
马铃薯块茎蛾对不同品种马铃薯块茎的适应性分析
2021-03-09 00:58张梦迪闫俊杰高玉林
中国农业科学 2021年3期
张梦迪,闫俊杰,高玉林,2
马铃薯块茎蛾对不同品种马铃薯块茎的适应性分析
张梦迪1,闫俊杰1,高玉林1,2
1中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室,北京 100193;2中国农业科学院国家薯类作物研究中心,北京 100081
【】马铃薯块茎蛾()是马铃薯上的重要害虫,严重影响马铃薯的产量和品质。明确马铃薯块茎蛾对不同品种块茎的适应性,为马铃薯抗性品种筛选以及制定有效的害虫综合治理策略提供依据。以目前生产上商业化推广的4个马铃薯品种(夏波蒂、龙薯4号、龙薯12号、克新17号)块茎为试验材料,采用年龄-阶段特定两性生命表,测定不同品种块茎上马铃薯块茎蛾的发育历期和种群参数,以评价马铃薯块茎蛾在不同品种上的适应性。取食不同品种块茎的马铃薯块茎蛾幼虫期、蛹期和雌成虫寿命存在显著差异。其中取食克新17号的马铃薯块茎蛾幼虫期(15.18 d)和蛹期(8.10 d)相较于取食其他3个品种(夏波蒂、龙薯4号、龙薯12号)长,而雌成虫寿命(8.46 d)较短。另外,取食同种品种块茎的马铃薯块茎蛾雄成虫寿命普遍高于雌成虫。取食不同品种块茎的马铃薯块茎蛾单雌产卵量存在显著差异,取食夏波蒂(235.06粒)和龙薯4号(254.48粒)的马铃薯块茎蛾单雌产卵量显著多于取食克新17号(165.71粒)。取食不同品种块茎的马铃薯块茎蛾内禀增长率()、周限增长率()、净增殖率(R)和平均世代时间()等种群参数存在极显著差异。其中取食克新17号的马铃薯块茎蛾的、、R值(=0.108,=1.125,R=26.513)均显著低于其余3个品种,值(30.28)显著高于其余3个品种。在幼虫选择性取食试验中,1龄马铃薯块茎蛾幼虫尤为偏好龙薯4号和夏波蒂,对克新17号选择性最低, 并随着龄期的增长,选择性逐渐降低。马铃薯块茎蛾对不同品种薯块的适应性不同,在夏波蒂和龙薯4号上适应性较好,龙薯12号次之,克新17号种群建立水平最低。在马铃薯块茎蛾危害严重区域,可以考虑选择种植马铃薯块茎蛾适应性较低的品种,作为田间抑制马铃薯块茎蛾种群的一个重要策略。
马铃薯块茎蛾;年龄-阶段特定两性生命表;马铃薯品种;品种适应性
0 引言
【研究意义】马铃薯块茎蛾()属鳞翅目,麦蛾科,是一种广泛分布于热带和亚热带地区的世界性茄科害虫[1],也是国际和国内检疫对象,尤以马铃薯()和烟草受害最重。此外,也可取食危害其他植物,如龙葵、银叶草、茄子等。马铃薯块茎蛾在马铃薯生长期和贮藏期均可产生危害[2],其田间危害可使马铃薯减产20%—30%;马铃薯收获后,一部分马铃薯块茎蛾可以继续遗留在田间的马铃薯块茎或自播植物上生长;另一部分随马铃薯贮藏,在长达4个月的储藏期中,若无冷藏,危害率可达100%[3]。马铃薯是仅次于玉米、水稻和小麦的第四大粮食作物[4-5]。在保障我国粮食安全、精准扶贫、种植业结构调整以及农业产业转型升级中发挥着至关重要的作用[5]。我国马铃薯的种植面积和总产量均位居世界首位,但单产水平因病虫害等瓶颈因素的制约,低于世界平均水平[5]。害虫是制约马铃薯商品化生产的主要因素之一。【前人研究进展】从20世纪初,就开始有关于马铃薯块茎蛾危害马铃薯的报道[6],是世界范围内影响马铃薯的主要害虫之一[7]。国外已进行了马铃薯品种抗性方面的研究。如国际马铃薯中心从众多马铃薯品系中筛选到一些抗性品系。Rondon等[8]在大田和实验室条件下筛选对马铃薯块茎蛾具有抗性的种质资源;Mansouri等[9]进行了伊朗马铃薯种质资源对马铃薯块茎蛾抗性的筛选工作。早在20世纪30年代,国外就有研究者开始运用天敌昆虫来防治马铃薯块茎蛾。Coll等[10]报道了以色列防治马铃薯块茎蛾的5种寄生性天敌(寄生蜂)和7种捕食性天敌(七星瓢虫1种、草蛉1种、蝽类1种、蚂蚁4种),其寄生率达到40%,捕食率达到79%。而国内主要采用化学手段防控马铃薯害虫,化学农药的密集使用不仅对环境、食品安全和人类健康产生负面影响,而且会导致昆虫产生抗药性[11],同时对天敌昆虫不利。另外,马铃薯块茎蛾作为一种钻蛀性害虫,杀虫剂很难直接接触到害虫,防治效果较差。近年来,研究者开始尝试采用生物和物理手段防控马铃薯块茎蛾,如YUAN等在室内筛选了对马铃薯块茎蛾幼虫有致病力的球孢白僵菌()菌株[7,12],YAN等[13]在室内采用斯氏线虫()防治马铃薯块茎蛾,但在实际应用中其防治效果受到很多不确定环境因素的影响,例如白僵菌对温湿度、光照具有严格的要求,不利于田间应用。马艳粉等[14]研究了植物挥发物对马铃薯块茎蛾的产卵引诱作用,但性诱剂仅对成虫起作用,另外实施这种推-拉策略,是否会使周围其他经济作物产生虫害还未可知。【本研究切入点】品种自然抗虫性是植物本身所具有的一种遗传特性,受环境限制小,对环境友好,可作为马铃薯害虫综合防控的重要策略。然而马铃薯品种抗虫性自然资源的挖掘相关研究在国内还处在一个空白阶段,因此筛选对马铃薯害虫自然抗性较高的马铃薯品种资源非常必要。夏波蒂、龙薯4号、龙薯12号和克新17号在北方主要马铃薯产区推广应用较为普遍,且市场需求量较高。【拟解决的关键问题】基于年龄-阶段两性生命表,在室内测定取食上述4种品种块茎后马铃薯块茎蛾种群各阶段的发育历期和产卵量,明确不同马铃薯品种对马铃薯块茎蛾生长发育和繁殖的影响,为筛选抗性品种并制定马铃薯块茎蛾综合防控策略提供依据。
1 材料与方法
试验于2019—2020年在中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室完成。
1.1 供试虫源
马铃薯块茎蛾于2013年6月采集自云南昆明,共500头左右,寄主植物为马铃薯。根据桂富荣等[15]和Rondon等[8]的方法,在中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室建立马铃薯块茎蛾实验种群。成虫饲养在玻璃罐(直径×高=14 cm×6 cm)内,在柱形容器顶端用医用纱布盖上,橡皮筋封口,在纱布上放置一张直径为8 cm的滤纸作为产卵基质,每个瓶子内放一个直径约为6 cm的马铃薯块茎供幼虫取食,并在瓶子底部铺一层刚好覆盖瓶底的细沙,为幼虫化蛹提供场所。小号塑料培养皿内塞脱脂棉,放入玻璃罐内用以盛蜂蜜水做成虫食物来源。成虫羽化后,用滴管将配备好的10%蜂蜜水滴在脱脂棉上供成虫取食。饲养于三洋人工气候箱内(MLR-351H,SANYO Electric Co,Ltd),环境温度为(26±1)℃,光周期为12 h﹕12 h(L﹕D),相对湿度(70±10)%。
试验所用全部完整卵粒均为1日龄成虫交配24 h后获得。
1.2 马铃薯品种
龙薯4号、龙薯12号与克新17号均由黑龙江省农业科学院提供,夏波蒂购自北京市回龙观菜市场。
1.3 4种块茎品种上马铃薯块茎蛾各虫态发育历期测定
试验在室内三洋人工气候箱中进行(气候条件设定同1.1)。从饲养种群中取下卵纸,借助于双目体视显微镜(XTZ-D,上海光学仪器一厂)用零号毛笔从卵纸上随机收集150头完整卵粒,为保证卵粒完整,在双目体视显微镜(XTZ-D,上海光学仪器一厂)下进行挑选,然后将挑好的150粒卵平均转移至3个饲养盒(每盒50粒卵)(长×宽×高=25 cm×15 cm×8 cm),每盒放入约700 g块茎(参考文献[16],马铃薯块茎蛾的饲养密度建议以1头/13.0—26.0 g块茎为宜)。每个品种处理方法相同。接卵后每日观察卵的孵化数和死亡数,统计卵存活率;待其化蛹后,统计幼虫死亡数,并将蛹放入2 mL离心管内;待其羽化后,统计蛹存活率,并对成虫进行雌雄鉴别、配对,配对时将离心管里的一对雌、雄成虫接入塑料瓶(高×直径=14 cm×6 cm),瓶口用医用纱布盖封口,在纱布上放一张滤纸(直径=5 cm),并用玻璃片压住,每隔24 h记录产卵量,每日更换滤纸和蜂蜜水。
1.4 幼虫对4种品种块茎的选择性取食
采用王冬妍等[17]培养皿叶盘选择性试验方法,将切好的4种马铃薯块茎(长×宽×高=2 cm×2 cm×1 cm)等距放入直径15 cm的圆形玻璃培养皿中,用零号毛笔挑取20头初孵1龄幼虫,接入玻璃培养皿的中央,封口膜封住,扎孔透气。移入三洋人工气候箱内(MLR-351H,SANYO Electric Co,Ltd),环境温度为(26±1)℃,光周期为12 h﹕12 h(L﹕D),相对湿度(70±10)%。24 h后统计各块茎上的幼虫数。2、3、4龄幼虫处理同上。每个处理设置10次重复。
1.5 数据分析
马铃薯块茎蛾各阶段发育历期、雌雄虫寿命和雌成虫产卵量均使用年龄-阶段两性生命表进行分析[18-20]。年龄-阶段特定存活率(age-stage specific rate,S)由每日记录的存活数计算得到,雌虫年龄-阶段生殖力(age-stage fecundity,f)由每日记录的产卵量计算得到。根据CHI等[18]计算方法,其中代表年龄,代表阶段,代表龄期数。s′代表一个年龄,阶段的个体存活到年龄阶段的概率,假设s=1。
计算得到以上数据后,根据以下公式分别算出特定年龄存活率(age-specific survival rate,)、特定年龄生殖力(age-specific fecundity,m)、年龄期望(life expectancy,e)。

生命表参数内禀增长率(intrinsic rate of increase,)表示在给定的物理和生物条件下,具有稳定年龄组配种群的最大瞬时增长率,可通过使用二分迭代法[21]和Euler-Lotka方程计算,计算公式如下:,初始年龄为0。
周限增长率(finite rate of increase,)指在不受外界环境限制的情况下,种群平均每天增长的速率。计算公式如下:
λ=e
净增殖率(net reproductive rate,R)指一个个体一生中所产生的后代数。计算公式如下:
平均世代时间(mean generation time,)指种群达到稳定增长率(和)时,增加R所需的时间。=(lnR)/。
根据上述计算公式,在Visual BASIC环境下使用TWOSEX-MSChart[22]对原始数据和生命表参数进行相关运算。利用bootstrap技术估计种群参数的方差和标准误差[23-24],采用TWOSEX-MSChart程序里的Paired bootstrap test对数据进行差异显著性检验,Sigmaplot 12.5软件作图。
采用单因素方差分析(ANOVA)和Tukey HSD成对检验对幼虫选择性取食进行显著性分析。在R软件环境中进行统计分析(v.3.0.3)。
2 结果
2.1 4个品种块茎对马铃薯块茎蛾生长发育和繁殖的影响
4个品种块茎饲养的马铃薯块茎蛾卵期发育时间、成虫平均寿命与雄成虫寿命无显著差异。而幼虫期、蛹期发育时间、雌成虫寿命、单雌产卵量存在极显著差异(=7.463,=11.44,=8.975,=13.01,<0.001)。取食克新17号的马铃薯块茎蛾幼虫期和蛹期(15.18、8.10 d)高于夏波蒂、龙薯4号和龙薯12号(14.14、13.52和14.46 d,7.89、7.48和7.28 d),而雌成虫寿命和单雌产卵量(8.46 d、165.71粒)低于其他3个品种(13.50 d、235.06粒,12.11 d、254.48粒,11.00 d、171.79粒)。其中,龙薯4号和克新17号上的马铃薯块茎蛾差异最为显著(表1)。
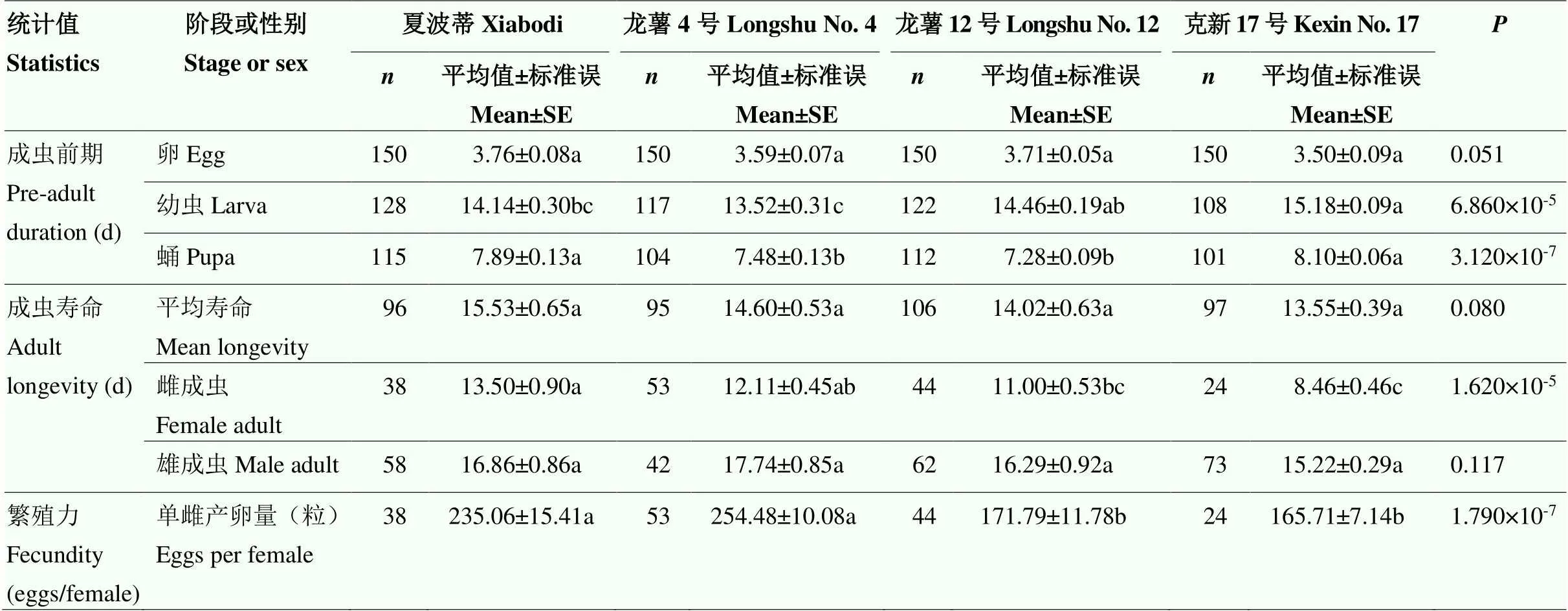
表1 取食4种马铃薯品种块茎的马铃薯块茎蛾生活史
“”代表该时期马铃薯块茎蛾存活个体数。采用Paired bootstrap test对数据进行差异显著性检验,同行数据后不同小写字母表示差异显著(<0.05)。表2同
“” represents the number of surviving individuals ofduring this period. Values followed by the different lowercase letters within a row indicate significantly different using paired bootstrap test (<0.05). The same as Table 2
2.2 4个品种块茎对马铃薯块茎蛾存活率和繁殖率的影响
存活率曲线s描述的是一粒卵发育至年龄,阶段的可能性。曲线的重叠是由于个体间发育速率不同造成的,重叠程度越高,代表个体之间发育速率差异越大,且不稳定[24]。如图1所示,马铃薯块茎蛾各组存活率曲线重叠程度较低,表明不同品种之间各阶段发育速率比较一致。另外取食不同品种块茎的马铃薯块茎蛾存在一个共同现象,即雄成虫的存活率高于雌成虫。相较于取食其他3种块茎(82.72%、80.27%和84.36%),取食克新17号的卵存活率最低(74.62%);而整个幼虫和蛹阶段,取食各品种的马铃薯块茎蛾存活率无明显变化。
特定年龄存活率(l)、雌虫特定年龄繁殖力(f)、群体特定年龄繁殖力(m)和特定年龄繁殖值(lm)如图2所示。f是年龄阶段的日平均产卵量[25]。马铃薯块茎蛾第4阶段时开始产卵(f,4),取食夏波蒂、龙薯4号、龙薯12号和克新17号的雌成虫日产卵量最高峰值均在25 d左右,分别为(26.09 d,59.34粒)、(23.75 d,54.71粒)、(25.75 d,45.50粒)和(26.03 d,46.90粒),取食龙薯12号的产卵峰值最低,取食克新17号的次之。另外,有的曲线有2个或2个以上的峰值,这是由于有个别配对的雄成虫寿命较雌虫短,重新配对后导致再次出现产卵峰值。
l是年龄的存活率,描述了群体存活率随年龄的变化[25]。取食克新17号的马铃薯块茎蛾迅速下降,且下降速度快于其他3个品种。另外第50天左右,取食龙薯12号和克新17号的马铃薯块茎蛾全部死亡;第60天左右,夏波蒂和龙薯4号上的成虫全部死亡。因此取食夏波蒂和龙薯4号的马铃薯块茎蛾寿命长于取食龙薯12号和克新17号(图2)。
m表示年龄时所有个体平均生产子代数[25]。m曲线显示,取食夏波蒂、龙薯4号、龙薯12号和克新17号的马铃薯块茎蛾m峰值分别为10.58、22.71、14.98和8.74。其中取食龙薯4号的马铃薯块茎蛾m值最高,克新17号上m值最低(图2)。
lm是l与m的乘积,是年龄时所有存活个体的繁殖值,即总单雌产卵量值[25]。取食夏波蒂、龙薯4号、龙薯12号和克新17号的马铃薯块茎蛾lm峰值分别为7.32、14.58、10.82和5.27,在龙薯4号上马铃薯块茎蛾繁殖最快,克新17号上繁殖最慢(图2)。
特定年龄-阶段生命期望(e)表示年龄阶段个体的剩余存活时间,生命期望会随年龄增长逐渐降低[25]。如图3所示,取食各品种块茎的马铃薯块茎蛾中存在一个普遍现象,即雄成虫的生命期望明显高于雌成虫。另外,克新17号上的马铃薯块茎蛾幼虫阶段生命期望值高于其他3个品种,这表明相较于其他3个品种,在克新17号上马铃薯块茎蛾幼虫发育较慢。这与克新17号上马铃薯块茎蛾幼虫历期最长结果一致。
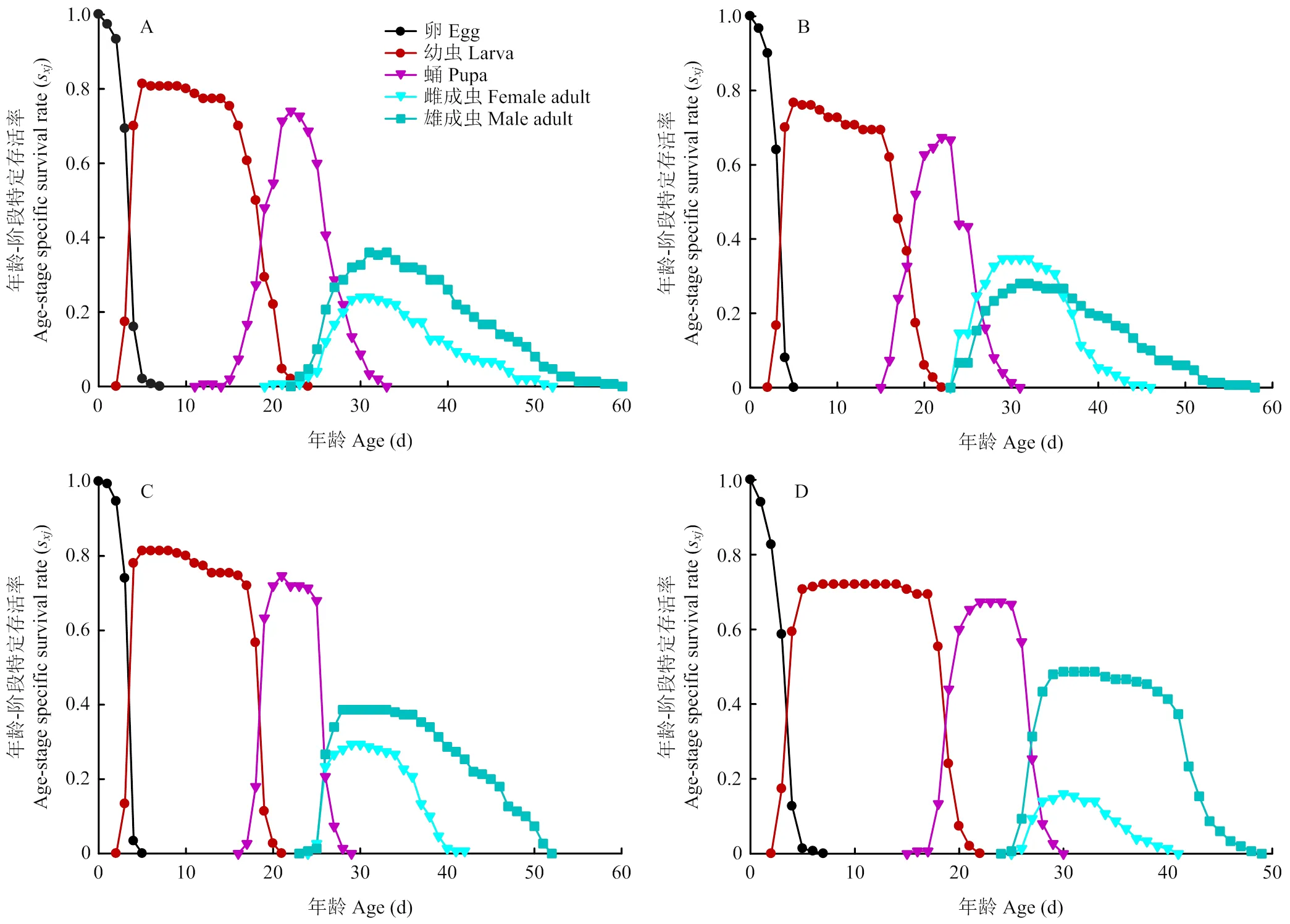
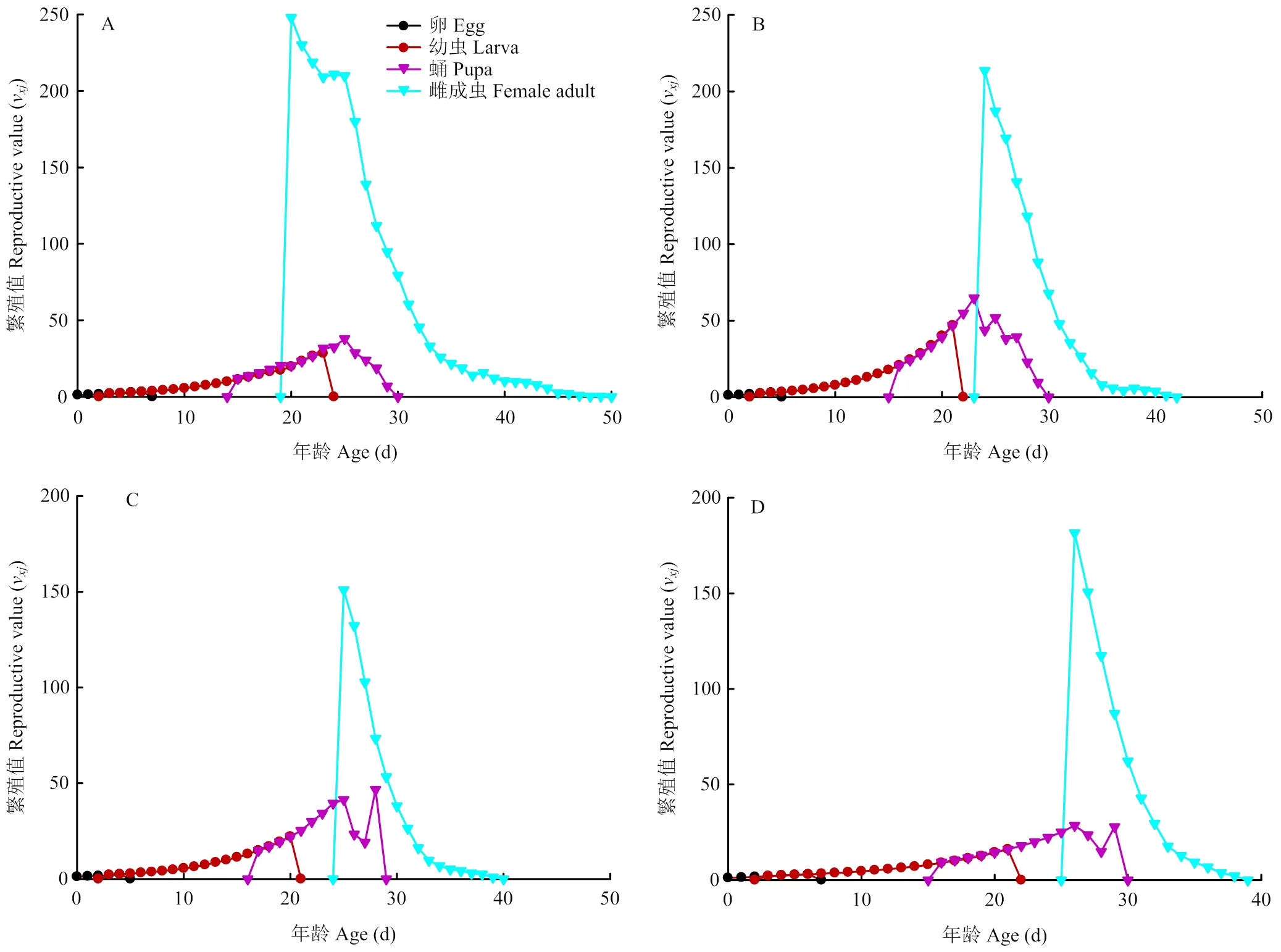
A:夏波蒂Xiabodi;B:龙薯4号Longshu No. 4;C:龙薯12号Longshu No. 12;D:克新17号Kexin No. 17
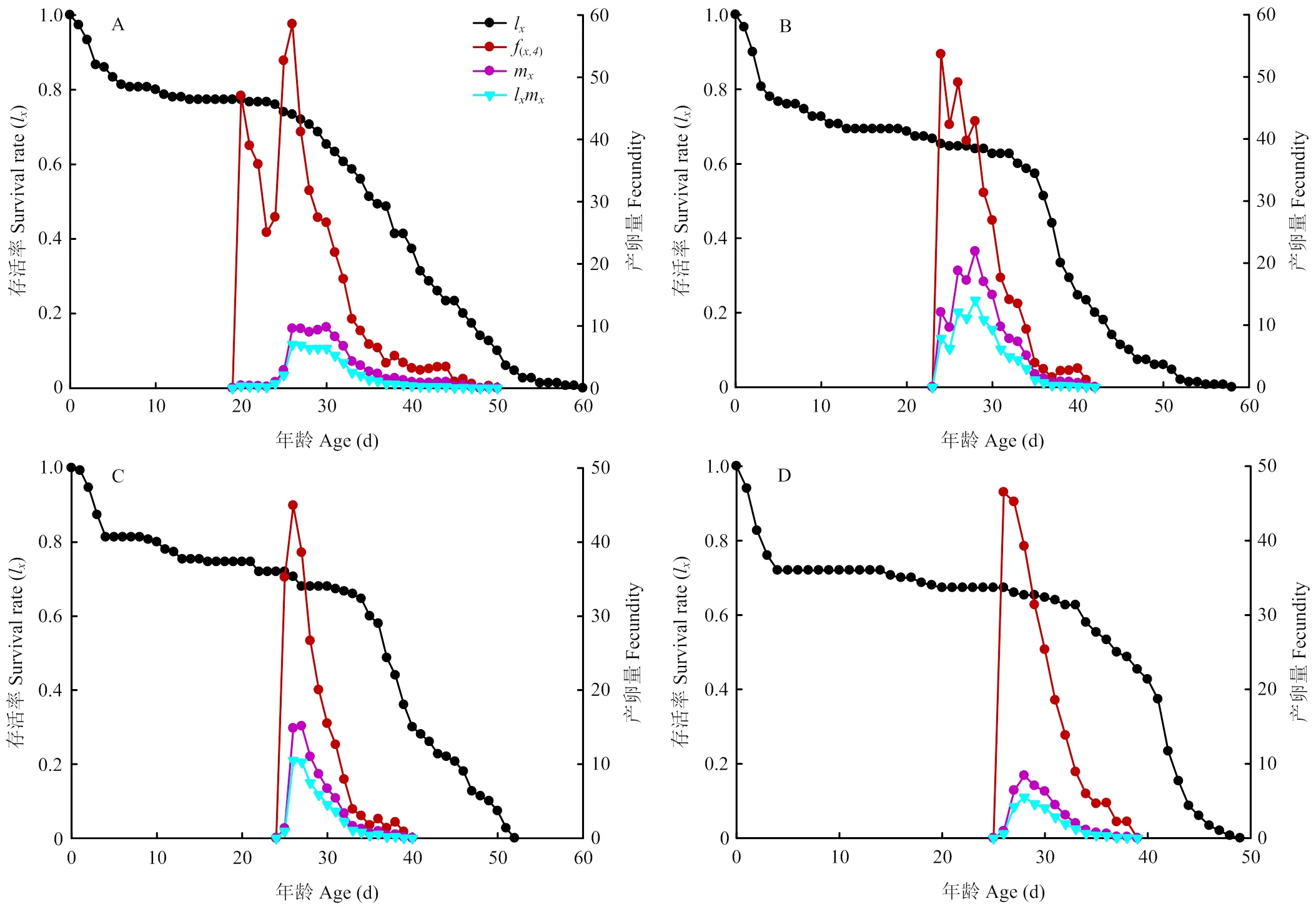
A:夏波蒂Xiabodi;B:龙薯4号Longshu No. 4;C:龙薯12号Longshu No. 12;D:克新17号Kexin No. 17
特定年龄-阶段繁殖值(v)是指年龄阶段的个体对未来种群增长的平均贡献度[25]。初孵幼虫的繁殖值(01)等于周限增长率()[25]。取食夏波蒂、龙薯4号、龙薯12号和克新17号的01分别为1.145、1.170、1.143和1.125。其中,取食夏波蒂品种的雌蛾繁殖值峰值出现最早(20.07 d),且峰值为251.41粒,取食其他3种的马铃薯块茎蛾繁殖峰值出现日期及峰值分别为(24.09 d,215.45粒)、(25.16 d,154.83粒)和(25.75 d,184.68粒)。相较于夏波蒂繁殖峰值出现最早,且繁殖值最大,龙薯12号繁殖值峰值最小,克新17号品种上出现最晚,且克新17号上雌虫繁殖结束最早(37.72 d)(图4)。
2.3 4个块茎品种对马铃薯块茎蛾后代种群参数的影响
取食龙薯4号的马铃薯块茎蛾的、和R值分别为0.157 d-1、1.170 d-1和92.740,显著高于夏波蒂(0.135 d-1、1.145 d-1和56.573)、龙薯12号(0.134 d-1、1.143 d-1和48.640)和克新17号(0.108 d-1、1.125 d-1和26.513)(=1301009,=1311478,=104452,<0.001)。此外,取食龙薯4号的马铃薯块茎蛾值(28.87 d)显著低于克新17号(30.28 d)、夏波蒂(29.85 d)和龙薯12号(29.10 d)(= 480154,<0.001)(表2)。
2.4 马铃薯块茎蛾幼虫对4个品种块茎的选择性取食
初孵幼虫对4种块茎的取食选择为龙薯4号>夏波蒂>龙薯12号>克新17号,4种块茎上的幼虫数平均分别为9.3、8.0、1.6和1.1头;2龄幼虫数分别为8.0、6.9、3.0和2.1头;3龄幼虫数分别为6.5、5.9、4.0和3.6头;4龄幼虫数分别为5.4、5.4、4.5和4.7头。随着龄期变大,选择性逐渐减弱,但马铃薯块茎蛾对龙薯4号和夏波蒂取食选择性始终显著高于龙薯12号和克新17号(=168.7,=30.07,=7.938,<0.001)(图5)。
3 讨论
昆虫-植物的互作关系对于制定可持续的害虫控制策略至关重要[26]。这一问题促使研究人员努力寻找有效的替代控制方法,包括寄主植物抗性[27-31]。植物抗虫性涉及可遗传特性,使植物能够避免、容忍或从虫害的影响中恢复[32-33]。寄主植物作为昆虫幼虫的食物,其质量因植物种类而异[34]。因此,昆虫幼虫的发育和存活受到寄主植物的强烈影响[35]。年龄-阶段两性生命表可以全面、准确地描述昆虫种群在特定实验条件下的表现[36-37]。本研究采用年龄-阶段两性生命表,对马铃薯块茎蛾在几种重要的马铃薯品种(夏波蒂、龙薯4号、龙薯12号和克新17号)上的生物学特性进行了系统评估,研究发现马铃薯块茎蛾在这4个品种上均能完成世代发育,但是取食不同品种的马铃薯块茎蛾在存活和发育时间上存在显著差异。成虫阶段,各品种上雄成虫寿命无明显差别,但雌成虫寿命间存在显著差异。取食品种克新17号的雌成虫寿命、单雌产卵量低于另外3个品种。研究表明,合适的寄主对卵和早期幼虫极为重要,它们的生活史特征可能会受到取食不合适寄主的负面影响[35,38]。产卵前期越长,内禀增长率越低。此外,内禀增长率是反映特定环境条件下昆虫种群增长潜力的重要参数,该值越大,则昆虫种群发展越快[39-40]。因此,在供试的4个品种中,克新17号为最不利于马铃薯块茎蛾生长发育的寄主品种。

表2 取食4个块茎品种的马铃薯块茎蛾种群参数
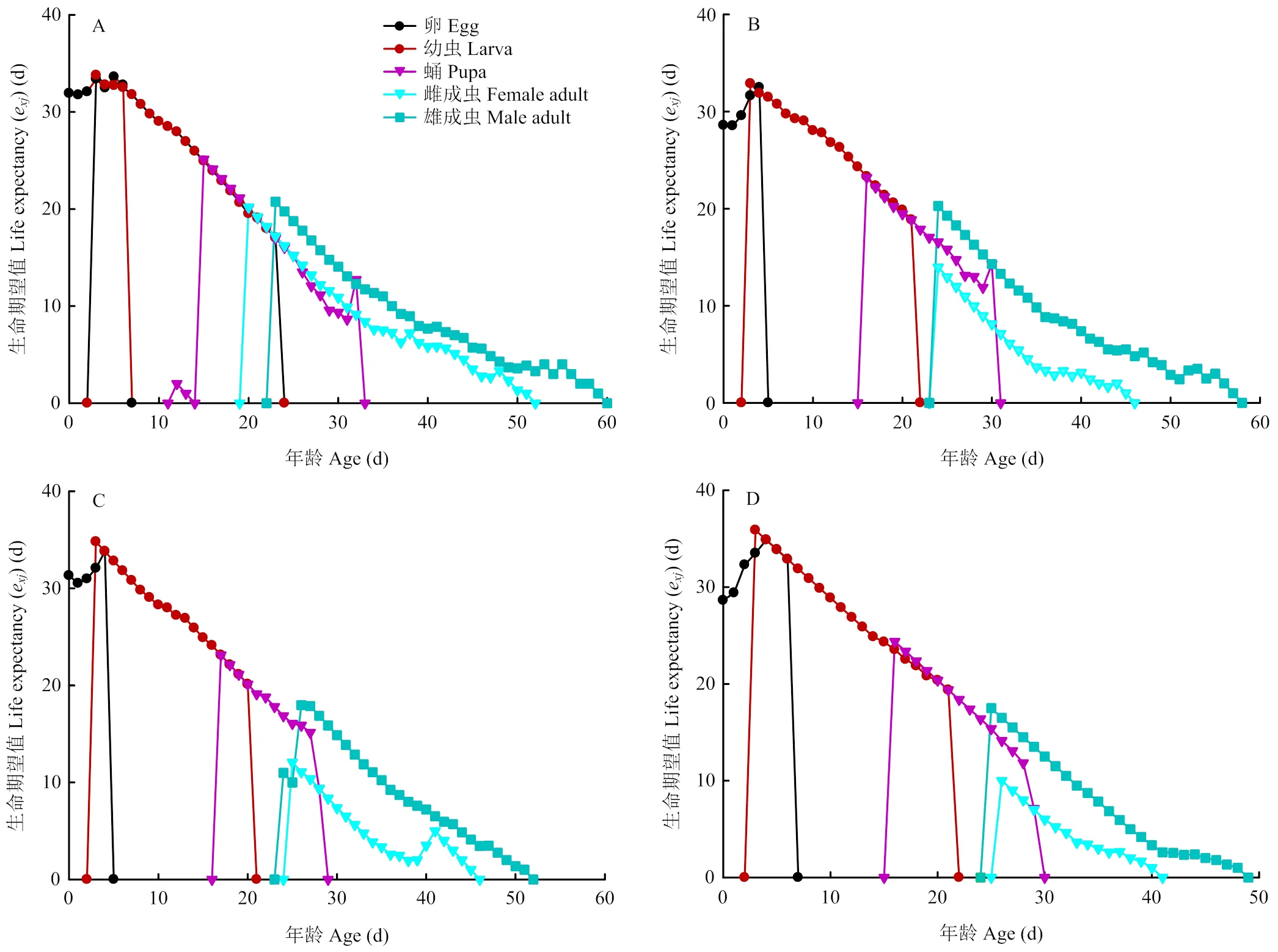
A:夏波蒂Xiabodi;B:龙薯4号Longshu No. 4;C:龙薯12号Longshu No. 12;D:克新17号Kexin No. 17
A:夏波蒂Xiabodi;B:龙薯4号Longshu No. 4;C:龙薯12号Longshu No. 12;D:克新17号Kexin No. 17
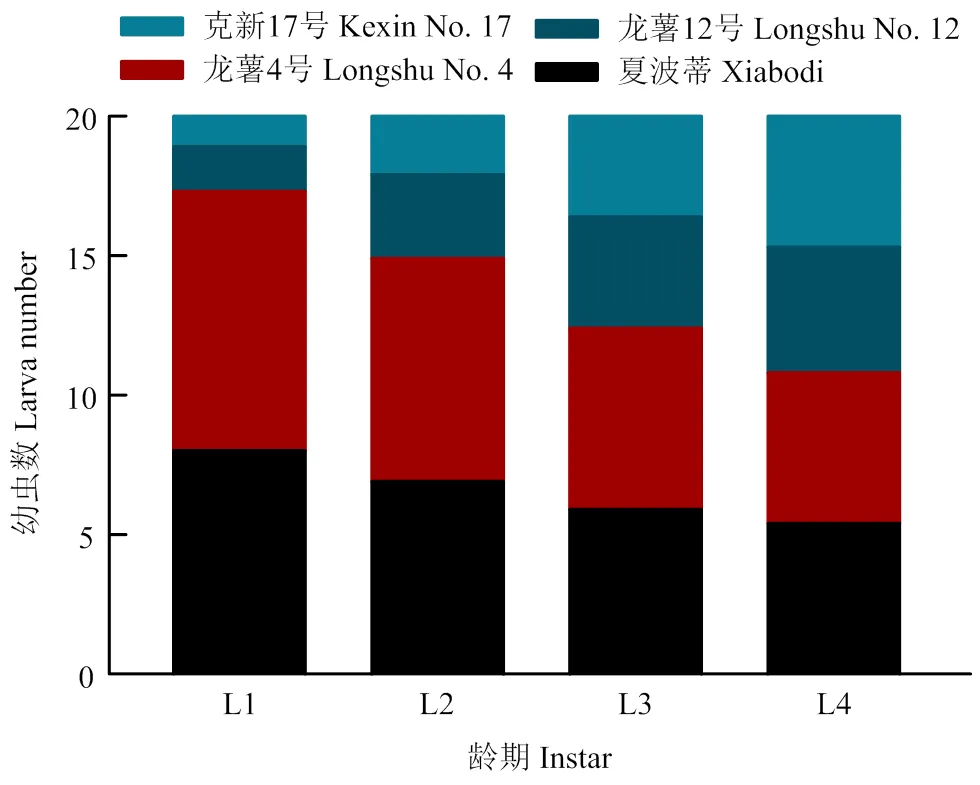
图5 马铃薯块茎蛾不同龄期幼虫对4个品种块茎的选择性取食
马铃薯表皮粗糙程度和结构与幼虫存活率显著相关[8],在本试验中,卵存活率是根据钻蛀进薯块的幼虫数进行计算的。卵孵化后能否成功钻蛀,可能会导致不同品种间卵发育时间和孵化率出现差异。因此,在本试验中卵期不一致,可能与马铃薯表皮粗糙程度和结构有关。
不同的马铃薯种质会导致幼虫发育时间和成虫生殖力存在显著差异[37]。Mansouri等[9]研究表明,马铃薯淀粉、矿物质等必需成分含量较低会导致幼虫发育时间延长。在淀粉和矿物质含量较高的块茎上饲养的雌蛾,其卵巢中会产生更多的卵,而在淀粉和矿物质含量较低的块茎上饲养的雌蛾产卵量较少。因此,推测夏波蒂[41]和龙薯4号[42]淀粉含量可能高于克新17号。
本研究中,1龄幼虫时马铃薯块茎蛾对龙薯4号表现出极高的选择性,对克新17号选择性最低,且成虫阶段,取食克新17号的雌成虫平均产卵量最低,该结果支持偏好-表现假说,即植食性昆虫雌虫通过选择与幼虫表现强相关的寄主植物来最大化适应度[43-44]。闫俊杰等[45]研究提出寄主植物的品质会影响害虫偏好。不过还需在田间调查雌成虫产卵偏好,进一步验证这一结论。
开展不同马铃薯品种对马铃薯块茎蛾抗性评价工作,对马铃薯块茎蛾的综合治理具有重要意义[46]。然而,马铃薯块茎蛾抗性品种筛选方面的研究较少[47-49]。年龄-阶段两性生命表可以全面、准确地描述昆虫种群在特定实验条件下的表现[36-37]。通过马铃薯块茎蛾在不同品种块茎上生长发育和繁殖能力的差异比较,有望筛选出对马铃薯块茎蛾抗性较强的品种。将抗虫品种与杀虫剂结合使用,并辅以适当的栽培措施,如底膜覆盖和限制田间暴露时间,应该是未来最佳的管理选择[8]。
本研究仅重点关注了马铃薯块茎抗虫性评价,而实际上马铃薯块茎蛾初孵幼虫大部分先钻入叶肉进行危害,随着幼虫的生长发育,逐渐开始爬出叶肉钻入块茎中危害。叶片受损会影响植物的光合作用,进而影响马铃薯产量和品质。因此,下一步需开展田间工作,观察马铃薯块茎蛾在不同品种叶片上的发生情况。探究叶片受损与块茎受损的相关性,才能更加准确的筛选出马铃薯块茎蛾叶片和块茎同时适应性较低的品种。
4 结论
使用生命表方法研究马铃薯块茎蛾在不同马铃薯品种块茎上的危害情况,可以作为评价马铃薯对马铃薯块茎蛾自然抗性的一个重要指标。供试品种中,马铃薯块茎蛾在夏波蒂和龙薯4号块茎上危害较重,龙薯12号次之,克新17号最轻。因此,在我国马铃薯重要的种植区,可以考虑当地实际情况,优先选择种植克新17号或龙薯12号,以减轻马铃薯块茎蛾的田间危害。
[1] FENEMORE P G. Host-plant location and selection by adult potato moth,(Lepidoptera: Gelechiidae): A review. Journal of Insect Physiology, 1988, 34(3): 175-177.
[2] 李秀军, 金秀萍, 李正跃. 马铃薯块茎蛾研究现状及进展. 青海师范大学学报(自然科学版), 2005(2): 67-70.
LI X J, JIN X P, LI Z Y. The present status and developing tendency inresearch.Journal of Qinghai Normal University (Natural Science edition), 2005(2): 67-70. (in Chinese)
[3] 孙茂林, 李先平, 赵志坚, 杨万林, 段玉云, 李树莲, ENRIQUE C. 云南马铃薯贮存损失的调查和评价. 中国马铃薯, 2002, 16(5): 263-266.
SUN M l, LI X P, ZHAO Z J, YANG W L, DUAN Y Y, LI S L, ENRIQUE C. The investigation and evaluation on the losses of potato storage in Yunnan. Chinese Potato Journal, 2002, 16(5): 263-266. (in Chinese)
[4] 高玉林, 徐进, 刘宁, 周倩, 丁新华, 詹家绥, 成新跃, 黄剑, 鲁宇文, 杨宇红. 我国马铃薯病虫害发生现状与防控策略. 植物保护, 2019, 45(5): 106-111.
GAO Y L, Xu J, LIU N, ZHOU Q, DING X H, ZHAN J S, CHENG X Y, HUANG J, LU Y W, YANG Y H. Current status and management strategies for potato insect pests and disease in China. Plant Protection, 2019, 45(5): 106-111. (in Chinese)
[5] 徐进, 朱杰华, 杨艳丽, 汤浩, 吕和平, 樊明寿, 石瑛, 董道峰, 王贵江, 王万兴, 熊兴耀, 高玉林. 中国马铃薯病虫害发生情况与农药使用现状. 中国农业科学, 2019, 52(16): 2800-2808.
XU J, ZHU J H, YANG Y L, TANG H, Lü H P, FAN M T, SHI Y, DONG D F, WANG G J, WANG W X, XIONG X Y, GAO Y L. Status of major diseases and insect pests of potato and pesticide usage in China. Scientia Agricultura Sinica, 2019, 52(16): 2800-2808. (in Chinese)
[6] GRAF J E. The potato tuber moth. Bulletin of the United States Department of Agriculture, 1917, 427: 58.
[7] YUAN H G, WU S Y, LEI Z R, RONDON S I, GAO Y L. Sublethal effects ofon field populations of the potato tuberworm(Zeller) in China. Journal of Integrative Agriculture, 2018, 17(4): 911-918.
[8] RONDON S I, HANE D C, BROWN C R, VALES M I, DOGRAMACI M. Resistance of potato germplasm to the potato tuberworm (Lepidoptera: Gelechiidae). Journal of Economic Entomology, 2009, 102(4): 1649-1653.
[9] MANSOURI S M, FATHI S A A, NOURI-GANBALANI G, RAZMJOU J, NASEIR B, RONDON S I. Screening of Iranian potato germplasm for resistance to the ptato tuberworm(Lepidoptera: Gelechiidae). American Journal of Potato Research, 2013, 90(6): 533-540.
[10] COLL M, GAVISH S, DORI I. Population biology of the potato tuber moth,(Lepidoptera: Gelechiidae) in two potato cropping systems in Israel. Bulletin of Entomological Research, 2000, 90(4): 309-315.
[11] DOGRAMACI M, TINGEY W M. Comparison of insecticide resistance in a North American field population and a laboratory colony of potato tuberworm (Lepidoptera: Gelechiidae). Journal of Pest Science, 2008, 81(1): 17-22.
[12] 孙跃先, 李正跃, 桂富荣, 严乃胜, 陈斌, 许若清, 和淑琪. 白僵菌对马铃薯块茎蛾致病力的测定. 西南农业学报, 2004, 17(5): 627-629.
SUN Y X, LI Z Y, GUI F R, YAN N S, CHEN B, XU R Q, HE S Q. Susceptiblity of(Zeller) larvae to 8 isolates of. Southwest China Journal of Agricultural Sciences, 2004, 17(5): 627-629. (in Chinese)
[13] YAN J J, SHOVON C S, MENG R X, REITZ S, GAO Y L. Potential of(Weiser) as a biological control agent against potato tuber moth,(Zeller) (Lepidoptera: Gelechiidae). Journal of Integrative Agriculture, 2020, 19(2): 389-393.
[14] 马艳粉, 胥勇, 肖春. 10种寄主植物挥发物对马铃薯块茎蛾产卵的引诱作用. 中国生物防治学报, 2012, 28(3): 448-452.
MA Y F, XU Y, XIAO C. Oviposition attraction effect of ten host-plant volatiles on potato tuber moth,.Chinese Journal of Biological Control, 2012, 28(3): 448-452. (in Chinese)
[15] 桂富荣, 李正跃. 用马铃薯人工饲养马铃薯块茎蛾的方法. 昆虫知识, 2003, 40(2): 187-189.
GUI F R, LI Z Y. A method for rearing the potato tuber mothon potato. Entomological Knowledge, 2003, 40(2): 187-189. (in Chinese)
[16] 马艳粉, 胥勇, 李娜, 李正跃, 何月秋, 肖春. 幼虫密度对马铃薯块茎蛾生长发育及繁殖的影响. 昆虫知识, 2010, 47(4): 694-699.
MA Y F, XU Y, LI N, LI Z Y, HE Y Q, XIAO C. Effect of larval density on growth, development and reproduction of potato tuber moth,.Chinese Bulletin of Entomology, 2010, 47(4): 694-699. (in Chinese)
[17] 王冬妍, 王振营, 何康来, 丛斌. 转Bt基因抗虫玉米对亚洲玉米螟幼虫取食行为的影响. 昆虫知识, 2005, 42(3): 270-274.
WANG D Y, WANG Z Y, HE K L, CONG B. Feeding behavior oflarvae on transgenic Bt corn expressing Cry1Ab toxin. Chinese Bulletin of Entomology, 2005, 42(3): 270-274. (in Chinese)
[18] CHI H, LIU H. Two new methods for the study of insect population ecology. Academia Sinica, 1985, 24(2): 225-240.
[19] CHENG C, HUANG C Y, DAI S M, ATLIHAN R, CHI H. Genetically engineered ricin suppresses(Diptera: Tephritidae) based on demographic analysis of group-reared life table. Journal of Economic Entomology, 2016, 109(3): 987-992.
[20] CHI H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environmental Entomology, 1988(1): 26-34.
[21] GOODMAN D. Optimal life histories, optimal notation, and the value of reproductive value. The American Naturalist, 1982, 119(6): 803-823.
[22] CHI H. TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis. Taichung, Taiwan: National Chung Hsing University, 2009.
[23] EFRON B, TIBSHIRANI R J. An introduction to the bootstrap. Chapman and Hall, New York, 1993.
[24] HUANG Y B, CHI H. Age-stage, two-sex life tables of(Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Science, 2012, 19(2): 263-273.
[25] CHI H, YANG T C. Two-sex life table and predation rate ofThunberg(Coleoptera: Coccinellidae) fed on(Sulzer) (Homoptera: Aphididae). Environmental Entomology, 2003, 32(2): 327-333.
[26] PICKETT J A, WADHAMS L J, WOODCOCK C M. Developing sustainable pest control from chemical ecology. Agriculture, Ecosystems and Environment, 1997, 64(2): 149-156.
[27] GURR G M, SYMINGTON C A. Resistance to the potato tubermoth,(Zeller) (Lepidoptera: Gelechiidae), in potato (ssp.L.) tubers. Australian Journal of Entomology, 1998, 37(1): 49-51.
[28] SMITH C M. Plant resistance toarthropods. The Netherlands: Springer, 2005.
[29] DOGRAMACI M, TINGEY W M. Host resistance and influence of tuber surface on larval performance of potato tuberworm (Lepidoptera: Gelechiidae). Journal of Pest Science, 2009, 82(2): 109-114.
[30] HORGAN F G, QUIRING D T, LAGNAOUI A, PELLETIER Y. Tradeoff between foliage and tuber resistance toin wild potatoes. Entomologia Experimentalis et Applicata, 2009, 131(2): 130-137.
[31] RONDON S I, BROWN C R, MARCHOSKY R. Screening for resistance of potato line to the potato tuberworm,(Zeller) (Lepidoptera: Gelechiidae). American Journal Potato Research, 2013, 90(1): 71-82.
[32] TINGEY W M, LAUBENGAYER J E. Glandular trichomes of a resistant hybrid potato alter feeding behavior of the potato leafhopper (Homoptera: Cicadellidae). Journal of Economic Entomology, 1986, 79(5): 1230-1234.
[33] PANDA N, KHUSH G S.Host plant resistance to insect. CAB International, 1995.
[34] AWMACK C S, LEATHER S R. Host plant quality and fecundity in herbivorous insects. Annual Review of Entomology, 2002, 47: 817-844.
[35] MCCORMICK A C, ARRIGO L, EGGENBERGER H, MESCHER M C, DE MORAES C M. Divergent behavioral responses of gypsy moth () caterpillars from three different subspecies to potential host trees. Scientific Reports, 2019, 9(1): 8953.
[36] AKCA I, AYVAZ T, YAZICI E, SMITH C L, CHI H. Demography and population projection of(Hemiptera: Aphididae): with additional comments on life table research criteria.Journal of Economic Entomology, 2015, 108(4): 1466-1478.
[37] TUAN S J, YEH C C, ATLIHAN R, CHI H, TANG L C. Demography and consumption of(Lepidoptera: Noctuidae) reared on cabbage and taro. Journal of Economic Entomology, 2015, 109(2): 732-739.
[38] ZALUCKI M P, CLARKE A R, MALCOLM S B. Ecology and behavior of first instar larval Lepidoptera. Annual Review of Biochemistry, 2002, 47: 361-393.
[39] 胡良雄, 何正盛, 张小谷. 猫眼尺蠖在三种女贞属植物上的实验种群两性生命表. 昆虫学报, 2014, 57(12): 1408-1417.
HU L X, HE Z S, ZHANG X G. Age-stage two-sex life tables of the experimental population of(Lepidoptera: Geometridae) on threespecies. Acta Entomologica Sinica, 2014, 57(12): 1408-1417. (in Chinese)
[40] 秦建洋, 张蕾, 程云霞, 罗礼智, 雷朝亮, 江幸福. 不同温度下东方粘虫年龄-阶段两性实验种群生命表的构建. 植物保护学报, 2017, 44(5): 729-736.
QIN J Y, ZHANG L, CHENG Y X, LUO L Z, LEI C L, JIANG X F. Age-stage two-sex life table for laboratory populations of oriental armyworm(Walker) under different temperatures. Journal of Plant Protection, 2017, 44(5): 729-736. (in Chinese)
[41] 刘江娜, 张爱萍, 王航, 赵亮. 新疆加工马铃薯夏波蒂品种引育及栽培技术要点. 种子, 2018, 37(2): 109-110.
LIU J N, ZHANG Y P, WANG H, ZHAO L. The main points of breeding and cultivation technique of potato variety Xiabodi from Xinjiang. Seed, 2018, 37(2): 109-110. (in Chinese)
[42] 盛万民, 张丽娟, 牛志敏, 李庆全, 解艳华, 南相日, 夏善勇, 钱创建. 马铃薯新品种龙薯4号的选育. 中国种业, 2020(3): 62-63.
SHENG W M, ZHANG L J, NIU Z M, LI Q Q, XIE Y H, NAN X R, XIA S Y, QIAN C J. Breeding of a new potato variety longshu No. 4. China Seed Industry, 2020(3): 62-63. (in Chinese)
[43] BERNAYS E A, GRAHAM M. On the evolution of host specificity in phytophagous arthropods. Ecology, 1988, 69(4): 886-892.
[44] GARCIA-ROBLEDO C, HORVITZ C C. Parent-offspring conflicts “optimal bad motherhood” and the “mother knows best” principles in insect herbivores colonizing novel host plants. Ecology and Evolution, 2012, 2(7): 1446-1457.
[45] 闫俊杰, 张梦迪, 高玉林. 马铃薯块茎蛾生态学、生物学与综合治理. 昆虫学报, 2019, 62(12): 1469-1482.
YAN J J, ZHANG M D, GAO Y L. Biology, ecology and integrated management of the potato tuber moth,(Lepidoptera: Gelechiidae). Acta Entomologica Sinica, 2019, 62(12): 1469-1482. (in Chinese)
[46] HORGAN F G, QUIRING D T, LAGNAOUI A, SALAS A, PELLETIER Y. Periderm- and cortex-based resistance to tuber-feedingin two wild potato species. Entomologia Experimentalis et Applicata, 2007, 125(3): 249-258.
[47] FOOT M A. Susceptibility of twenty potato germplasms to the potato tuber moth at Pukekohe a preliminary assessment. New Zealand Journal of Experimental Agriculture, 1976, 4(2): 239-242.
[48] RAMAN K V, PALACIOS M. Screening potato for resistance topotato tuberworm. Journal of Economic Entomology, 1982, 75(1): 47-49.
[49] MUSMECI S, CICCOLI R, GIOIA V D, SONNINO A, ARNONE S. Leaf effects of wild species ofand interspecific hybrids on growth and behaviour of the potato tuber moth,Zeller. Potato Research, 1997, 40(4): 417-430.
The Adaptive Analysis ofto Different Potato Tuber Varieties
ZHANG MengDi1, YAN JunJie1, GAO YuLin1,2
1State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193;2National Center of Excellence for Tuber and Root Crop Research, Chinese Academy of Agricultural Sciences, Beijing 100081
【】Potato tuberworm ()is an important pest on potatoes, which seriously affects the quality and value of potatoes. The objective of this study is to clarify the adaptability ofon different potato tuber varieties, and to provide a basis for screening resistant potato varieties and developing effective integrated pest management strategy.【】To evaluate the adaptability ofon different tuber varieties and screen the high natural-resistance tubers, in this study, four varieties (Xiabodi, Longshu No. 4, Longshu No. 12 and Kexin No. 17) which were planted in a large scale at present were selected as tested materials to determine the development and population parameters ofon different tuber varieties using an age-stage specific two-sex life table.【】There were significant differences in larval stage, pupal stage and female longevity offeeding on different tuber varieties. Compared with those fed on Xiabodi, Longshu No. 4 and Longshu No. 12, those fed on Kexin No. 17 exhibited significantly longer larval stage (15.18 d) and pupal stage (8.10 d), and the female longevity (8.46 d) was shorter than the other three varieties. Moreover, the longevity of male was significantly higher than female in a same variety. The fecundity offeeding on Xiabodi (235.06 eggs) and Longshu No. 4 (254.48 eggs) was significantly larger than that fed on Kexin No. 17 (165.71 eggs). In addition, there were significant differences among intrinsic rate of increase (), finite rate of increase (), net reproductive rate (R) and mean generation time () offeeding on different tuber varieties. The,,Rvalues offed on Kexin No. 17 (=0.108,=1.125,R=26.513) were significantly lower than those on the other three tuber varieties, andvalue offed on Kexin No.17 (30.28) was significantly higher than those on the other three tuber varieties.Larvae feeding preference experimentsshowed that the larvae fed on Xiabodi and Longshu No. 4 in preference to Kexin No. 17. with the increase of age, the selectivity gradually decreased. 【】There was a different adaptability ofamong different potato tuber varieties. Xiabodi and Longshu No. 4 are the most suitable hosts, followed by Longshu No. 12, and Kexin No. 17 is the least suitable host. In areas whereis seriously damaged on potato field system, planting potato varieties with low adaptability can be selected as an important strategy for reducing population ofin the field.
potato tuberworm (); age-stage specific two-sex life table; potato variety; variety adaptability

10.3864/j.issn.0578-1752.2021.03.008
2020-07-27;
2020-08-25
国家重点研发计划(2018YFD0200802)
张梦迪,E-mail:sdaumengdizhang@163.com。通信作者高玉林,E-mail:gaoyulin@caas.cn
(责任编辑 岳梅)
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
中国种业(2022年7期)2022-07-21
体育科技文献通报(2022年5期)2022-06-05
今日农业(2021年15期)2021-11-26
信阳农林学院学报(2021年1期)2021-04-01
现代农业科技(2017年13期)2017-08-09
农业科技与装备(2017年1期)2017-06-09
现代园艺(2016年7期)2017-01-09
农村农业农民·B版(2016年7期)2016-10-21
现代园艺(2016年9期)2016-07-16
