
滤除自然光中UV-B辐射成分对杜仲光合和次级代谢产物的影响
2021-01-04 07:39李德文杜丹丹季倩如郭晓瑞于雪莹唐中华
经济林研究 2020年4期
李德文,杜丹丹,季倩如,郭晓瑞,刘 英,于雪莹,唐中华
(东北林业大学 a.森林植物生态学教育部重点实验室;b.化学化工与资源利用学院,黑龙江 哈尔滨 150040)
紫外线-B(UV-B)辐射是太阳辐射光谱中一个非常重要的组成部分,极易受多种因素的影响[1]。近年来,一些人为污染物,如氯氟化碳化合物(CFC)和其他拮抗剂等,严重破坏了大气平流层中的臭氧(O3),导致地球表面UV-B(波长280 ~315 nm)辐射强度不断增加。有研究结果表明,大气平流层中臭氧每减少1%,到达地球表面的太阳紫外线辐射就会增加2%[2]。尽管到达地球表面的UV-B 辐射强度大约仅占太阳短波辐射的1.5%,但由于其能被一些核酸、蛋白质等生物大分子有效吸收,地球生态系统中的生物尤其是高等植物的生长发育会遭到严重干扰。自然条件下,环境UV-B 辐射强度增加会对植物的生长、形态特征、生理生化过程、基因表达、抗氧化酶活性及次级代谢产物含量等产生严重影响[3]。今后较长一段时间内,陆地上的生物将继续受到太阳UV-B 辐射的影响,因此,揭示UV-B 辐射对生态系统中动植物的影响,并制订相关的有效举措,将会成为研究者关注的焦点。
光合作用是植物重要的生理代谢反应之一,也是植物形态建成的基础。光合机构是UV-B 辐射的中心靶点。UV-B 对光合作用的影响主要包括光系统Ⅱ(PS Ⅱ)反应中心失活、类囊体膜系统损伤及光合色素含量下降等[4]。Jansen 等[5]认为,植物对UV-B 辐射的响应机制是胁迫、修复和驯化三者之间综合平衡的结果,在光修复过程中,可见光和UV-A(315 ~400 nm)能激活光复活酶,并分解经紫外线照射而形成的嘧啶二聚体,使UV-B 辐射造成的DNA 和蛋白质等大分子物质损伤得以修复。驯化是指给予植物一定的非致死的胁迫处理,植物体会产生一些可遗传的行之有效的防御策略来应对随之而来的更恶劣的逆境,与未经驯化的植物相比其抗逆性更强,这种现象像是植物具有了对胁迫的“记忆力”或者印记。驯化过程中植物体可产生一些有益的生物化学物质,如何凤等[6]发现经过低温驯化后杜仲花芽抗氧化物质和酶活性均显著增加。不同作物对UV-B 辐射的响应机制有明显差异,有些物种能被UV-B 辐射刺激而产生抗性,有些则非常敏感。
杜仲Eucommia ulmoides,别名思仙、思仲、玉丝皮棉等,是中国珍稀濒危孑遗树种,具有重要的药用价值和经济用途,是中国特有的十分重要的战略资源树种[7]。野生杜仲在我国亚热带至温带的27 个省(区、市)均有栽培,由于其适应性强,现已被广泛引种到亚洲、欧洲和北美洲等地区[8]。但杜仲栽培生境差异较大,各地引种杜仲长势明显不同,尤其高UV-B 辐射地区杜仲生长受到抑制,光合速率和部分紫外吸收物质(如桃叶珊瑚苷、京尼平苷酸和绿原酸)的含量等显著降低[9], 因此对杜仲培育技术及栽培生理进行深入研究具有重要的现实意义。本研究中以药用植物杜仲为材料,在自然光辐射背景下,通过滤除UV-B 辐射,研究UV-B 辐射对杜仲光合系统的影响,并结合光合色素和次级代谢产物的变化,探讨了杜仲对自然环境中UV-B 组分的适应性,以期为杜仲的定向生态培育提供参考。
1 材料与方法
1.1 研究区概况
试验样地位于哈尔滨市东北林业大学森林植物生态学教育部重点实验室温室内。哈尔滨市地处中国东北平原东北部地区,黑龙江省南部,位于东经125°42′~130°10′、北纬44°04′~46°40′,属中温带大陆性季风气候,冬长夏短,四季分明,全年平均降水量569.1 mm,年平均温度3.6 ℃,集中降水期为每年7—8月,集中降雪期为每年11—1月(次年)。
1.2 植物材料和栽培条件
试验材料为3年生杜仲盆栽实生苗,常规水肥管理。栽培土壤有机碳、氮、磷含量分别为(571.11± 38.48)、(24.92±0.98)、(2.35±0.25) g/kg。
1.3 试验方法
1.3.1 自然光中UV-B 辐射成分的滤除
滤除自然光谱中UV-B 辐射成分的模拟试验主要参照Flint 等[10]的方法,并略有改进。可移动的样方框架采用三角铝材制作,顶部为长方形,长220 cm,宽120 cm,高180 cm。在框架上端覆盖Mylar 型Luminar 薄膜(Toray,日本),厚度 0.08 mm,可滤除自然光谱中绝大部分UV-B 成分。在膜下部留80 cm 高度,以便风和雨水通透,在覆膜顶部留出直径为0.6 cm、孔距为20 cm×20 cm 的小孔。分别覆盖1 层膜(T1 处理组)和2 层膜(T2处理组)模拟不同程度UV-B 滤减,以自然环境处理(不减弱UV-B)作为对照(CK)。每处理20盆,各设3 个重复。各处理的UV-B 辐射剂量滤除率如图1 所示,各处理的UV-B 平均滤除率分别为51.64%和80.68%。2017年6月23日开始处理,处理时间为每天9:00—16:00,根据天气情况等略有调节,并分别于处理进行22 和41 d 时取样。
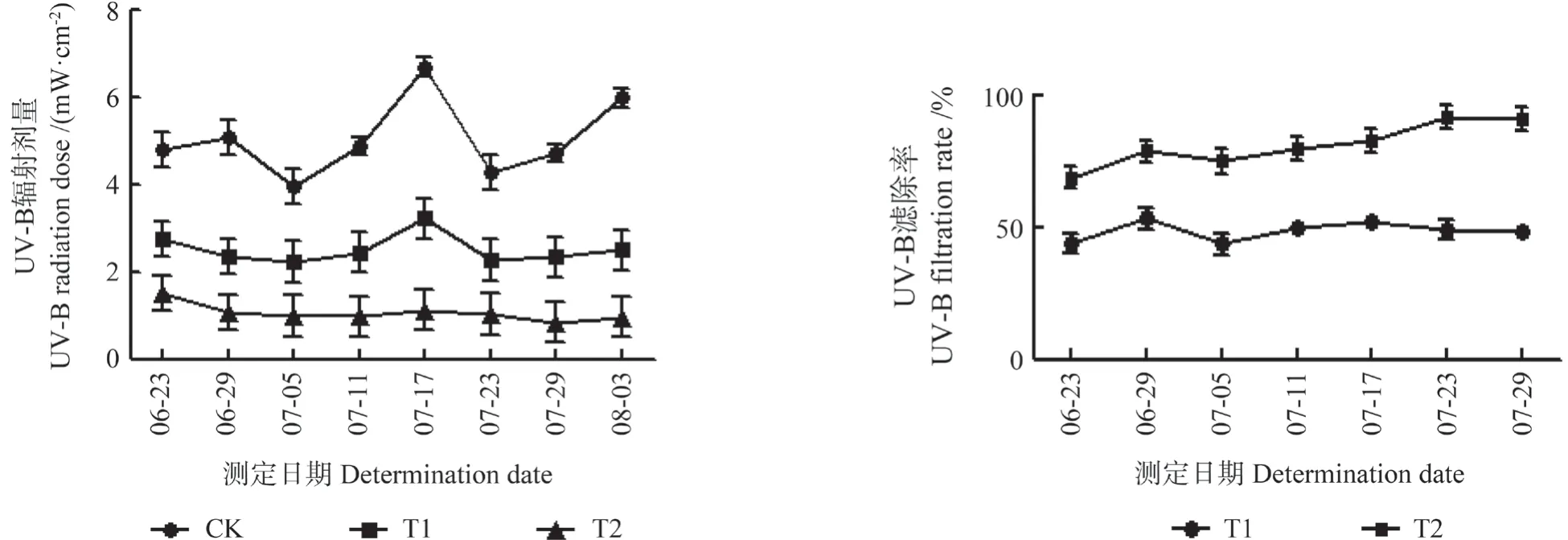
图1 各处理的UV-B 辐射剂量和UV-B 滤除率Fig.1 UV-B radiation dose of each treatment and UV-B filtering proportion of T1 and T2 treatment groups
1.3.2 光合参数和光合色素含量的测定
于晴天9:30—11:00,选取植株已完全展开的生长状况、叶龄和叶位一致的叶片,采用Li-6400型便携式光合仪测定供试植株净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。按照董晓颖等[11]的方法计算水分利用效率(EWU)、叶片羧化效率(EC)和气孔限制值(Ls)。

式中:Ca表示大气CO2浓度,为400 μmol/mol。
参考Cen 等[12]的方法,使用便携式PAM-2500叶绿素荧光仪测定叶绿素荧光参数,并根据Yao 等[13]的方法计算PS Ⅱ最大光合效率(Fv/Fm)、PS Ⅱ实际光合效率(ΦⅡ)和电子传递速率(RET)。
参考魏晓雪[14]的方法测定光合色素含量。取0.05 g 叶片鲜样,剪碎,置于试管中,加入5 mL二甲基亚砜(DMSO),黑暗条件下60 ℃水浴反应,直至样品组织完全变白。以DMSO 为空白,使用Shimadzu UV-2550 紫外可见分光光度计(日本)分别测定提取液在480、649、665 nm 处的吸光值,并计算叶绿素a、叶绿素b、总叶绿素、类胡萝卜素含量及叶绿素a 和叶绿素b 含量比值。
1.3.3 次级代谢产物含量的测定
杜仲叶片次级代谢产物的提取参考Dong[15]等的方法,取1 g 叶片鲜样置于试管中,加入8 mL色谱级甲醇于Xin-yi48 高通量组织研磨仪(宁波新艺超声设备有限公司)中研磨震荡(60 Hz, 2 min),使用超声波清洗器超声处理1 h(功率100 W,频率40 kHz,40 ℃),震荡混匀后使用低温高速离心机离心10 min(8 000 r/min)。抽取上清液,在40 ℃下利用真空旋转蒸发仪浓缩挥干,挥干后再次用甲醇复溶至2 mL,离心10 min,提取上清,密封,-20 ℃保存备用。使用0.45 μm微孔滤膜过滤,随后采用液相色谱-质谱联用仪进行色谱分析,C18 色谱柱(2.1 mm×100 mm, 1.6 μm),柱温25 ℃,流动相为溶剂A(62%水)与溶剂B(38%甲醇),流速为0.25 mL/min,进样量5 μL。桃叶珊瑚苷、绿原酸、京尼平苷酸、京尼平苷和京尼平标准品均购于Sigma 公司,纯度大于或等于98%。
1.4 数据处理
使用SPSS 19.0 和Excel 2010 软件进行数据分析,采用单因素方差分析方法(One-way AVONA)检验各处理组间差异的显著性,采用相关分析法进行各指标间相关性的分析,图表中数值为“平均值±标准误”。
2 结果与分析
2.1 滤除UV-B 辐射处理对杜仲叶片光合参数的影响
滤除自然光中UV-B 辐射成分对杜仲光合参数的影响如图2 所示。由图2 可以看出,处理22 d 时,杜仲叶片Pn和Gs随滤除率的增加显著降低,Ci和Tr在T1 处理组无差异,T2 处理组显著降低(P< 0.05)。处理41 d 时叶片Pn、Gs和Tr显著增加,且T2 处理组增加最大,与对照相比分别增加了61.9%、73.8%和70.1%;Ci在T1 处理组有显著下降趋势,在T2 处理组与对照无显著差异(P> 0.05)。滤除太阳辐射光谱中UV-B 成分后,杜仲叶片Fv/Fm在2次取样时与对照相比均无显著差异;ΦⅡ在2 次取样时变化趋势相反,第1 次显著下降,第2 次显著上升;RET在各处理组显著高于对照 (P<0.05),但T1、T2 处理间无显著差异。说明短期滤除自然光谱中UV-B 成分对杜仲叶片的光合机构产生负效应,随着滤除时间的延长,能引起叶片稳态光化学效率的增加,并对叶片光合速率产生正效应。
滤除自然光中UV-B 辐射成分后杜仲叶片EWU、EC及Ls见表1。由表1 可知,与对照相比,处理22 d 时,杜仲叶片EWU显著降低,T1 处理组下降幅度最大,较CK 下降了40%,EC在各处理间无显著差异,Ls在T2 处理组显著增加,T1 与CK 间无显著差异(P>0.05)。处理41 d 时,杜仲叶片EWU值与CK 相比在T1 处理组显著上升,在T2 处理组显著下降,EC随UV-B 滤除率的增加呈上升趋势,Ls在T2 处理组显著降低(P<0.05)。
2.2 滤除UV-B 辐射处理对杜仲叶片光合色素的影响
滤除自然光中UV-B 辐射成分对杜仲叶片光合色素的影响如图3 所示。由图3 可以看出,2 次取样时间的杜仲叶片叶绿素a、叶绿素b 和总叶绿素含量有一致的变化趋势,与CK 相比均显著增加,且处理41 d 时叶绿素含量显著大于处理21 d。滤除UV-B 辐射后,2 次取样时间的杜仲叶片类胡萝卜素含量均表现为降低的趋势,叶绿素a 和叶绿素b含量比值在各处理间均无显著差异(P>0.05),但处理41 d 时其比值显著大于处理22 d。表明长期滤除UV-B 辐射处理中杜仲叶片光合色素的含量比短期滤除处理高。
2.3 滤除UV-B 辐射处理对杜仲叶片次级代谢产物含量的影响
为了解滤除自然环境中UV-B 辐射成分对杜仲叶片主要药用活性物质的影响,通过UPLC-Q/TOF-MS 分析收集样品中目标化合物的代谢变化,并根据相应的标准参考化合物的质谱信息和保留时间来鉴定样品中的目标化合物。滤除自然光中UV-B 辐射成分对杜仲次级代谢产物含量的影响如图4 所示。由图4 可以看出,滤除自然光谱中的UV-B 辐射后,处理22 d 时杜仲叶片次级代谢产物中京尼平、京尼平苷酸和桃叶珊瑚苷含量均显著增加,京尼平苷含量呈下降趋势。绿原酸含量在T1 处理组增加,在T2 处理组与对照无显著差异。与对照相比,T1 处理组和T2 处理组的京尼平含量分别增加了43.5%和39.4%,京尼平苷酸含量分别增加了18.8%和6.7%。桃叶珊瑚苷含量在T1处理组与对照无显著差异,在T2 处理组较对照增加了58.3%。京尼平苷含量在T1 处理组和T2 处理组分别下降了21.7%和51.1%(P<0.05)。处理41 d 时,叶片中5 种次级代谢物质含量均显著增加(P<0.05)。与对照相比,T1 处理组和T2处理组的京尼平含量显著增加了40.2%和30.0%,京尼平苷含量分别增加了33.3%和8.8%,京尼平苷酸含量分别增加了57.4%和23.2%,桃叶珊瑚苷含量分别增加了60.7%和65.8%,绿原酸含量分别增加了21.1%和23.7%。对比2 次取样叶片的次级代谢产物含量发现,处理41 d 时各处理组除京尼平苷和京尼平苷酸含量显著低于处理22 d外,其他次级代谢产物含量均显著高于处理22 d。

图2 滤除自然光中UV-B 辐射成分对杜仲叶片光合参数的影响Fig.2 The effect of filtering out UV-B radiation components in natural light on photosynthetic parameters of E.ulmoides leaves

表1 滤除自然光中UV-B 辐射成分后杜仲叶片水分利用效率、叶片羧化效率和气孔限制值†Table 1 The leaf water use efficiency, leaf carboxylation efficiency and stomatal limitation value of E.ulmoides after UV-B radiation was filtered out from natural light
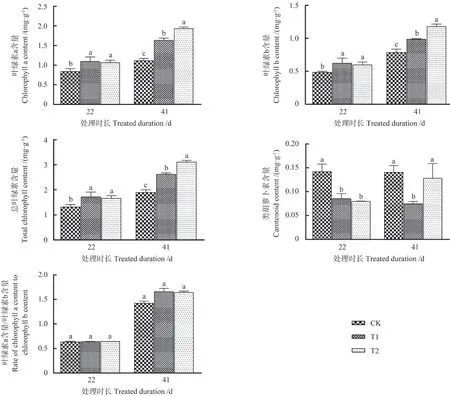
图3 滤除自然光中UV-B 辐射成分对杜仲叶片光合色素的影响Fig.3 The effect of filtering out UV-B radiation components in natural light on photosynthetic pigments of E.ulmoides leaves
3 结论与讨论
综上所述,长期滤除太阳辐射光谱中UV-B成分后,杜仲叶片光合速率和稳态光化学效率增加,光合色素含量提高,处理41 d 时次级代谢产物除京尼平苷酸含量显著低于处理22 d 外,绿原酸、京尼平、京尼平苷和桃叶珊瑚苷含量均显著高于处理22 d。结果表明,在长期滤除自然光谱中UV-B 成分处理中,杜仲能正常从根系吸收水分,提高蒸腾,保证植株正常发育。另外,长期滤除UV-B 成分后,杜仲叶表皮层中积累较多的次级代谢产物,以充分抵御环境中UV-B 辐射波动引起的伤害。可见,杜仲更大程度上通过叶片结构和光合作用的变化来适应UV-B 辐射,自然环境中UV-B 成分依然是限制药用植物杜仲叶片光化学效率的因素之一。
由于植物在环境中无法移动位置,在其生长周期中不可避免地会受到太阳紫外辐射这个自然环境因子的影响。UV-B 辐射对植物生长发育、光合生理过程及次级代谢的影响一直是国内外学者关注的重点[16]。植物光合作用和次级代谢产物的改变影响地球生物圈中动物、微生物及人类的生长发育,决定着生态系统对UV-B 辐射的响应程度。采用滤光薄膜滤除自然环境中大部分UV-B 辐射成分的方法,是研究UV-B 的植物生物学效应的重要途径之一。
光合作用是一个极为复杂的生理过程,包括一系列光化学步骤和物质转变过程,其对UV-B 辐射的敏感度在不同作物间各有差异。根据前人研究结果,UV-B 辐射增强导致光合作用受阻(主要包括叶片气孔或非气孔因素)[17]、叶绿体结构被破坏并使叶绿素分解[18]、希尔反应活性和RUBP再生率下降[19]等。王丹等[20]发现UV-B 辐射下紫花苜蓿幼苗Pn、Gs和Tr显著降低,认为UV-B 辐射使叶绿体结构中光敏色素B 被破坏,从而导致光合速率下降。本研究结果表明,与对照组相比,滤除UV-B 辐射22 d 时杜仲叶片Pn、Tr和Gs显著下降,而滤除41 d 时叶片Pn、Tr和Gs显著增加,Ci在T1 处理组显著降低,说明自然环境中UV-B辐射对杜仲叶片的Pn具有一定的限制作用。经过短期滤除(22 d),杜仲叶片可能产生了胁迫记忆,随着滤除时间的延长,与短期滤除相比其耐受能力更强。拟南芥和玉米植株经过多次脱水、复水处理后,会表现出比第1 次处理更高的水分利用率[21]。在本试验中,对照组杜仲叶片Ci升高且Gs值降低,因此影响Pn的主要原因是非气孔限制因素。此外,EWU的降低表明杜仲在滤除UV-B辐射后积累有机物所需水分减少,足以保证植株正常生长发育。EC是表征植物光合能力的重要参数,对光合速率起着决定作用[22],本研究中发现,长期滤除UV-B 后杜仲叶片EC显著增加,说明UV-B 辐射是影响杜仲叶片Pn的原因。据此推测,长期滤除环境中UV-B 辐射对杜仲叶片光合速率有正效应。
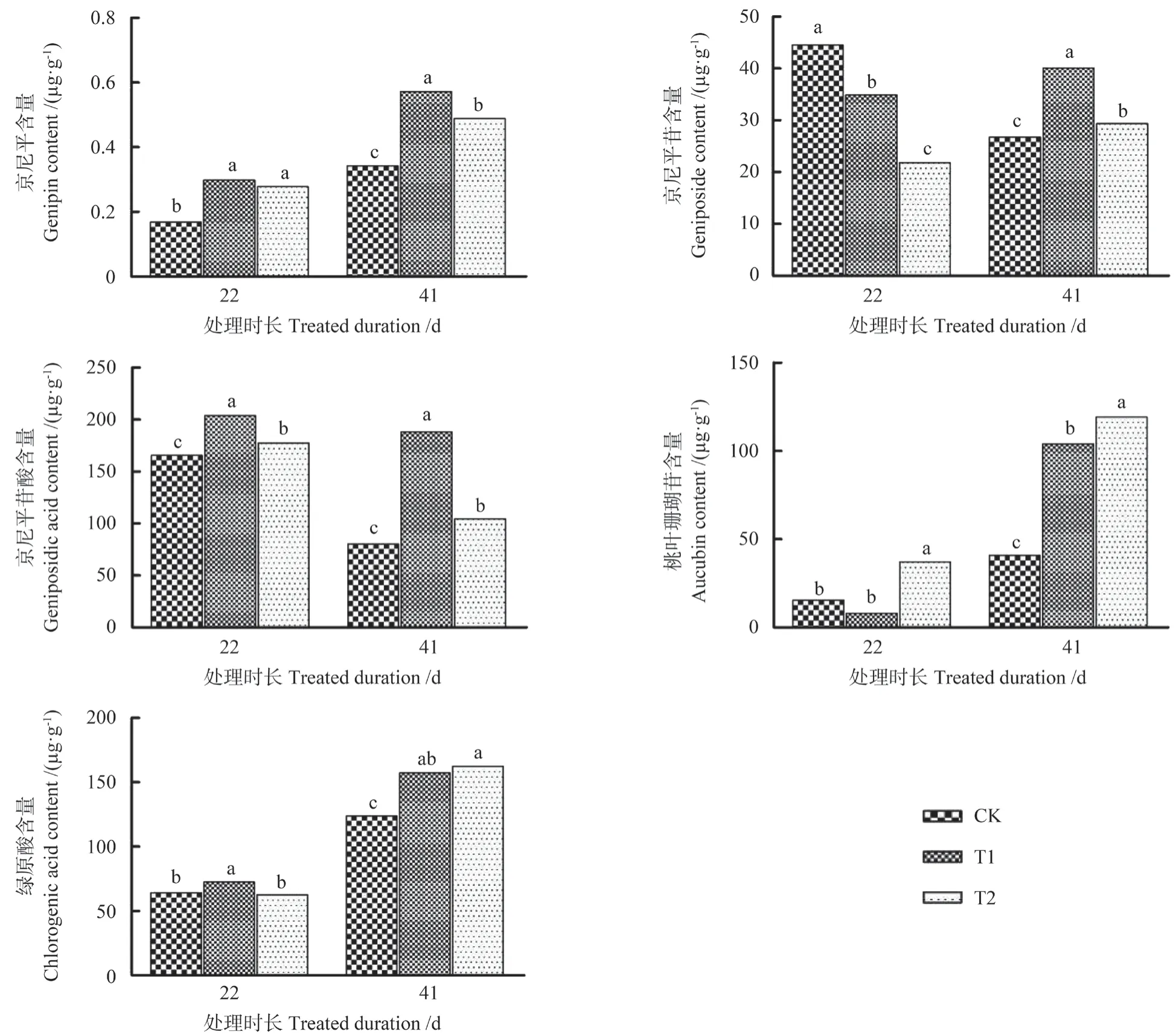
图4 滤除自然光中UV-B 辐射成分对杜仲叶片次级代谢产物含量的影响Fig.4 The effect of filtering out UV-B radiation components in natural light on secondary metabolite contents of E.ulmoides leaves
叶绿素荧光是光合作用与外界环境间的探针,利用该指标可以探究植物光合系统对环境UV-B 变化的响应机制[23]。PS Ⅱ是UV-B 对光合作用损伤的主要靶子,Fv/Fm被用于估测植物叶片PS Ⅱ反应中心的最大光能转化效率,ΦⅡ代表PS Ⅱ反应中心电荷分离的实际化光化学效率[24]。本研究中发现,滤除UV-B 辐射后杜仲叶片Fv/Fm与对照无显著差异,RET在各处理组均显著高于对照,ΦⅡ在滤除41 d 时显著增加。这与前人的研究结果一致,丁伟等[4]经研究发现高山植物美丽风毛菊在滤除UV-B 辐射后Fv/Fm和ΦⅡ均显著增加,认为自然环境中的UV-B 成分抑制了Fv/Fm的恢复,限制了PS Ⅱ中原初电子受体QA的氧化速率,也制约着PS Ⅱ反应中心的电子传递和光化学量子效率的恢复。因此,导致杜仲光合速率降低的原因是UV-B 影响了PS Ⅱ系统光化学能的转换,破坏了电子传递体,从而降低了光能转换效率,自然环境中UV-B 成分是限制药用植物杜仲叶片光化学效率的重要因素。
光合色素参与光合作用中光能的吸收、传递和转化,其含量在一定程度上反映植物利用光能和制造有机物的能力,制约着植物生长发育,是植物干物质积累和产量的基础[25]。通常认为,UV-B 辐射会使植物叶绿体超微结构遭到破坏,表现为叶绿体结构变形、膜系统损伤、类囊体片层和基粒排列稀疏紊乱[26],叶绿素水平对UV-B 的响应依赖于植物的发育过程和环境条件,强UV-B辐射下光合色素含量的降低可能缘于其对光合色素合成过程的限制或其光抑制及光损伤作用[27]。本研究结果也表明,滤除UV-B 辐射成分后,杜仲叶片的叶绿素a、叶绿素b 和总叶绿素含量均表现为增加的趋势,能免于强UV-B 辐射的损伤;对照组中UV-B 辐射成分降低了叶片叶绿素的含量,表明UV-B 辐射破坏了杜仲叶绿体结构,抑制光合蛋白复合物和细胞器的形成,导致叶绿素含量下降。类胡萝卜素分子能与生物膜结合,吸收紫外光,是一种重要的抗氧化剂,在电子的吸收和传递过程中起重要作用,保护细胞、叶绿体和叶绿素免受伤害[28]。本研究结果表明,滤除UV-B 辐射后杜仲叶片类胡萝卜素含量呈降低趋势。说明随UV-B 滤除率的增加,植物叶片受到紫外辐射的强度减弱,因此叶绿素含量增加,类胡萝卜素含量减少,也验证了自然环境中UV-B 辐射成分对叶片光合色素合成过程的抑制和膜脂过氧化作用是光合速率降低的本质原因[29]。
植物对UV-B 辐射的一个重要防御策略是通过在叶片表皮层中形成、控制和诱导胁迫反应相关植物化学物质来提高自身的抗逆性。与抗性相关的植物化学物质中次级代谢产物是一大类普遍存在的植物特异性化合物,对植物生长、发育和缓解胁迫至关重要。这些次级代谢产物对中波紫外线具有强烈的吸收作用,允许可见光通过,不影响光合作用进行,能形成一道天然屏障,是植物的紫外线保护剂[30]。李锦馨等[31]经研究发现,植物次级代谢产物的合成与环境UV-B 辐射强度呈正相关,即叶片照射的UV-B 强度越高,植株生成的次级代谢产物越多。付金颖[32]的研究结果表明,短期增强UV-B 辐射能够增加杜仲叶片绿原酸含量,降低桃叶珊瑚苷、京尼平苷酸和京尼平苷含量。马春晖[33]经研究发现,增强UV-B 辐射后祁菊花序中绿原酸和黄酮类物质含量显著提高。本研究结果表明,长时间滤除环境UV-B 成分后,杜仲叶片中仅京尼平苷酸含量降低,绿原酸、京尼平、京尼平苷和桃叶珊瑚苷含量呈增加趋势,表明长期生活在自然环境下不利于杜仲叶表皮层中次级代谢产物的积累,滤除UV-B 成分后次级代谢产物的增加足以抵御环境UV-B 辐射波动的影响。如杨超[34]经研究发现,UV-B 胁迫影响了南方红豆杉体内紫杉醇的含量,而紫杉醇可以显著降低植株内过氧化氢和超氧阴离子等自由基的含量,证实了紫杉醇等生物碱的合成有利于植物对环境UV-B辐射的适应。因此,长期的UV-B 滤除对杜仲产生了驯化作用,驯化是提高药用植物药效成分含量的关键步骤,有些药用植物在轻度胁迫下反而能产生更多的次级代谢化合物,以提高其适应性。
本研究中仅对温室盆栽杜仲实生苗进行试验处理,而温度、水分和光照时间等不可控因素均会影响杜仲叶片中主要药用活性物质的含量。为了提高杜仲叶产量和品质,下一步将研究在大田推广中采用滤除自然光谱中UV-B 辐射成分等措施对杜仲的生长特征、光合作用及生理代谢特征的影响。
猜你喜欢
娃娃乐园·综合智能(2022年9期)2022-08-16
科学大众(2021年9期)2021-07-16
阅读(科学探秘)(2020年8期)2020-11-06
绿色科技(2019年2期)2019-05-21
中国果业信息(2019年1期)2019-01-05
中成药(2018年8期)2018-08-29
中国调味品(2017年2期)2017-03-20
陕西画报(2016年1期)2016-12-01
小雪花·小学生快乐作文(2015年9期)2015-09-29
小雪花·小学生快乐作文(2015年9期)2015-09-29
