
多花水仙种球膨大期对乙烯的响应研究
2021-04-22 07:37罗可欣郭志雄陈桂信佘文琴潘东明潘腾飞
核农学报 2021年6期
姜 琳 孙 兴 罗可欣 郭志雄,2 陈桂信,2 佘文琴,2 潘东明,2 潘腾飞,2,*
(1 福建农林大学园艺学院,福建 福州 350002;2 福建农林大学园艺产品贮运保鲜研究所,福建 福州 350002)
乙烯是一种重要的植物激素,广泛存在于高等植物中,对生长发育起重要作用[1]。细胞的分化如种子萌发[2]、鳞茎膨大[3]等过程均受乙烯的调控,尤其是对半水生植物,乙烯能够刺激叶片萌发[4],以上反应是乙烯通过信号传导途径进行响应的[5]。
乙烯响应因子(ethylene response factor,ERF)为植物特有转录因子,是乙烯传导途径中最下游元件[6],属AP2/ERF大家族中的其中一个亚家族[7]。AP2/ERF基因家族庞大,每种植物有100 种以上ERF 转录因子[8-9]。ERF作为乙烯信号传导途径的最后一个受体蛋白,能够激活目标基因的表达,通常参与生物及非生物胁迫的响应过程[10]。在众多的ERF转录因子中,只有少数参与乙烯介导的生长调控,表现出正调控和负调控功能[11-12]。吴凡等[13]在牡丹(Paeoniasuffruticosa)花瓣的转录组数据中筛选到3个ERF转录因子,并通过试验表明PsERF1参与乙烯响应途径,PsERF2和PsERF3却参与多种信号调控途径。
水仙[NarcissustazettaL. var.chinensis(M. Roem.)]是典型的夏休眠球根类花卉,3—5月为球根类花卉鳞茎膨大期,在此期间,种球迅速膨大,是营养生长期,为7月下旬的花芽分化积累养分[14-15]。研究表明,一定浓度的乙烯能够促进洋葱(Alliumcepa)种球的生长[16],糖和乙烯共同调控乙烯信号传导途径中CTR1转录因子的表达[17],但对ERF转录因子的调控尚未明确,乙烯对水仙种球的营养生长的作用也鲜见报道。为研究乙烯在水仙鳞茎膨大过程中的作用,本试验拟通过测定种球膨大期乙烯含量的变化,研究ERF基因的表达规律,探讨其与种球生长的关系,以期为进一步了解水仙种球的生长发育提供研究基础。
1 材料与方法
1.1 材料
以漳州市龙文区蔡坂村第二年栽培的漳州多花水仙种球为试验材料,种球露地栽培。取样时期从2018年3月14日至2018年5月13日,每隔10 d取样一次,分别选取鳞片,内芽的组织样品(取样部位如图1所示),用于乙烯释放量测定和基因表达量分析。所有样品均设置3个生物重复。
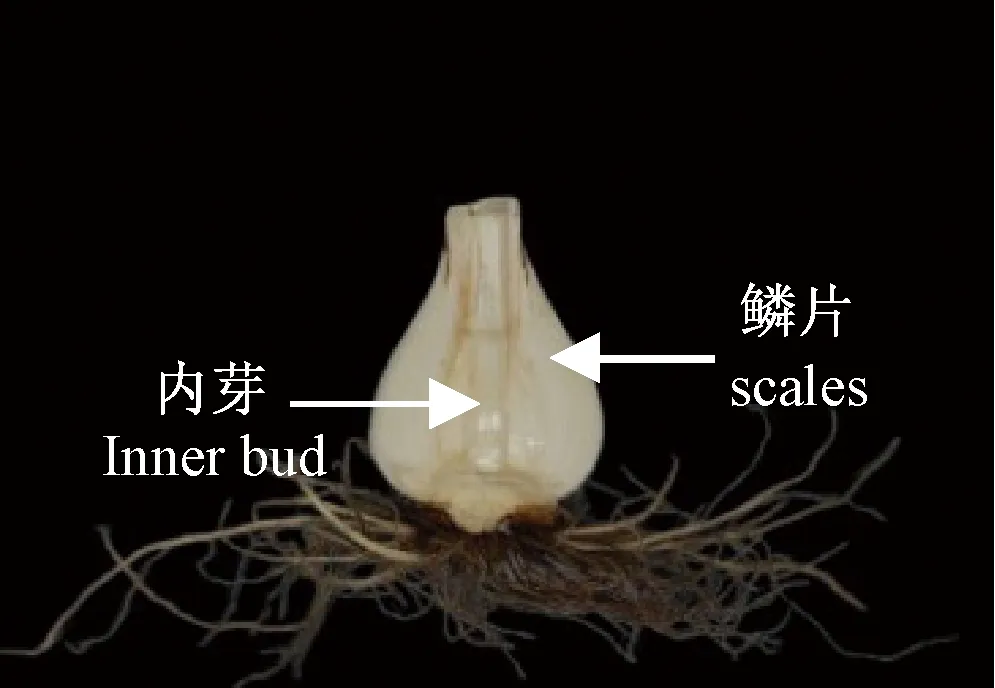
图1 多花水仙种球纵切图
从福建农林大学园艺产品贮藏保鲜研究所获得的水仙种球内芽和鳞片的转录组数据库中筛选得到7个在种球膨大期间有差异表达(FDR<0.01且Fold Change≥2)的NtERFs基因。
1.2 试验方法
1.2.1 总RNA提取与cDNA合成 多花水仙总RNA提取采用Trizol试剂法;cDNA合成采用PrimeScript RT reagent Kit试剂盒(大连TaKaRa公司),具体操作参照试剂盒说明书进行。
1.2.2 乙烯气体释放量测定 采用ETD-300乙烯测定仪(荷兰Sensor Sense公司)测定多花水仙乙烯释放量。乙烯检测仪通电预热60 min后,打开烃分解器预热5 min,分别将鳞片和内芽装进样品室后连接到阀门控制箱,阀门控制箱压强控制在0.1 MPa,选择积累测定模式,测定流速5 L·h-1。乙烯气体在样品室中积累30 min后测定乙烯释放量,每组处理重复测定3次。
1.2.3 实时荧光定量PCR 根据转录组数据中得到的NtERFs基因序列设计实时荧光定量PCR(quantitative real-time PCR, qRT-PCR)引物。采用Beacon Designer软件设计引物,验证基因可行性,引物序列见表1,选用actin为内参基因。
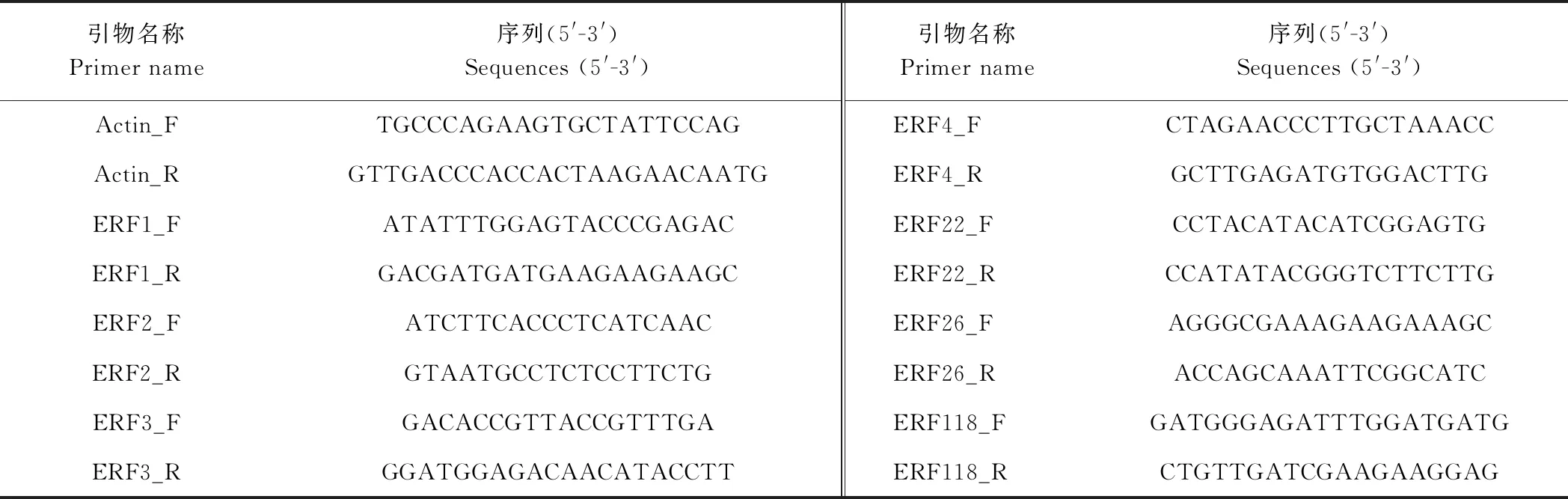
表1 ERFs基因qRT-PCR引物序列
1.2.4 qRT-PCR反应体系及程序 参照SYBR® Premix Ex TaqTM(Tli RNase plus)(大连TaKaRa公司)说明书,使用qtower 3.0定量PCR仪(德国耶拿公司)进行定量分析,PCR反应条件为PCR反应激活95℃预变性30 s,95℃ 变性5 s,60℃ 退火延伸30 s,40个循环。采用2-ΔΔCT的计算方法分析相对表达量,以第一个时期的表达量为对照。
1.2.5 数据分析 所有数据使用软件SPSS 21.0进行统计。采用Duncan检验进行样品间的差异显著性分析,用Excel制图。
2 结果与分析
2.1 水仙各部位乙烯释放量的变化
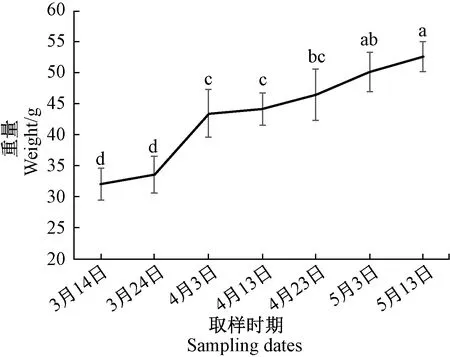
由图2可知,从3月份至5月下旬,水仙种球的重量逐渐增加,说明该时期为水仙的营养生长期,种球在不断增大。在整个膨大期,不同部位的乙烯释放量各不相同(图3)。水仙鳞片的乙烯释放量在3月下旬至5月上旬之间无显著差异,5月中旬乙烯释放量显著上升,释放量为0.39 nL·h-1·g-1。内芽释放的乙烯量明显高于鳞片,乙烯释放高峰出现在4月下旬(释放量为7.27 nL·h-1·g-1),比鳞片提前出现释放高峰,随后又呈现下降的趋势,到5月中旬略有上升的趋势。根据内芽乙烯的释放规律推测,内芽释放的乙烯可能与种球的膨大有关,且能够诱导鳞片乙烯的释放。在漳州地区,水仙种球的地上部在5月份出现黄化萎蔫,说明水仙种球鳞片乙烯释放与地上部的衰老有关。
注:不同小写字母表示不用时期的样品差异显著(P<0.05)。下同。
2.2 水仙膨大期鳞片NtERFs的表达规律分析
定量分析结果表明,水仙NtERFs基因在膨大期鳞片中的表达差异显著(图4)。其中,NtERF1、NtERF3、NtERF4、NtERF22和NtERF118的表达量在5月下旬显著高于其他时期,NtERF1、NtERF4和NtERF118的表达量在种球膨大期间呈现显著上升的趋势,而NtERF3和NtERF22则表现上升-下降-上升的变化趋势,分别在4月上旬和3月下旬出现第一次释放高峰。NtERF2的表达量表现为先上升后下降的变化趋势,在4月中旬出现表达高峰。NtERF26的表达量在3月份最高,而后下降,在4月下旬又显著升高。NtERF2和NtERF26的表达量均在4月23日内芽乙烯释放高峰期显著升高,这2个NtERFs可能是鳞片对内芽释放的乙烯的响应因子。从相对表达量变化来看,NtERF4和NtERF118的相对表达量从3月中旬到5月中旬分别升高2.2倍和2.4倍;NtERF1的相对表达量高于其他的NtERFs基因的相对表达量,5月中旬的相对表达量是3月中旬的5.1倍,且NtERF1表达与鳞片的乙烯释放规律一致,因此推测,NtERF1、NtERF4和NtERF118参与了鳞片乙烯信号的响应以及水仙种球的生长,NtERF1可能是关键的因子。
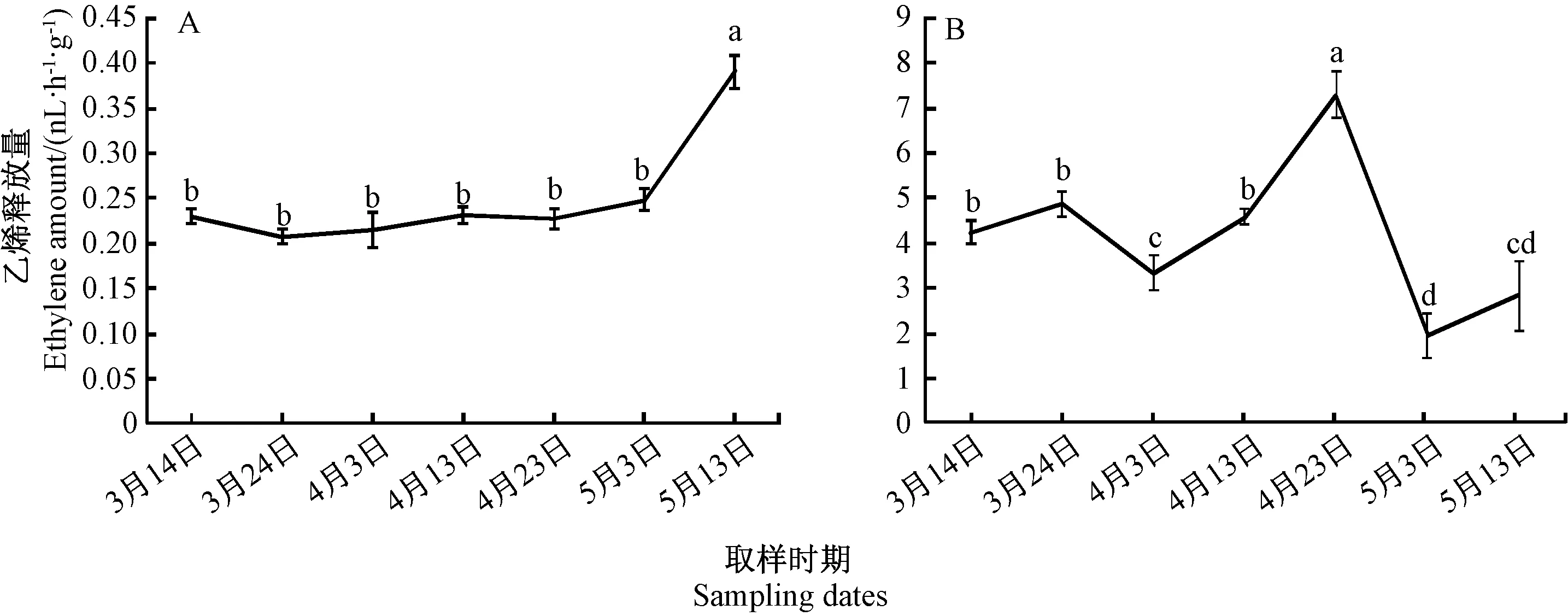
注:A: 水仙鳞片乙烯释放量;B:水仙内芽乙烯释放量。
2.3 水仙膨大期内芽NtERFs的表达规律分析
水仙种球内芽NtERFs的定量分析结果表明(图5),NtERF1基因在4月3日出现表达高峰,表达量是其他时期的2.5~9.0倍;NtERF2在4月23日的表达量是其他时期的7倍左右,其他时期的表达量维持在较低水平,且无显著性差异,NtERF2的表达高峰与内芽的第二次乙烯释放高峰期相吻合,因此推测NtERF2基因可能是参与内芽乙烯信号响应的重要因子。NtERF3和NtERF4表达规律与内芽乙烯的释放规律基本相符,说明NtERF3和NtERF4也可能参与了内芽对乙烯的响应;NtERF22的表达量在5月初显著高于其他时期,NtERF26和NtERF118的表达量在4月底显著升高,但这5个NtERFs的相对表达量较低,可能不是内芽乙烯释放和响应的主要因子。
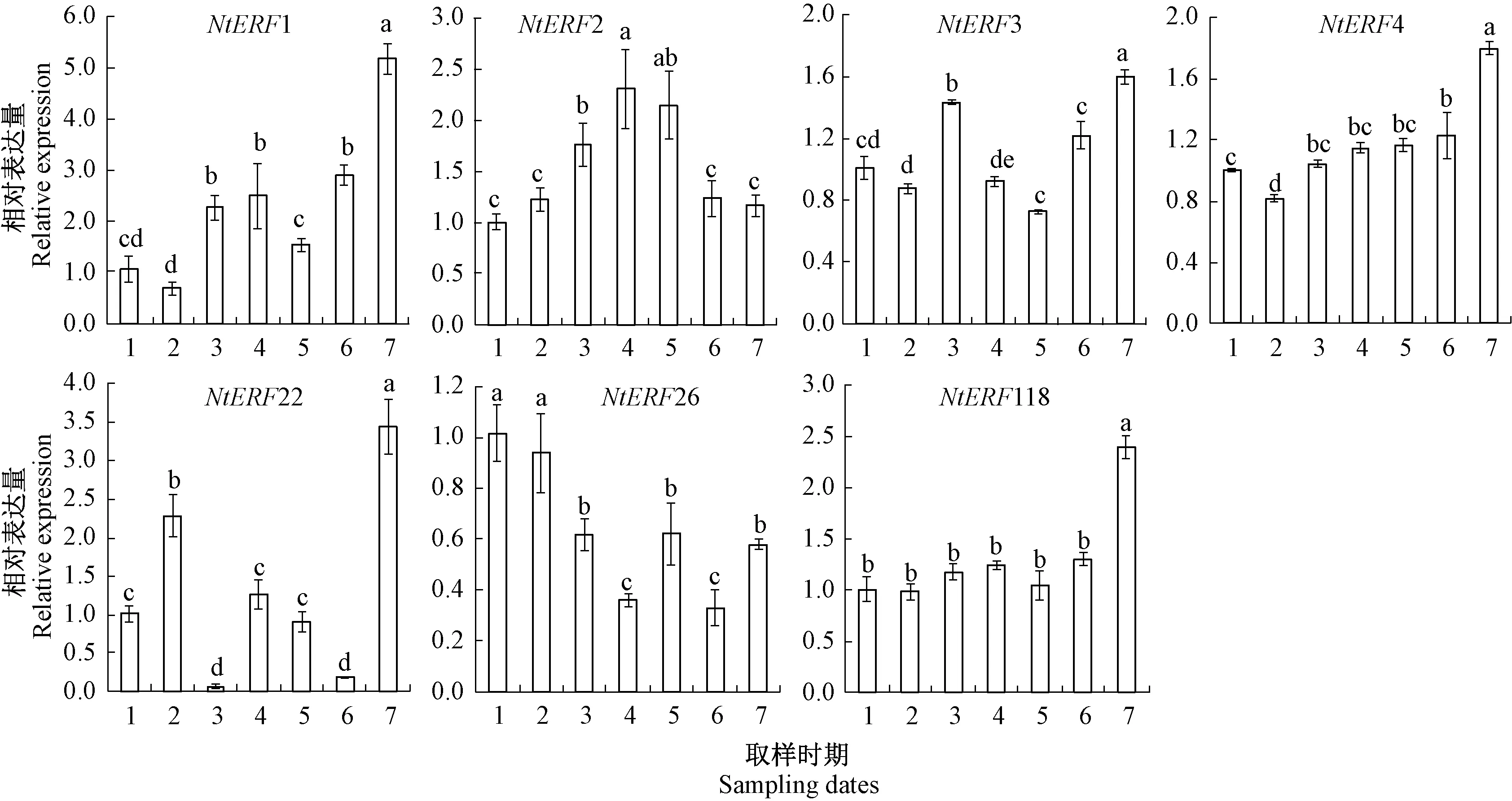
注:1~7分别表示3月14日、3月24日、4月3日、4月13日、4月23日、5月3日、5月13日。下同。
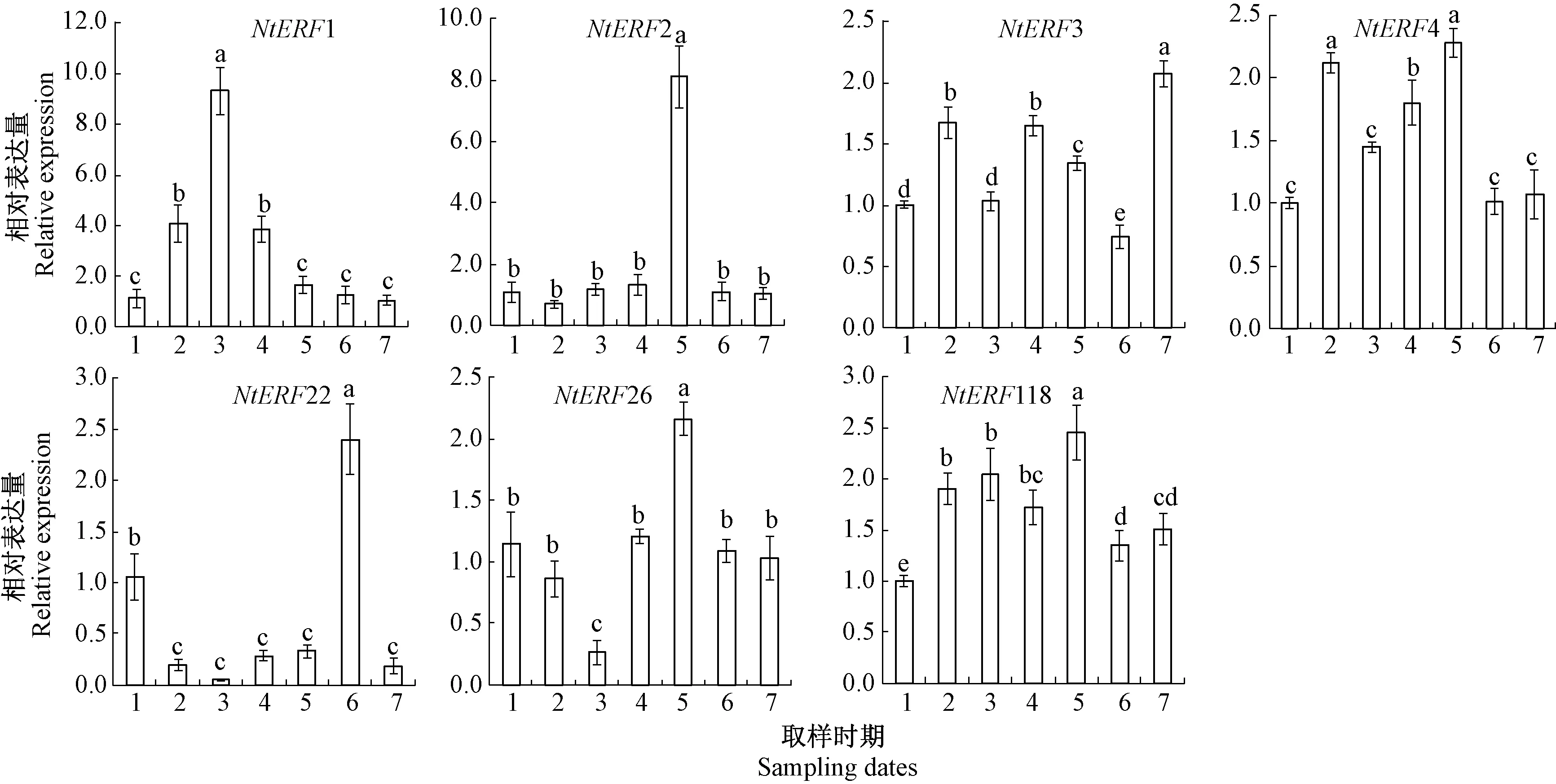
图5 水仙膨大期内芽NtERFs基因的表达量变化
3 讨论
研究表明,乙烯抑制植物的生长,如在根系发育早期,乙烯抑制了根系伸长[18-20]。然而,根据Pierik等[21]提出的双相乙烯反应模型,乙烯对根系伸长既有抑制作用又有促进作用。Abts等[22]研究表明在甜菜根发育早期,生长素促进乙烯的合成,从而促进了根的生长发育,这可能是由于乙烯对木质部的生长起到促进的作用[23],外源乙烯的处理证实了乙烯能够刺激形成层和不定根的生长[24-25]。对乙烯敏感型的番茄,乙烯能够影响其营养生长,在乙烯缺乏的情况下,番茄成年植株显示出与乙烯相关的典型变化,如叶片变白、花期提前和果实脱落加速等[26]。乙烯也能影响鳞茎的生长发育。在洋葱鳞茎分化过程中,乙烯作为鳞茎形成的直接诱导物质参与其中[27]。方少忠等[3]研究表明乙烯可诱导百合试管鳞茎的形成和膨大。本研究发现内芽乙烯的释放高峰出现在4月下旬,在同一时期种球处于迅速生长阶段,说明乙烯可能参与了水仙鳞茎的发育生长过程,这与前人的研究结果一致。
ERF蛋白存在于表现出乙烯三重反应的植物中,与乙烯的生物合成和信号传导有关[28]。ERFs是乙烯信号传递途径的最后一个下游受体,其传递的信号与ERF基因的转录激活有关[29],且与乙烯调控的生长控制有关,具有正调控和负调控功能[30]。前人研究表明,ERF蛋白通过调控乙烯生物合成基因的表达参与乙烯生成的反馈调节[31]。抑制ERF蛋白的活性能够下调氨基环丙烷-1-羧酸合成酶(Aminocyclopropane-1-carboxylic acid synthase,ACS)和氨基环丙烷-1-羧酸氧化酶(aminocyclopropane-1-carboxylic acid oxidase,ACO)基因的表达量,减少乙烯生成量[26]。ERF1是ERF家族成员之一,含有ERF DNA结合区域[32]。Yu等[33]研究发现ERF1正调控乙烯信号,与生长发育呈正相关。本研究通过分析NtERF1的表达规律,推测该基因可能在鳞茎发育的过程中也起到正调控作用。ERF2蛋白在烟草或番茄中能够激活乙烯合成途径基因的表达[34],过表达SlERF2基因能够引起乙烯的释放,导致番茄(Solanumlycopersicum)果实早熟,刺激种子萌发[35],MdERF2能够受到乙烯的调节而在苹果(Malusdomestica)果实中特异表达[36],但MdERF2是通过抑制MdACS1基因的表达而负调控乙烯的生物合成[31]。相反,MdERF3作为MdACS1基因的转录因子诱导其表达,MdERF2和MdERF3相互作用,共同调节乙烯的生物合成[31]。本研究发现鳞片中的NtERF2和NtERF3基因的表达水平也存在着此消彼长的关系,在5月中旬,NtERF2表达受到抑制,NtERF3的表达量显著升高,从而产生大量乙烯,但是在内芽中,NtERF2的表达量则与乙烯的释放量呈正相关。因此,本研究认为NtERF2和NtERF3基因调控乙烯生物合成的功能具有组织特异性,2个基因之间在种球中的相互关系还有待进一步研究。
乙烯通过乙烯信号途径刺激维管细胞的分化[37],也能通过与其他激素的相互作用,刺激细胞的生长[38]。研究表明,ERF3是根冠生长的的重要因子,通过调节参与细胞分裂素信号的基因表达,促进根冠的伸长生长[39],ERF1和ERF4表达受乙烯和生长素的影响,与月季花瓣的生长和脱落有关[40],在木本植物中,ERF118和ERF119是木质部细胞的膨胀和次生细胞壁形成的重要调控因子,参与了茉莉酸和生长素介导的生理反应[41],关于ERF26的功能则鲜见报道。水仙种球中的水杨酸和玉米素与芽的生长有关[42],本研究发现,在种球膨大期间,NtERFs基因能够在鳞片和内芽中响应乙烯信号,但是这些基因的表达是否受到其他激素信号的影响还需要进一步研究。
4 结论
本研究结果表明,在多花水仙种球膨大期,种球重量逐渐增加,鳞片释放的乙烯与地上部的衰老有关,内芽释放大量的乙烯,所释放的乙烯与种球的膨大有关,并且能够诱导鳞片乙烯的释放。NtERF1、NtERF4和NtERF118基因可能参与鳞片对乙烯信号的响应,NtERF2、NtERF3、NtERF4和NtERF26基因则可能参与内芽乙烯信号的响应,NtERF2可能是鳞片响应内芽乙烯信号的纽带,在种球乙烯的生物合成中具有关键作用。本研究初步分析了乙烯在多花水仙种球膨大期间的作用,为进一步了解水仙种球的生长发育提供了研究基础,但由于相关研究基础较少,今后应进一步挖掘乙烯信号途径中的相关受体基因,明确乙烯在多花水仙种球生长发育过程中的作用及其信号传递过程。
猜你喜欢
种子(2022年7期)2022-08-27
当代石油石化(2022年2期)2022-03-11
野生动物学报(2022年1期)2022-02-24
江河文学(2021年3期)2021-07-14
作文大王·低年级(2020年4期)2020-04-19
阅读与作文(小学低年级版)(2019年12期)2019-12-26
创新作文·初中版(2019年7期)2019-12-17
当代工人·精品C(2019年1期)2019-04-29
儿童故事画报·发现号趣味百科(2014年1期)2014-03-31
现代农业研究(2009年9期)2009-10-21
