
土壤镉胁迫对檫木光合特性的影响
2021-04-19 08:35赵弘益管珏镧张雪媛胡红玲
植物研究 2021年4期
赵弘益 管珏镧 张雪媛 梁 清 张 健 胡红玲*
(1. 四川农业大学林学院,生态林业研究所,成都 611130;2. 四川农业大学林学院,成都 611130)
近年来,由于工业化及城市的快速发展,环境污染和生态破坏问题日益严重,其中,土壤重金属污染已成为当下全球范围内突出的环境问题之一。镉(Cd)是一种银白色的有毒金属元素,由于其本身难以降解和较强的隐蔽性,是目前最具威胁的重金属元素之一[1~2]。土壤中的Cd 可被植物根系吸收并在植物体内积累,当植物体内积累的Cd 浓度达到一定水平,会对植物产生毒害作用[3]。光合作用是植物物质生产重要的生理生化过程,相关研究表明,植物的光合生理特性在一定程度上能体现植物对生境的响应情况[4~5],且光合作用对重金属离子的作用较为敏感[6]。土壤中重金属Cd 可能通过不同机制影响植物的光合作用。相关研究指出,当土壤中Cd含量为100 mg·kg-1时,植物的光合能力及生物量的积累受到显著抑制[7]。过量的Cd 会损害植物的光合机制,抑制光合作用,Dalla等[8]研究表明Cd 胁迫会破坏植物的叶绿体内囊体膜结构,从而抑制叶绿体的增加[9]。光合参数能表征植物的光合特性,通常可由光合—光响应曲线拟合得出[10~11],获得的光合参数可用于判断植物光合机构的状态以及反映叶片利用光、CO2等的能力[12]。
檫木(Sassafras tzumu Hemsl.)属于樟科(Lau⁃raceae)檫木属(Sassafras)落叶高大乔木,是四川珍贵的乡土用材树种,材质优良,常用于造船及上等家具;树形优美,秋叶变红,同时也是优良的园林观赏树种[13~14]。因此国内大多对檫木的研究集中在遗传选育与繁殖技术、栽培与造林技术、病虫害防治等方面[15~17],而有关檫木在重金属胁迫下的生长生理状况鲜有研究报道。鉴于此,本研究通过研究Cd胁迫对檫木光合特性的影响,分析檫木光合机制对Cd胁迫的响应,旨在为进一步研究檫木对重金属Cd的耐受性及富集策略提供一定的理论依据。
1 材料与方法
1.1 试验地概况
试验地位于四川农业大学(成都校区)教学科研 试 验 基 地,地 理 位 置 为103°51′29″E,30°42′18″N,属于亚热带湿润气候区,四季分明,气候温和,四季雨量充沛。年平均气温约为16.8°C,月平均最高气温为25.9°C(7 月),月平均最低气温为6.9°C(12月),极端最高气温为38°C(7月),极端最低气温为-2°C(1 月);年平均相对湿度为84%,全年平均降水量为759.10~1 155.00 mm;年平均日照时数为840.2 h。
1.2 试验材料及试验设计
1.2.1 试验材料
供试植物为四川地区常见的乡土树种檫木1年生幼苗。取自同一种源,无病虫害。供试土壤为黄壤,取自四川省雅安市洪雅县林场,土壤pH为5.75,N 为9.10 g·kg-1,P 为0.64 g·kg-1,K 为12.59 g·kg-1。
1.2.2 试验设计
本试验采取盆栽控制试验的方法,称取约12 kg 混匀后的供试土壤装入花盆中,并根据土壤干重计算各组花盆土壤中Cd2+含量(mg·kg-1),其中Cd2+以CdCl2形式水溶施入。采用单因素试验设计,共设置5 个镉处理水平:CK(0 mg·kg-1),T1(5 mg·kg-1),T2(20 mg·kg-1),T3(50 mg·kg-1),T4(100 mg·kg-1)。以上各处理组均设置5 个重复,且所有处理植株均放置在大棚内培养,试验于2019年5 月初开始,处理前测定植株的初始株高,分5次施入CdCl2,每间隔15 d 施入一次。施入时,将配置的溶液均匀浇在盆内土壤表面,另用托盘垫在盆底收集可能渗出的CdCl2溶液,并将其浇回土壤中,CK 处理浇入等量的纯水。于2019 年7 月中旬结束施入CdCl2,待植株生长30 d后,于8月中旬开始,选择晴天的上午9:00~11:00 测定植株叶片的各光合参数。
1.3 测定指标及方法
1.3.1 叶绿素含量测定
随机选取各处理檫木植株健康成熟的功能叶片2~3 片进行测定,测定时将叶片去除叶中脉并剪碎后,准确称取0.1 g,装入10 mL 离心管中,加入9 mL 叶绿素提取液(80%丙酮与无水乙醇1∶1混合),放入暗处,静置24 h 以上直至叶片完全变白,在663和646 nm波长下进行比色。
各叶绿素计算公式如下:
式中:A663和A646分别为叶绿素提取液在紫外分光光度在波长663和646 nm处的吸光光度值。
1.3.2 光合气体交换参数测定
选择晴天,于上午9:00~11:00,对各处理植株树冠中上部选取健康成熟的功能叶片,采用Li-6800 便携式光合测定仪(Li-Cor Inc.,USA)进行测定。设定光合有效辐射为(PAR)1 200µmol˙m-2˙s-1,CO2浓度为400µmol˙mol-1,温度为30℃,相对湿度为45%~65%,测定各处理植株净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci),每个处理选择3 株,每株选择3 片叶子,每片叶片记录10个数据。测定叶片做标记。
1.3.3 光合-光响应特性的测定
选取相同部位叶片,采用Li-6800便携式光合测定仪测定各处理叶片光合—光响应曲线,光合有效辐射(PAR)梯度设定值为1 800、1 600、1 200、1 000、800、600、400、200、100、75、50、25、0 µmol˙m-2˙s-1,CO2浓度设定为400 µmol˙mol-1,叶室温度设定为30℃。测定时保持叶片在植株的原方位上,测定前对每个叶片进行20 min 的光诱导,本试验光诱导强度为800µmol˙m-2˙s-1。每个处理选择3株,每株选择3片叶子记录测定数据。
利用叶子飘直线双曲线修正模型对不同浓度镉处理下檫木叶片光合—光响应曲线进行拟合。直线双曲线修正模型拟合方程为:
式中:Pn为净光合速率(µmol˙m-2˙s-1);Pnmax为光饱和时的最大净光合作用速率(µmol˙m-2˙s-1);α为初始量子效率;I为光合有效辐射(µmol˙m-2˙s-1);Rd为暗呼吸速率(µmol˙m-2˙s-1);β 为光抑制系数,是修正系数(m2˙s˙µmol-1);γ 为光饱和系数(m2˙s˙µmol-1),是植物叶片光响应曲线的初始斜率与植物最大净光合速率之比。
当Pn=0时,可得光补偿点Ic:
1.4 数据处理
采用Excel2016 和SPSS24.0 软件对数据进行统计分析,采用单因素(one-way ANOVA)和LSD法进行方差分析和多重比较(α=0.05)。利用Ori⁃gin8.5软件作图,图中数据为平均值±标准误差。
2 结果与分析
2.1 镉胁迫下檫木叶片叶绿素含量
由图1 可知,随着镉处理浓度增加,檫木叶片叶绿素含量先增加后下降,且均在T2 处理下达到最大值。叶绿素a 含量和叶绿素总量在T1、T2、T3处理下与CK 相比并无显著性差异,T4 处理较CK差异显著。叶绿素b含量在T1、T2处理较CK 无显著差异,T3、T4处理较CK差异显著。
2.2 镉胁迫下檫木光合气体交换参数的变化
由图2可知,不同浓度镉处理仅对叶片的净光合速率(Pn)影响较大,随着镉处理浓度增大,檫木叶片的净光合速率(Pn)呈现下降趋势,T1 处理较CK 差异不显著,T2、T3 和T4 处理下,净光合速率(Pn)较CK 差异显著。不同浓度镉处理对气孔导度(Gs)、胞间二氧化碳浓度(Ci)和蒸腾速率(Tr)并没有产生明显影响,各处理间没有显著差异。
2.3镉胁迫下光合—光响应曲线的变化
随着镉处理浓度的增加,檫木叶片光合—光响应曲线变化如图3 所示。当光合有效辐射较弱时(PAR≤200 µmol˙m-2˙s-1),叶片净光合速率几乎呈现线性增加趋势,且CK 的净光合速率增幅显著高于其他镉处理浓度,T1、T2、T3、T4 处理下光合速率增幅随镉处理浓度增加而依次下降。当光合有效辐射超过200 µmol˙m-2˙s-1,不同镉浓度处理条件下檫木叶片净光合速率增加趋势减缓,当光合有效辐射在800~1 800 µmol˙m-2˙s-1时,檫木叶片净光合速率变化趋于平缓,最终达到光饱和点后,叶片的光合作用出现了光抑制现象。
根据叶子飘模型,利用直角双曲线修正模型拟合不同镉浓度处理下檫木叶片的光合—光响应曲线,并计算其光响应特征参数(见表1)。由表1可知,随着镉处理浓度增大,各处理条件下檫木叶片最大净光合速率(Pnmax)显著降低,CK的最大净光合速率(Pnmax)最大,为9.19µmol˙m-2˙s-1,T1、T2、T3和T4 处理下与CK 相比分别下降了37.11%、50.38%、80.20%和84.22%。不同镉浓度处理下檫木叶片的暗呼吸速率(Rd)与CK相比并无显著差异。
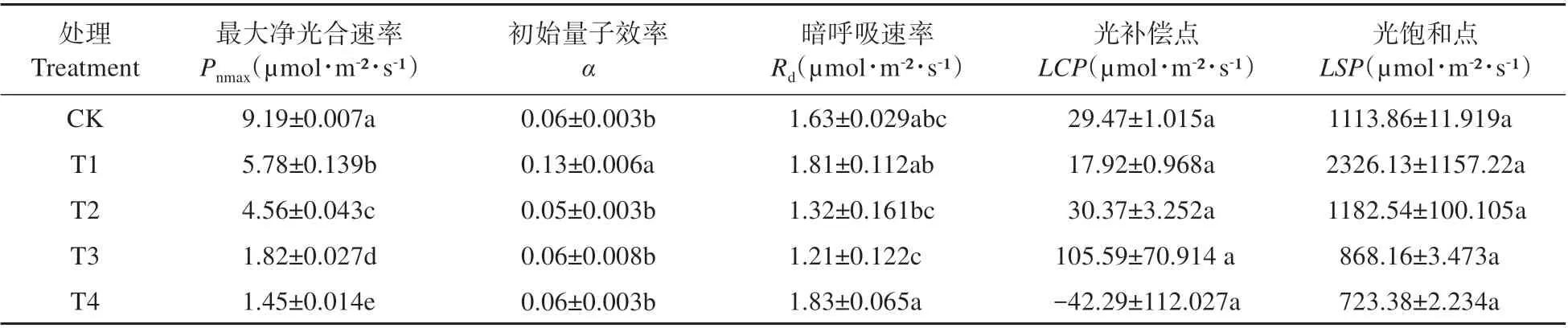
表1 光合-光响应参数Table 1 Photosynthetic light-response parameters
檫木叶片的光补偿点(LCP)在T3 处理时最大,在T4 处理时最小,但各处理间无显著差异;檫木叶片的光饱和点(LSP)在T1 处理时最大,在T4处理时最小,各处理间也无显著差异。5个浓度镉处理条件下檫木叶片光合—光响应曲线拟合方程的决定系数R2范围为0.94~0.99,除T2、T3 和T4 未达0.99以外,CK和T1处理的拟合曲线均具有较高拟合度。
3 讨论
光合作用是植物重要的生理过程,大量研究表明,重金属镉胁迫能抑制植物光合作用,且与重金属胁迫浓度显著相关[18~19],其对植物光合作用的影响主要体现在破坏叶绿素结构、降低光合色素含量等方面,而光合色素含量则能影响植物光合作用的能力[20]。叶绿素含量与光合作用强弱直接相关,可以反映叶片对光能的吸收和转化能力。作为参与光合作用的主要色素,叶绿素a在光反应中心负责将光能转化为化学能,叶绿素b则负责捕获和传递光能[21]。在不同浓度Cd 胁迫下,檫木叶片叶绿素含量先增加后下降,说明低浓度Cd 促进了叶绿素含量增加,而高浓度Cd 抑制叶绿素形成,这可能是由于低浓度Cd 的络合物会使植物加速从土壤中吸取Mg、Fe、K、P 等离子,促进叶片卟琳环形成,进而促使叶绿素含量增加[22];高浓度Cd处理破坏叶绿素体结构和生理活性,抑制了光合色素的形成,从而使叶绿素含量降低[23]。
研究表明,随着重金属浓度升高,植物的净光合速率一般呈现下降趋势[24],与本试验研究结果一致。影响植物叶片光合速率的因素可分为气孔因素和非气孔因素,Farquhar 等[25]认为,判断影响植物光合速率的因素,要同时观测气孔导度的大小和胞间二氧化碳浓度的变化,若光合速率下降时,两者同时下降则表明光合速率主要受气孔因素限制,若气孔导度下降而胞间二氧化碳浓度升高则主要受非气孔因素限制。本试验中,檫木叶片的净光合速率、气孔导度下降,胞间二氧化碳浓度升高,表明檫木叶片光合速率受非气孔因素限制。
光合—光响应参数能反映植物在逆境条件下的光合潜能、光能利用率及光抑制水平等[26~27]。Pnmax能体现植物潜在光合能力,其值越大则植物叶片的光合潜能越大,表明叶片能在有效的光照条件下合成更多的光合产物[28]。本研究中,随着Cd胁迫浓度升高,檫木叶片的Pnmax呈现显著下降趋势,表明重金属胁迫显著影响檫木叶片的光合能力。α(初始量子效率)能反映植物对光能的利用效率,可表征植物在弱光条件下捕获光量子用于光合作用的能力[29];α 越大,则表明植物利用弱光的能力越强,植物吸收与转换光能的色素蛋白复合体可能越多[30]。T1 处理下,檫木叶片的α 值最大,表明此时檫木叶片对弱光利用率的能力最强,可能由于低浓度Cd 处理促使叶绿素含量增加,从而提高了植物叶片光合作用的能力。相关研究指出,植物一般在适宜生长环境下的α 值为0.04~0.07[31],本试验中檫木叶片在不同Cd 浓度处理下的α 值为0.05~0.13,与该研究结果不一致。LCP(光补偿点)、LSP(光饱和点)分别反映植物利用弱光和强光的能力[32],本研究中,随着Cd 浓度处理的增加,檫木叶片的光补偿点和光饱和点总体呈现下降趋势,表明重金属Cd 胁迫减弱了檫木叶片对弱光、强光的利用能力。齐欣等[33]研究表明,降低LCP 是植物对低光环境的适应反应,能有利于植物在低光强下保持碳平衡。叶片较低的暗呼吸速率(Rd)和光补偿点(LCP)更有利于减少植物叶片光合产物消耗,进而增加净光合积累以获得最大碳积累量[28],檫木叶片在重金属Cd 胁迫条件下通过降低暗呼吸速率及光补偿点以此维持叶片光合作用的正常运作,体现了檫木叶片光合作用对重金属Cd胁迫的响应机制。
综上所述,在不同浓度Cd 胁迫处理条件下,檫木叶绿素含量先增后减,在T1、T2 处理下高于CK,体现了Cd 胁迫“低促高抑”的效应;光合气体交换参数及光合—光响应参数较CK 总体上均呈现下降趋势,体现了Cd 胁迫对檫木光合作用的危害。本研究仅从光合特性方面探讨了Cd 胁迫对檫木叶片光合作用的影响,未能从生理特性和叶片结构等多方面来阐述Cd 胁迫对植物光合生理机制的影响,未来拟结合生理指标测定等多种手段进一步研究Cd 胁迫对檫木叶片光合作用的影响。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
阅读(科学探秘)(2020年8期)2020-11-06
农业与技术(2020年12期)2020-07-04
绿色科技(2019年2期)2019-05-21
新课程·下旬(2017年7期)2017-08-14
女性天地(2016年10期)2017-04-25
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
食品工业科技(2014年6期)2014-05-10
