
果糖在食欲调控中的作用机制
2021-03-26 03:01甘凯文于浩泳
医学综述 2021年5期
甘凯文,于浩泳
(上海交通大学附属第六人民医院内分泌代谢科,上海 200233)
果糖是一种单糖,为葡萄糖的同分异构体,主要以游离形态存在于水果和蜂蜜中。1970年果糖的生产技术取得突破,自此果糖以果葡糖浆为主要形式被大量生产,并广泛用于食品加工行业[1-2]。研究显示,美国人均果糖摄入量从1970年末的37 g/d增加至1999—2004年的49 g/d[1]。近年来果糖消费量和摄入量剧增,每日人均摄入糖量远超世界卫生组织指南的推荐量[3],全球范围内肥胖的发生率与果糖摄入量呈正相关[2]。研究表明,摄入果糖可导致食欲亢进,引起2型糖尿病、胰岛素抵抗、高脂血症以及神经损伤等[4-6],与单纯添加葡萄糖的等热量饮食相比,果糖更易导致肥胖和内脏脂肪蓄积,降低饱腹感,从而导致更严重的代谢紊乱[7]。
中枢神经系统(central nervous system,CNS)和外周组织产生的食欲相关信号联合作用调控食欲。在CNS中,下丘脑起主导作用,与弓状核、腹内侧核、背中核、室旁核、外侧下丘脑等共同参与食欲信号的合成、释放及调控不同区域的食欲信号[8];外周的食欲信号通路始于受摄食刺激分泌的胃肠道激素,然后通过不同神经通路传入中枢,并通过中枢调节摄食行为[9]。现就果糖在食欲调控中的作用机制进行综述。
1 果糖对中枢食欲信号的调控
1.1果糖与下丘脑食欲调节神经肽 下丘脑作为摄食中枢在能量代谢调控起主导作用,可合成食欲调节相关的神经肽,包括促食欲神经肽[神经肽Y、刺鼠基因相关蛋白(agouti-related protein,AgRP)、黑色素浓集激素、增食欲素]和抑制食欲的神经肽[阿片黑素促皮质素原(proopiomelanocortin,POMC)、可卡因-苯丙胺调节转录肽][5,8]。研究发现,接受高果糖饮食或脑室内注射果糖溶液的小鼠进食量和体重均有所增加,并观察到POMC信使RNA(messenger RNA,mRNA)的表达水平下降[10-12]。关于果糖对神经肽Y和AgRP表达的影响存在争议,可能与各研究中果糖剂量及观察时长不同有关。长期仅给予果糖的间歇性摄食大鼠模型中观察到伏隔核内增食欲素神经元活性下降和外侧核增食欲素神经元活性增强,增食欲素神经元激活机制可能与果糖摄入引起外周循环三酰甘油水平显著升高相关[13],表明长期果糖摄入可降低伏隔核中γ-羟基丁酸能神经元对外侧下丘脑和腹侧苍白球的输出,激活外侧下丘脑内增食欲素神经元释放增食欲素-A触发摄食效应,导致果糖摄入持续增加,形成正反馈循环[12]。
1.2与果糖代谢途径相关的AMP活化蛋白激酶(AMP-activated protein kinase,AMPK)/丙二酰辅酶A(malonyl-coenzyme A,MCoA)信号通路 果糖通过与肠黏膜上皮的葡萄糖转运体结合进入血液循环,并运输至肝脏进行代谢,葡萄糖转运体5是果糖唯一的特异性转运体,不受胰岛素影响,主要存在于小肠、肌肉组织、海马体、大脑皮质小胶质细胞以及小脑浦肯野细胞中[14-15]。研究指出,高果糖饮食喂养小鼠的小肠和中枢葡萄糖转运体5 mRNA的表达上调,影响大脑能量代谢稳态[16-17]。虽然葡萄糖和果糖均通过糖酵解途径代谢,但在肝脏代谢的起始步骤不同,果糖绕过糖酵解中的限速步骤自主代谢,由不依赖胰岛素的特异性果糖激酶催化果糖磷酸化生成1-磷酸果糖。在中枢果糖绕过限速步骤会引起ATP的迅速消耗及AMP的代偿性增多,造成下丘脑 AMP/ATP比值升高,进而激活AMPK使乙酰辅酶A羧化酶磷酸化失活,减少其催化产物MCoA的生成[6]。AMPK的激活受激素水平和营养状态的调节,如胃饥饿素和低血糖,中枢AMPK激活刺激摄食,而瘦素、胰岛素和葡萄糖则抑制摄食[18-19]。MCoA是下丘脑信号级联中调节高等动物能量平衡的关键中间体,通过下丘脑食欲调节神经肽系统调控食欲信号和摄食行为[6]。MCoA的升高可激活弓状核神经元,降低神经肽Y和AgRP的表达,并上调α-促黑素(POMC的蛋白水解产物)和可卡因-苯丙胺调节转录肽的表达,最终导致食欲及进食量下降[10]。相反,果糖可导致下丘脑MCoA水平下降[6,18],食欲亢进、摄食增加及肥胖[6](图1)。果糖对血脑屏障通透性的作用尚未明确,曾有学者质疑膳食果糖直接进入CNS影响食欲的学说,但研究证实小鼠大脑的多个区域存在果糖代谢所需的酶类及果糖转运蛋白,同时葡萄糖转运体5也在血脑屏障中表达[20]。其他研究表明,外周静脉注射果糖后下丘脑细胞外液中果糖水平增加,支持果糖直接经血脑屏障进入CNS进行代谢,而不需经肝脏转化为葡萄糖后再进入CNS代谢的结论[16,21]。
1.3果糖与内源性大麻素系统(endocannabinoid system,ECS) 生物体自身可产生大麻素,称为内源性大麻素(endocannabinoid,EC),是一类神经递质分子,主要包括N-花生四烯酸氨基乙醇和2-花生四烯酸甘油[22]。N-花生四烯酸氨基乙醇主要是在N-酰基磷脂酰乙醇胺磷脂酶D和磷脂酶C的催化下合成,二酰基甘油脂肪酶α和β主要参与2-花生四烯酸甘油的合成,可分别被脂肪酸酰胺水解酶和单酰基甘油酯酶降解灭活,EC、与EC结合的受体以及其合成和降解酶类共同组成ECS[23]。大麻素具有促进食欲的作用。多项研究表明,ECS在食欲调节、脂肪合成、胰岛素抵抗、能量代谢及摄食等方面发挥重要作用[23-24]。EC通过与CNS突触前神经末梢表达的大麻素受体1(cannabinoid 1 receptor,CB1R)结合促进食欲[4,24-25]。目前已证实选择性CB1R拮抗剂利莫那班可通过作用于CNS及外周组织的CB1R达到有效降低食欲和减重的目的,特别是降低对甜食、饮料及可口食物的摄食欲望,而CB1R激动剂则起相反作用[4,26]。研究表明,果糖喂养的小鼠下丘脑CB1R mRNA水平显著升高,而葡萄糖喂养的小鼠则未观察到明显的CB1R mRNA水平上升[4,25]。另一项研究指出,果糖的摄入可影响EC的合成和降解酶系统,在果糖喂养的小鼠中可观察到脂肪酸酰胺水解酶、N-酰基磷脂酰乙醇胺磷脂酶D、二酰基甘油脂肪酶β mRNA表达升高,而磷脂酶C、单酰基甘油酯酶、二酰基甘油脂肪酶α mRNA表达下降,提示果糖较其他糖类(蔗糖和葡萄糖)对ECS的影响更大[4],但该研究并未对相关基因及EC蛋白的表达水平进行分析,仅根据EC相关酶系统的基因转录水平变化难以直接评价果糖对ECS的实际影响。此外,ECS除直接通过中枢效应调节食欲,还可通过与脑内调节食欲的相关通路(如奖赏回路、5-羟色胺系统、增食欲素-A信号通路、内源性阿片肽)相互联系,共同发挥调节食欲和摄食行为的功能[24,27]。
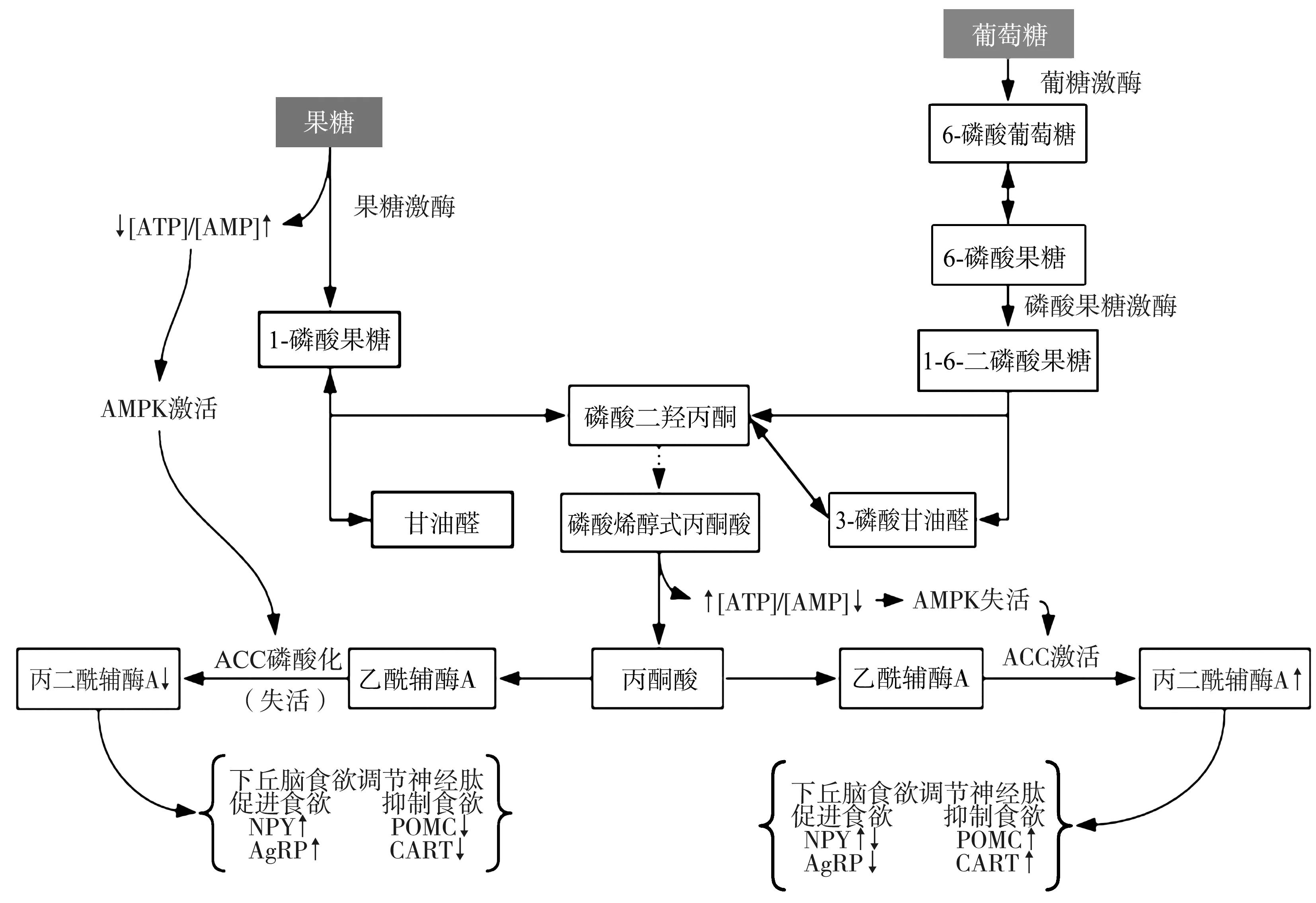
ATP:腺苷三磷酸;AMP:腺苷一磷酸;AMPK:AMP活化蛋白激酶;ACC:乙酰辅酶A羧化酶;NPY:神经肽Y;AgRP:刺鼠基因相关蛋白;POMC:阿片黑素促皮质素原;CART:可卡因-苯丙胺调节转录肽;MCoA:丙二酰辅酶A
1.4果糖对大脑区域活动的影响 下丘脑为食欲调节中枢,其与参与调节进食动机及奖赏系统的大脑其他区域(包括纹状体、眼窝前额皮质、杏仁体和脑岛)共同组成综合网络调控食欲[8]。有研究采用功能性磁共振成像技术以局部脑血流量作为神经元激活的间接指标,探讨大脑区域活动与果糖介导的摄食行为改变的关联[21,28]。正常生理状态下,机体从饥饿状态转为饱腹状态时,下丘脑及纹状体活动减弱,但摄入果糖后,人类丘脑、海马、后扣带皮质、梭状回和视觉皮质的脑血流量显著减少,但并未降低下丘脑活动,反而引起下丘脑活动短暂且小幅度的升高,同时下丘脑与纹状体功能连接未见增强[21]。此外,海马体的记忆过程也参与了进食行为的调节,海马体损伤的啮齿动物可失去识别饥饿和饱足状态的能力,导致食欲亢进[29]。人类海马体的激活与食物渴求、饥饿状态等有关,提示摄入果糖后海马体活动减少可能诱导食欲亢进的发生[21]。另一方面,果糖与享乐摄食及奖赏机制有关,相对于葡萄糖,果糖摄入导致大脑视觉皮质和左眼眶额叶皮质对食物线索的反应性增强,表明果糖增强了对食物线索的奖赏和动机信号的传递,并刺激大脑中负责处理注意力和奖赏的区域,最终表现为对食物强烈的渴望和饥饿感[30]。
2 果糖对外周食欲信号的影响
摄食行为刺激胃肠分泌细胞分泌约20种多肽激素,部分激素与CNS相互作用组成脑-肠轴调控摄食行为及能量稳态,这些胃肠激素被称为脑肠肽,分为促食欲因子(胃饥饿素)和饱腹因子[瘦素、胰高血糖素样肽1(glucagon-like peptide-1,GLP-1)]两类,除脑肠肽外,胰岛素也在人体代谢和饮食行为中起重要作用。
2.1胃饥饿素 胃饥饿素由胃肠的内分泌细胞和大脑弓状核分泌,是唯一在外周刺激食欲的胃肠激素,与短期饥饿、食物摄入和体重有关,与血糖、胰岛素及瘦素水平呈负相关[8,31]。胃饥饿素调节进食和能量稳态依赖于其与生长激素促分泌素受体的结合,生长激素促分泌素受体广泛分布于脂肪组织、肾上腺、生殖腺、胰腺、胃以及CNS(如下丘脑、腹侧被盖区、海马体、大脑皮质、背侧迷走神经复合体)[32]。下丘脑内生长激素促分泌素受体的激活可直接提高促食欲神经元的活性,引起突触连接方式的改变,从而促进食欲回路的激活,同时抑制促进饱腹感的通路[33]。研究指出,摄入果糖后引起外周循环中胃饥饿素水平升高可导致食欲亢进[34-35],这可能与外周胃饥饿素穿过血脑屏障作用于多个大脑回路有关。胃饥饿素可直接激活弓状核内神经肽Y/AgRP及外侧下丘脑内增食欲素神经元,并抑制POMC神经元[36];还可通过激活Gq/11蛋白介导的磷脂酶C/三磷酸肌醇信号通路及Gs蛋白介导的环磷酸/蛋白激酶A信号通路激活AMPK[32,37],磷脂酶C/三磷酸肌醇信号通路及AMPK自身可促进细胞内钙水平升高,有利于2-花生四烯酸甘油生成,EC可间接激活下丘脑AMPK,AMPK的激活(磷酸化)抑制乙酰辅酶A羧化酶和脂肪酸合酶,介导细胞质内MCoA和长链脂肪酰辅酶A的减少,促进下丘脑内脂肪酸氧化[33,38]。AMPK介导的脂肪酸氧化过程中,线粒体内解偶联蛋白-2水平上调,神经肽Y和AgRP的转录水平升高,最终驱动摄食行为[39]。另外,胃饥饿素可作用于边缘奖赏系统,亦可同时作用于突触前和突触后的生长激素促分泌素受体以激活多巴胺神经元[33],在动物外周或脑室内注射胃饥饿素均能引起腹侧被盖区及伏隔核中多巴胺水平升高[31],可观察到动物摄食量增加,以及更倾向选择高热量的食物[40]。正常体重人群进行享乐性进食时血浆中胃饥饿素和2-花生四烯酸甘油水平升高,提示胃饥饿素与ECS在奖赏相关反应的调节中存在联系[24]。胃饥饿素亦可激活人体边缘奖赏系统的其他区域(如杏仁核、眼窝额叶皮质和纹状体),增加进食快感及摄食渴求[8-9]。
2.2胰岛素 中脑边缘多巴胺奖赏回路主要由腹侧被盖区、伏隔核、杏仁核、海马等构成,腹侧被盖区是奖赏索求和抑制摄食的关键部位[41]。已知胰岛素作用于CNS内相应受体可产生饱腹感,并削弱边缘奖赏系统及大脑皮质对食物刺激的反应[5,21],其机制为胰岛素直接抑制腹侧被盖区中多巴胺能神经元的活性,或间接通过EC介导的长时程抑制多巴胺能神经元上的兴奋性突触传递[22],还可通过抑制下丘脑食欲调节神经肽的表达[5,8],实现饱腹效应。研究发现,与普通饮食以及加用葡萄糖饮料的动物和人类的胰岛素水平相比,加用果糖饮料胰岛素水平更低[11-12,35],且血糖水平无明显上升[42],摄入果糖不引起外周胰岛素水平上升可能与胰岛β细胞内低表达葡萄糖转运体5有关,认为果糖基本不刺激胰岛素分泌[35],并指出长期果糖摄入可引起胰岛素抵抗及下丘脑胰岛素信号紊乱,抑制CNS内胰岛素信号转导,从而降低饱腹感[8]。
2.3瘦素 瘦素是脂肪组织分泌的抑制食欲性激素,通过瘦素受体调控下丘脑内多个与食欲相关的系统。瘦素可通过激活乙酰辅酶A羧化酶、降低AMPK活性增加弓状核内MCoA的合成,通过AMPK/MCoA信号通路调节下丘脑神经肽的分泌和表达[8,43-44];瘦素是下丘脑EC水平的强调节剂,通过降低细胞内钙水平抑制EC合成和信号转导,间接影响下丘脑神经肽的表达[24]。在下丘脑CB1R部分敲除的小鼠中发现,瘦素介导的抑制摄食的作用减弱,提示瘦素通过下丘脑CB1R信号抑制食欲[24]。瘦素可直接介导下丘脑食欲神经肽的表达,直接刺激弓状核中的POMC神经元,促进蛋白水解产物α-黑色素细胞刺激素释放,抑制AgRP及神经肽Y分泌,还可刺激邻近POMC神经元释放阿片类物质以抑制外周释放胃饥饿素[8]。果糖对瘦素水平影响的研究结果并不一致,可能与瘦素水平受体脂分布与体脂含量影响有关[4,45]。研究发现,长期喂食果糖的小鼠出现瘦素敏感性下降及瘦素抵抗,并与果糖饮食引起的高三酰甘油血症有关,三酰甘油可导致瘦素血脑转运障碍、瘦素受体基因表达下降及细胞因子信号转导抑制剂基因表达增加[11]。外周循环中瘦素水平下降可刺激AgRP及神经肽Y的表达,抑制POMC和可卡因-苯丙胺调节转录肽表达[46],而神经肽Y表达增加与瘦素抵抗直接相关[11]。故果糖介导外周瘦素水平下降以及瘦素抵抗,影响CNS内瘦素相关信号转导,从而削弱了瘦素介导的饱腹效应。
2.4GLP-1 GLP-1是肠道L细胞和孤束核分泌的一种短效饱腹肽类激素,与胰岛β细胞中GLP-1受体结合刺激胰岛素分泌,同时具有延迟胃排空、抑制食欲和摄食等效应[47]。其中,GLP-1的抑制食欲作用是最早发现的胰外效应,GLP-1受体激活介导食欲抑制及摄食量降低效应主要是通过抑制下丘脑的活性实现[18],此外也可通过谷氨酸受体α-氨基-3-羟基-5-甲基-4-异唑丙酸/红藻氨酸介导的多巴胺能神经元突触兴奋性增加实现[48]。果糖可降低餐后GLP-1的上升幅度,延迟GLP-1释放[21,35],削弱GLP-1激活对AMPK活性的抑制作用[18],从而降低GLP-1介导的饱腹效应。
3 展 望
果糖不仅作用于下丘脑的食欲调节相关信号通路,还可通过介导大脑不同区域神经元的活动实现促食欲效应,通过研究果糖对CNS和外周胃肠激素形成的复杂食欲调节网络的影响,有利于理解果糖在分子学层面与摄食行为改变之间的关系。目前关于果糖在调控食欲方面的研究较少,对于予以果糖后各种食欲调节相关因子及下丘脑神经肽基因水平的变化尚未完全了解,目前研究方向集中在糖类对食欲调节相关通路的酶类及相应受体、转运蛋白受体、糖代谢相关酶类基因水平的影响,但基因表达水平上调并不一定意味着编码蛋白质表达水平的增加,为准确判断具体基因或编码产物调节食欲的机制,应进一步探讨果糖摄入后食欲调节相关通路基因在蛋白转录水平的变化;同时对于食欲因子在食欲调控机制中的作用亦需进一步研究。
猜你喜欢
癌症进展(2022年18期)2022-11-26
四川蚕业(2022年2期)2022-11-19
特种经济动植物(2022年2期)2022-02-15
四川蚕业(2021年2期)2021-03-09
中老年健康(2017年9期)2017-12-13
江苏农业科学(2017年17期)2017-11-15
中国医药导报(2016年34期)2017-06-05
湖北农业科学(2017年1期)2017-03-09
校园英语·下旬(2016年4期)2016-05-09
中学科技(2014年11期)2014-12-25
