
铅胁迫对洋常春藤叶绿素荧光特性的影响
2020-11-12 12:46匡经舸李琬婷程小毛黄晓霞
浙江林业科技 2020年5期
匡经舸,李琬婷,李 响,程小毛,黄晓霞
(西南林业大学 园林园艺学院,国家林业与草原局西南风景园林工程技术研究中心,云南 昆明 650224)
在工业化、城市化的大背景下,铅(Pb)、镉(Cd)、铜(Cu)等重金属不仅污染环境,并严重威胁着动、植物的生存。重金属的生物毒性可改变植物细胞质膜的通透性,破坏其功能和结构,导致重金属离子更容易进入细胞,导致质膜上的酶活性絮乱,膜系统通透性加大,各种代谢活动无法正常进行[1]。为降低重金属胁迫伤害,植物启动自身抗氧化酶系统及渗透调节系统[2],积极利用自身调节机制在细胞内外形成螯合肽、金属硫蛋白等来减少重金属的毒害作用[3]。随着重金属离子浓度的增加,植物的光合作用电子传递受阻,叶绿体被破坏,植物叶绿素含量显著降低,进而导致叶片光合速率下降,同化物合成受阻,加速细胞衰老,严重时甚至会影响植物信息传导途径,导致蛋白功能降低,蛋白质合成趋于停顿,导致植株生长受阻[4-5]。叶绿素含量与叶绿素荧光参数是反映植物生理状况的重要指标,叶绿素吸收光能后,能迅速地发生氧化还原反应变化,将光能直接转化为化学能,推动有机物的合成。
洋常春藤Hedera helix为五加科Araliaceae 常春藤属Hedera多年生常绿缠绕类藤本植物,又名英国常春藤,既有观赏价值[6-7],也具药用功能[8],原产欧洲,目前国内外种植十分广泛,在中国主要分布于福建省和湖北省等光照充足、温暖湿润地区。洋常春藤叶形优美,攀岩能力强,可应用于庭院垂直绿化[9],阳台走廊绿化及地被应用[10-11],其叶片上的气孔能吸收尼古丁、甲醛等有害物质,可盆栽于室内,达到净化空气,绿化空间的目的[12-13]。此外,因含有皂苷类化合物,常春藤具有一定的药用价值。目前有关洋常春藤的研究主要集中于园林绿化、药用等[14-15]方面,而关于洋常春藤抗性生理方面的研究还未见报道。研究洋常春藤的耐性生理,对改善环境以及提高洋常春藤的综合利用价值具有深远意义。本试验以洋常春藤幼苗为材料,设计不同浓度的铅胁迫处理,通过测定洋常春藤叶片叶绿素含量和叶绿素荧光参数,旨在阐明洋常春藤光系统PSII 对不同铅胁迫的响应,以期了解洋常春藤对铅污染的适应能力,为重金属污染背景下的植物修复和保护提供理论依据。
1 材料与方法
1.1 试验材料和试验设计
于2016 年4 月中旬,在洋常春藤生长旺季进行试验。试验材料取自云南省昆明市西南林业大学老校区后山苗木基地(25°06′ N,102°76′ E),年平均气温为15℃,相对湿度为74%,土壤类型为红壤,质地为黏土,从中选取长势良好,生长状况基本一致的洋常春藤,将半木质化枝条修剪成10 cm 长的插穗进行扦插。扦插基质为红土∶珍珠岩∶腐殖土=1∶1∶2(体积比)。待扦插苗生长势稳定后,带回实验室。
于6 月将长势一致,生长状况良好的扦插苗放入装有MS 培养基的水培瓶中培养。本试验共设5 个不同浓度的Pb(CH3COO)2溶液处理组:对照(CK)浓度为0 μmol·L-1,轻度胁迫浓度为50 μmol·L-1(Pb1),中度胁迫浓度为200 μmol·L-1(Pb2)和400 μmol·L-1(Pb3),重度胁迫浓度为800 μmol·L-1(Pb4)。每个处理组10 瓶,每瓶一株。所有幼苗放入人工气候箱中培养,每天光照12 h(光强≥56.6 μmol·m-2·s-1),温度(白天/晚上)为25℃/20℃,湿度80%。供试材料铅胁迫处理时间为2016 年6 月15 日至8 月14 日,共60 d。处理结束后,于8月15 日将叶片完全展开的并处同一高度的叶片作为叶绿素含量和叶绿素荧光参数测定材料。
1.2 试验方法
1.2.1 叶绿素含量的测定 叶绿素含量测定:参照Inskeep and Bloom 的方法,称取0.3 g 植物新鲜叶片,剪碎,用4℃的二甲基甲酰胺在黑暗环境下浸提72 h,采用分光光度法于663.8 和646.8 nm 波长下进行比色。计算单位叶片质量的叶绿素a、叶绿素b 的含量,并计算总叶绿素(叶绿素a+b)以及叶绿素a/b。
1.2.2 叶绿素荧光参数的测定 将待测试验苗暗处理3 h 后,采用Imaging-PAM 便携式调制叶绿素荧光仪测定叶绿素荧光参数。利用叶夹夹住洋常春藤叶片,设置测量光强为0.5 μmol·m-2·s-1,饱和光脉冲为2 700 μmol·m-2·s-1,脉冲时间0.8 s,作用光照强度为76 μmol·m-2·s-1。初始荧光(F0),PSII 最大光化学效率(Fv/Fm),PSII 实际光合效率(Yield),非光化学猝灭系数(qN),光化学猝灭系数(qP)和PS 域非循环光合电子传递速率(ETR)等叶绿素荧光参数均由仪器自动给出。试验植株测试之前应经充分暗适应(20~ 30 min),测量时应避免叶片的主脉位于叶室打光的中部,每个处理至少5 个重复。
1.3 数据处理
试验数据采用Excel 和SPSS 22.0 软件进行数据处理并做相关性分析,采用Duncan’s 进行多重比较(α=0.05),采用Origin 9.1 软件作图。图表中数据为平均值±标准差。。
2 结果与分析
2.1 铅胁迫对洋常春藤叶绿素含量的影响
由表1 可知,与CK 相比,洋常春藤叶绿素a、叶绿素b 和总叶绿素含量在铅胁迫下均显著下降趋势(P<0.05),但不同浓度铅胁迫处理间的差异较小;与CK 相比,在Pb4胁迫处理下叶绿素b 含量下降了43%,比叶绿素a 含量下降(32%)更为明显,这使得叶绿素a/b 在Pb4胁迫处理下显著高于CK(P<0.05),而该指标在Pb1,Pb2和Pb3(50,200 和400 μmol·L-1)胁迫处理间并无显著差异。
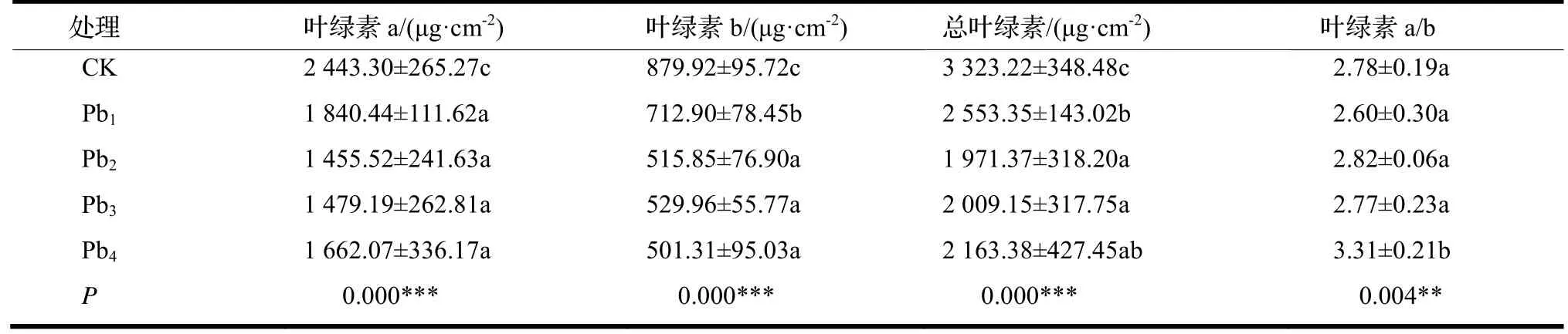
表1 铅胁迫对洋常春藤叶绿素含量的影响Table 1 Effect of different concentration of lead stress on the content of chlorophyll in H.helix
2.2 铅胁迫对洋常春藤F0 和Fv/Fm 的影响
由图1A 可以看出,洋常春藤幼苗受不同浓度铅胁迫后,随着铅胁迫浓度的增加其F0值整体上有所增加。与CK 相比,F0值在Pb1胁迫下无显著变化(P>0.05),在Pb2,Pb3和Pb4胁迫下显著上升(P<0.05),但3个处理组间无显著差异。此外,由图1B 可知,洋常春藤叶片的Fv/Fm在Pb1,Pb2,Pb3胁迫下有所上升,但与CK 之间均无显著差异(P>0.05),而在Pb4胁迫下其Fv/Fm显著下降(P<0.05),其Fv/Fm由CK 的0.72降至0.66。
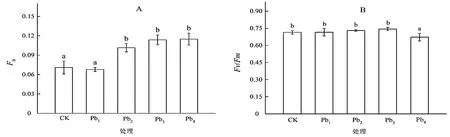
图1 铅胁迫对洋常春藤叶片F0 和Fv/Fm 的影响Figure 1 Effect of different concentration of lead stress on initial fluorescence and the maximum photochemical efficiency of PSII in H.helix
2.3 铅胁迫对洋常春藤叶片qN,qP 和Yield 和ETR 的影响
由表2 可看出,与CK 相比,铅胁迫对洋常春藤叶片的qN,qP,Yield以及ETR均有不同程度的影响。在轻度铅胁迫下,植株的qN,qP,Yield以及ETR与CK 相比均无显著差异;而在中度和重度铅胁迫下,植株的Yield,qP和ETR,随着铅浓度的增加而呈下降趋势,qN却呈上升的趋势,在重度铅胁迫时高出CK 15.7%。
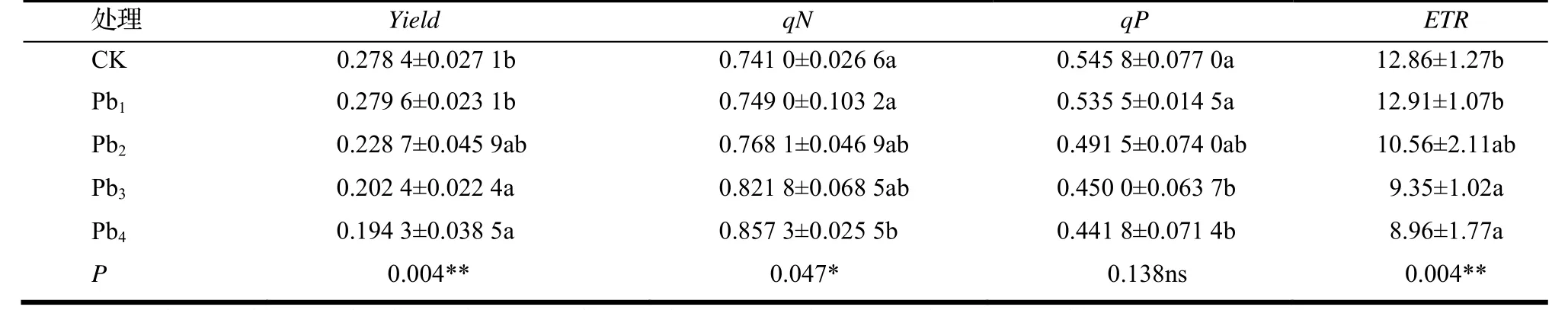
表2 铅胁迫对洋常春藤叶片Yield,qN,qP 和ETR 的影响Table 2 Effect of different concentration of lead stress on Yield, qN, qP and ETR of H.helix
2.4 铅胁迫下洋常春藤叶绿素指标与叶绿素荧光参数相关性分析
由表3 可知,在铅胁迫下,洋常春藤叶片的叶绿素a 和叶绿素b 极显著正相关(r=0.949*),总叶绿素与叶绿素a 和叶绿素b 均极显著正相关(r=0.996**,r=0.974**);F0与Fv/Fm,Yield,qN,qP及ETR均无显著相关性。Yield与qP和ETR均极显著正相关(r=0.992**,r=1.000**);qP与ETR极显著正相关(r=0.992**)。
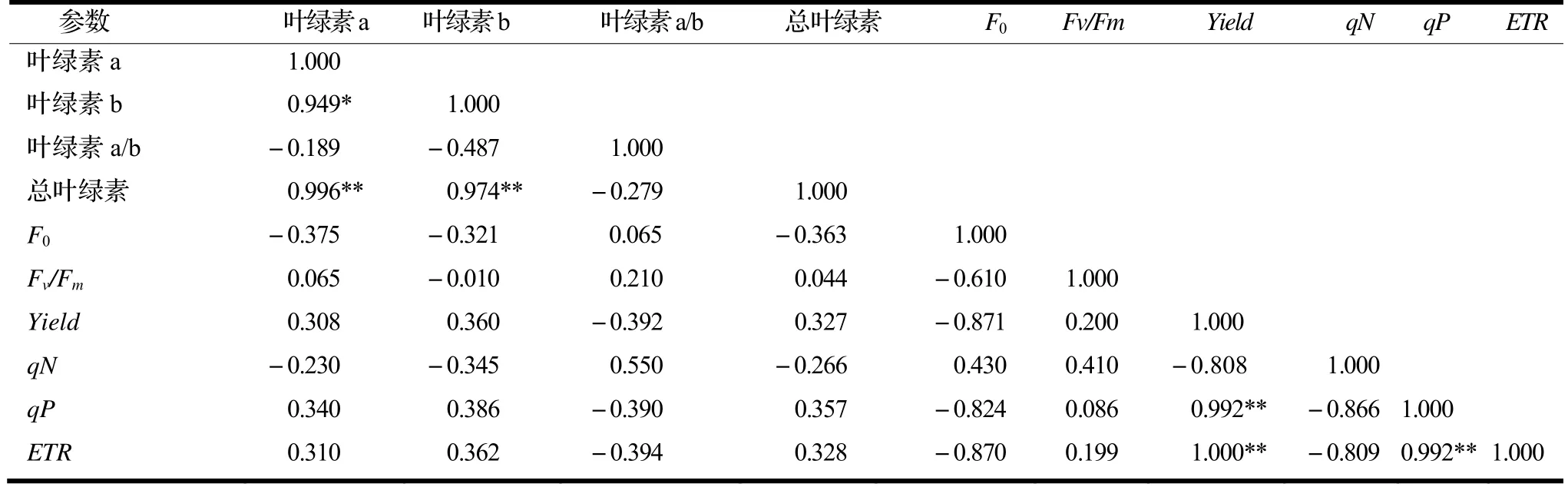
表3 铅胁迫下洋常春藤叶绿素荧光参数与叶绿素指标的相关性分析Table 3 Correlation between chlorophyll fluorescence parameters and chlorophyll parameters of H. helix treated by different concentration of lead
3 结论与讨论
叶绿素存在于植物叶绿体片层膜系统中,是植物进行光合作用的重要组成部分。叶绿素吸收光子后,就进入不稳定的、高能状态的激发态,由激发态能量推动光化学反应进行。重金属会使植物膜系统破坏,导致位于膜上的酶活性紊乱,同时影响叶绿素的合成,导致植物的光能利用率降低[16-18]。本研究中在铅胁迫下,洋常春藤的叶绿素含量显著低于对照的(P<0.05),这可能是铅胁迫造成的离子毒害导致叶绿体受损,使合成叶绿素的相关性酶失活或变性,叶绿素合成受阻,降解加快,最终导致洋常春藤叶片的叶绿素含量显著降低,说明洋常春藤叶绿素对铅胁迫较为敏感。此外,叶绿素a/b 在Pb4(800 μmol·L-1)胁迫下显著上升(P<0.05),说明洋常春藤叶片的叶绿素b 比叶绿素a 对铅胁迫更为敏感,出现这一情况可能是由于高浓度铅胁迫下植物光合机能调节器官受损,减缓了叶绿素的合成,为维持其基本光合作用而进行一系列自我保护措施,这一结果与李西[19-20]等的研究结果相类似,认为铅胁迫可显著降低植株叶片中的叶绿素含量。叶绿素荧光参数的变化反映了光系统对光能的吸收、传递与耗散,反映植物受胁迫的状况[21]。当叶绿体受到重金属胁迫时,金属离子抑制了Hill 反应,引起正电荷在水氧化系统水平上的积累,导致了围绕PSII 的循环电子流动,进而使PSII 和光合电子传递活性受到抑制[22]。
在本研究中,洋常春藤叶片初始荧光(F0)值在Pb2,Pb3和Pb4(200,400 和800 μmol·L-1)胁迫下均呈显著上升(P<0.05),且在重度铅胁迫下达到最大值,与此同时,植株叶片的PSII 最大光化学效率(Fv/Fm)显著降低(P<0.05),达到最小值。这说明洋常春藤叶片在Pb4时其PSII 反应中心的破坏或者可逆失活引起F0水平的增加,导致PSII 放氧复合体脱离,反应中心失活,原初反应的光能转换效率下降,最终导致光合机构受到损伤。在Pb1,Pb2,Pb3(50,200,400 μmol·L-1)铅胁迫下,其PSIIFv/Fm之间并没有显著差异,说明该浓度铅胁迫对洋常春藤PSII 的活性影响不大,说明洋常春藤对轻、中度铅胁迫具有较好的耐受性。前人[23-25]研究发现在铅胁迫初期不同植物可通过调节自身的应激性来增加PSII 活性去抵御重金属毒害,但由于PSII 活性中心受损,使得植株受到严重毒害,反应中心失活使得被动保护机制逐渐失去作用。本研究与这些研究结果存在一些不同之处,说明不同植物对重金属胁迫的荧光特性响应存在一定差异。光化学猝灭系数(qP)反映了原初反应中原初电子受体的还原情况和PSII 开放的程度;非光化学猝灭系数(qN)值反映PSII 反应中心对天线色素吸收过量光能后的以热能散发掉的部分,可反映光和机构的受损程度;光合电子传递速率了(ETR)反映了植物光合作用的表观电子传递;PSII 实际光合效率(Yield)是反映植物光合能力的重要指标之一,代表PSII 反应中心进行光化学反应的效率[26]。本研究结果与刘涛等[27]的研究结果相似,洋常春藤叶片在Pb1(50 μmol·L-1)铅胁迫下,qN、qP,Yield以及ETR与对照相比均无显著差异,表明此胁迫浓度下植株的电子传递活性较高,受抑制程度较轻;随铅浓度的增加,除qN显著上升外,Yield,qP和ETR均呈下降趋势,则说明铅胁迫抑制洋常春藤叶片PSII 反应中心的开放比例,导致PSII 光能转化效率下降,光合电子传递效率下降,光抑制加剧,以热或者其他形式耗散的光能增加,为避免光合机构损伤,表现出洋常春藤的自我保护机制。在铅胁迫处理下洋常春藤叶片的叶绿素a 和叶绿素b 呈极显著正相关,叶绿素(a+b)与叶绿素a 和叶绿素b 之间均呈极显著正相关;洋常春藤叶片的qP,ETR与Yield均呈极显著正相关,qP与ETR呈极显著正相关。这表明在铅胁迫处理下洋常春藤叶片的ETR受PSII 光能转换效率影响较大,其转变过程及机理尚不清楚,有待进一步试验研究。从本试验结果来看,在Pb1下,洋常春藤叶片的F0,Fv/Fm,qN,qP,Yield以及ETR与对照相比均无显著差异,F0上升程度和Fv/Fm的下降程度均在Pb2,Pb3和Pb4下逐渐增大,其他荧光参数Yield,qP 和ETR下降程度亦是如此。
综上所述,Pb1(50 μmol·L-1)铅胁迫对洋常春藤幼苗叶绿素含量及叶绿素荧光参数无显著影响,而Pb2和Pb3(200,400 μmol·L-1)铅胁迫对其有一定的影响,但光系统未受明显损伤,说明该植物对铅胁迫具有较好的耐受性。建议今后可对洋常春藤的铅耐受性及铅吸附富集能力等进行更深一步的研究,以期作为铅污染地区的生态修复地被物种进行推广应用。
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13
天津医科大学学报(2021年3期)2021-07-21
阅读(科学探秘)(2020年8期)2020-11-06
绿色科技(2019年2期)2019-05-21
中国果业信息(2019年1期)2019-01-05
中央民族大学学报(自然科学版)(2018年3期)2018-11-09
科学之友(2016年19期)2016-02-07
食品工业科技(2014年6期)2014-05-10
女士(2012年10期)2012-05-08
