学报,2017,37(4):75-78,99.
[7]王加国,李晓芳,安明态,等. 雷公山濒危植物台湾杉群落主要乔木树种种间联结性研究[J]. 西北林学院学报,2015,30(4):78-83.
[8]李江伟,杨琴军,刘秀群,等. 台湾杉遗传多样性的ISSR分析[J]. 林业科学,2014,50(6):61-66.
[9]Chiang N T,Ma L T,Lee Y R,et al. The gene expression and enzymatic activity of pinoresinol-lariciresinol reductase during wood formation in Taiwania cryptomerioides Hayata[J]. Holzforschung,2019,73(2):197-208.
[10]龚玉霞,张文慧,姜自见,等. 台湾杉叶挥发油的成分及其生物活性[J]. 江苏农业科学,2008,34(5):235-236.
[11]胡继文,肖 遥,翟文继,等. 香椿花粉离体萌发与花粉生活力研究[J]. 林业科学研究,2019,32(2):160-165.
[12]张海波,杨桂娟,高卫东,等. 香椿种子特定贮藏条件下活力变化的研究[J]. 林业科学研究,2019,32(2):152-159.
[13]孟 钢. 喀斯特地区灯台树人工林生长规律及林分地力研究[D]. 长沙:中南林业科技大学,2017:12-14.
[14]董利虎,李凤日. 三种林分生物量估算方法的比较[J]. 应用生态学报,2016,27(12):3862-3870.
[15]黄石嘉,李铁华,文仕知,等. 青冈栎的生长规律及生物量分布格局研究[J]. 中南林业科技大学学报,2017,37(3):57-62.
[16]谭桂菲,武建云,吴际友,等. 15年生香樟人工林生长规律研究[J]. 广西林业科学,2018,47(1):41-46.
[17]吕曼芳. 珍贵树种顶果木人工林生长规律及价值核算研究[D]. 南宁:广西大学,2013:13-15.
[18]林立彬,李铁华,文仕知,等. 闽楠木荷混交幼林生长规律及生物量分布特征研究[J]. 中南林业科技大学报,2019,39(4):79-84,98.
[19]罗文婷. 青坪生态站南酸枣林分生长及养分特征研究[D]. 长沙:中南林业科技大学,2014:9-11.
[20]赵 樟. 南亚热带杉木、红锥人工林的生物量和碳储量[D]. 南宁:广西大学,2017:13-14.
[21]李 况. 不同年龄桉树人工林生态系统碳氮储量分配特征[D]. 南宁:广西大学,2013:8-10.
[22]沈楚楚. 浙江省主要树种(组)生物量转换系数研究[D]. 临安:浙江农林大学,2013:15-17.
[23]钱国钦. 枫香人工林净生产量动态变化研究[J]. 江西农业大学学报,2000,22(3):399-404.
[24]张柳桦,齐锦秋,李婷婷,等. 林分密度对新津文峰山马尾松人工林林下物种多样性和生物量的影响[J]. 生态学报,2019,39(15):5709-5717.
猜你喜欢
今日农业(2022年13期)2022-09-15
中成药(2021年5期)2021-07-21
果树资源学报(2020年3期)2020-05-13
河北果树(2020年1期)2020-02-09
新疆农业科学(2019年11期)2019-12-31
天然产物研究与开发(2016年11期)2016-06-15
西南农业学报(2016年6期)2016-04-16
武夷学院学报(2015年3期)2015-07-18
小天使·四年级语数英综合(2015年6期)2015-07-17
新疆农垦科技(2014年7期)2014-02-28
