
油茶CYP78A10 同源基因及其SSR 位点
2021-05-27 07:05赵松子
南方林业科学 2021年2期
赵松子
(江西省林业科学院·江西省油茶种质资源保护与利用实验室,江西 南昌 330013)
种子大小是对作物产量具有重要影响的数量性状,拟南芥(Arabidopsis thaliana)CYP78A 亚基因家族基因参与种子大小的遗传控制,该调控机制在植物中似乎是相当保守的。拟南芥有6 个CYP78A 基因,分别 为KLU/CYP78A5、EOD3/CYP78A6、CYP78A7、CYP78A8、CYP78A9 和CYP78A10;CYP78A5 通过增加细胞分裂来促进叶、花和种子的生长,CYP78A5 功能缺失突变体的细胞分裂停止较早,导致花瓣、萼片、叶、茎等器官变小[1-4];CYP78A6 在几乎所有组织或器官遍在性地表达,功能缺失突变体没有突变表型,过表达能增加种子大小[5];CYP78A7 功能缺失突变体似乎具有正常表型,但CYP78A5 和CYP78A7 的双突变体产生胚柄向胚的转换并常在胚期死亡,幸存的植株矮小,并且不能产生种子[2];CYP78A8 和CYP78A9 的双突变体产生种子数量较少[6];CYP78A9 过表达导致果荚变大,只产生少量种子[7],通过RNAi 沉默樱桃(Prunus avium)PaCYP78A9 可以使果实变小[8];大豆(Glycine max)GmCYP78A10(CYP78A51)是CYP78A10的同源基因,野生大豆(G.soja)GmCYP78A10 的野生等位型与大豆的高产等位型间存在3 个SNP,等位型的不同导致种子重量7.2%的差异[9]。
油茶是山茶科山茶属(Camellia)中油用物种的总称,主要包括普通油茶(Camellia oleifera)、小果油茶(C.oleiferavar.monosperma)、攸县油茶(C.yuhsienensis)、越南油茶(C. vietnamensis)、浙江红花油茶(C.chekiangoleosa)、广宁红花油茶(C. semiserrata)、宛田红花油茶(C. polydonta)、腾冲红花油茶(C. reticulata)等[10-11]。油茶种子大小在物种内和物种间均存在明显差异,并且物种间的差异大于物种内的差异。黄勇等调查了小果油茶18 个不同群体,种子长度为12.75~19.52 mm,种子宽度为10.82~16.23 mm[12];马力等调查了11 种山茶属植物,最大的为越南油茶,种子千粒质量为1 878.77 g,最小的为攸县油茶,千粒质量为450.55 g[13]。简单重复序列(SSR)主要是以1~6 个核苷酸为基本重复单位的串联重复序列,利用SSR 差异可以开发SSR 标记,具有多态性高、重复性好及共显性等优点。油茶物种间杂交具有较高的亲和力,如越南油茶×小果油茶的座果率达36.6%[10],物种间种子大小的巨大差异为超高产杂交育种提供了良好的材料,鉴别油茶种子大小等产量性状相关基因,开发与之紧密连锁的SSR 标记,可以开展分子标记辅助育种,有助于实现产量性状相关优良等位基因的聚合,大幅提高油茶产量。
1 材料与方法
1.1 试验材料
从种植于江西省林业科学院院内的油茶良种“赣无1 号”采收成熟种子,室内发芽后采集长约0.5 cm的根尖组织进行转录组分析。
1.2 油茶根转录组分析
采集油茶根尖组织,委托深圳华大基因科技服务有限公司提取RNA 并进行转录组测序,共得到40 529 966 条读段,读长为90 nt,去除的低质数据后得到37 184 398 条读段。经过SOAP 软件的拼接,获得90 698 条Unigene。经与NR、NT、SwissProt、KEGG、COG 及GO 等6 个数据库比对,确定CDS 有63 008 条。
1.3 生物信息学鉴定
利用BLAST 软件,以拟南芥的CYP78A10 蛋白质序列搜索油茶良种赣无1 的幼叶、未成熟种仁、花蕾、根转录组CDS 数据库[14-16],得到同源基因的部分CDS 序列及其蛋白质序列,用Boetie2[17]软件将转录组测序数据单读段比对到CDS 序列,根据读段编号用自编的Perl 脚本提取成对的读段,在默认状态下用Cap3 软件[18]进行序列拼接,获得同源基因的mRNA 序列;再用Boetie2软件将转录组测序数据单读段比对到mRNA 序列,提取成对的读段,用Cap3 软件再进行序列拼接,获得新的mRNA 序列;重复上述过程,直到mRNA 序列不再延伸。
用油茶CYP78A10 同源基因的mRNA 序列搜索油茶良种赣无1 基因组三代测序数据[19],得到含有油茶CYP78A10 同源基因mRNA 序列的亚读段;亚读段经过校正后,用Augustus[20]软件进行基因预测,得到油茶CYP78A10 同源基因的全长CDS 序列。
1.4 系统进化树构建
利用COBALT[21]软件对油茶CYP78A10 同源基因及玉米(Zea mays)、大豆、松树(Pinus radiata)、拟南芥、水稻(Oryza sativa)、苔藓(Physcomitrella patens)等植物中已鉴别的CYP78A 进行蛋白质序列比对,构建CYP78A 蛋白质系统进化树(Neighbor-joining 法)。
1.5 SSR 位点信息分析
用Boetie2 软件将15 个油茶良种(赣无1、赣无2、赣190、赣447、赣70、赣抚20、赣6、赣永5、赣8、赣石84-3、赣石83-4、赣石84-8、赣无12、兴46、赣无15)的GBS 分析数据[22]以及油茶幼叶、未成熟种仁、花蕾、根转录组数据[14-16]比对到油茶CYP78A10 同源基因的基因组序列或mRNA 序列。用自编的Perl 脚本对比对结果进行统计分析,提取成对比对、读段距离小于300bp、仅比对1 次的读段用于搜索SSR 序列。
搜索SSR 序列:用自编的Perl 脚本对油茶CYP78A10 同源基因的基因组序列及提取的GBS 和转录组读段进行SSR 序列搜索;搜索SSR 序列的条件为:单核苷酸的重复次数≥16,二核苷酸的重复次数≥8,三核苷酸的重复次数≥5,四核苷酸的重复次数≥4,五核苷酸的重复次数≥3,六核苷酸的重复次数≥3。
2 结果与分析
2.1 油茶CYP78A10 同源基因
以拟南芥的CYP78A10 基因序列分别搜索油茶幼叶、未成熟种仁、花蕾、根转录组CDS 数据库,各得到1条最佳匹配序列,合计4 条序列,经过序列比对,从花蕾和根中得到的2 条序列为CYP78A10 的同源基因;将转录组测序数据单读段比对到油茶CYP78A10 的同源基因序列,提取成对读段,用Cap3 软件进行序列拼接,获得油茶CYP78A10 同源基因的mRNA 序列。
用油茶CYP78A10 同源基因的mRNA 序列分别搜索油茶基因组三代测序数据,得到2 条亚读段,亚读段经过校正后,长度分别为40817bp(GenBank 登录号:MW143063)、15341bp(GenBank 登录号:MW143061),用Augustus 软件进行基因预测,得到油茶CYP78A10同源基因的全长mRNA 序列、全长CDS 序列及aa 序列,其结果与转录组测序数据的拼接结果基本一致,2个基因分别命名为CoCYP78A10a(GenBank 登录号:MW143062)、CoCYP78A10b (GenBank 登录 号:MW143060),其aa 序列相似度为81.02%。
2.2 植物CYP78A 蛋白质序列进化分析
用油茶CYP78A10 同源基因及其它植物中CYP78A 亚基因家族的29 条蛋白质构建进化树(表1、图1),结果表明,CoCYP78A10a 和CoCYP78A10b 与CYP78A10、GmCYP78A10(CYP78A51)、CYP78A68 在一个分支,是CYP78A10 的同源基因,可能参与油茶种子大小的调控。

表1 系统进化树中的CYP78A 蛋白质及其编号Tab. 1 CYP78A proteins in phylogenetic tree and their accession numbers.
2.3 SSR 位点信息分析
对CoCYP78A10a 和CoCYP78A10b 的基因组序列进行分析,分别搜索到7、15 个SSR 位点(表2)。用赣无1 等15 个油茶良种(表3)的GBS 分析数据与SSR位点的序列进行比对,结果(表3)表明:CoCYP78A10a的7 个SSR 位点在GBS 数据中未检测到;CoCYP78A10b 的15 个SSR 位点中,在GBS 数据中检测到5 个,其中,SSR8、SSR17 和SSR18 具有多态性。SSR18 与CoCYP78A10b 的转录、翻译起始位点分别距离814bp、1009bp;SSR18 的基本重复单位为TTTCT,具有4 种重复序列,即4 个等位位点,分别为:(TTTCT)3、(TTTCT)4、(TTTCT)5、(TTTCT)6,不同等位位点测序深度的比例基本合理。
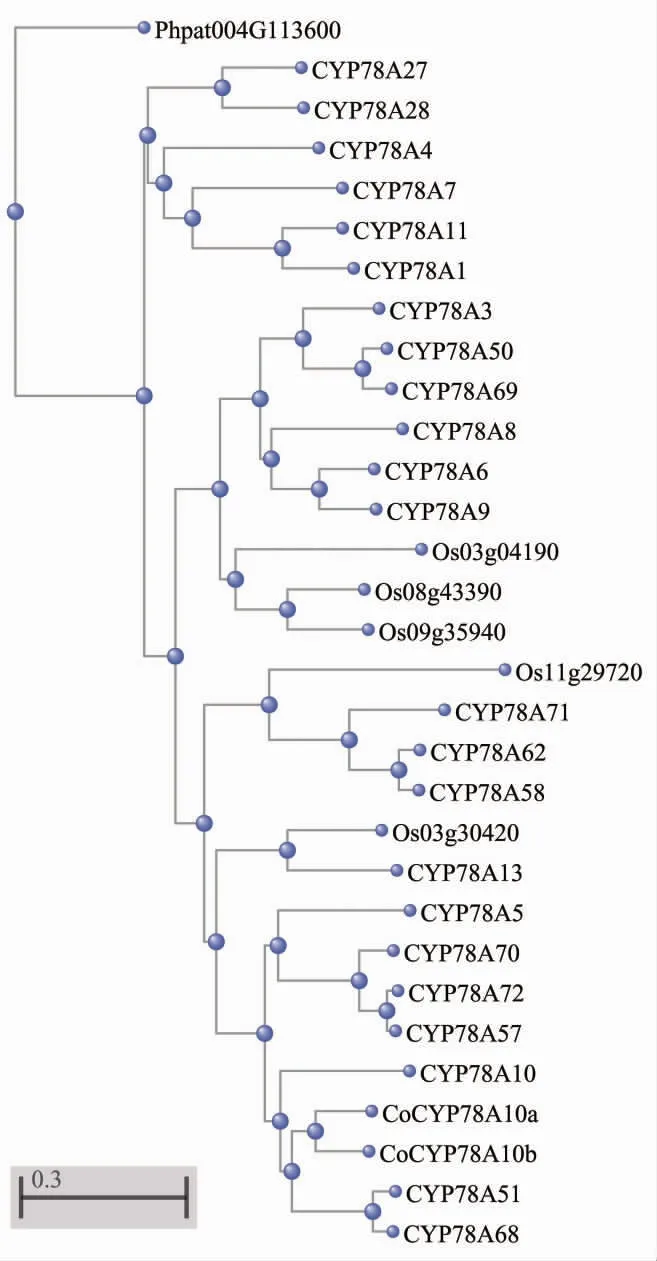
图1 油茶及其它植物中CYP78A 蛋白质的系统进化树Fig. 1 Neighbor-joining phylogenetic tree of CYP78A proteins in Camellia oleifera and other plants
在CoCYP78A10a 的7 个SSR 位点中,SSR7 位于第1 外显子5’端的非翻译区,与翻译起始位点距离35bp,用油茶幼叶、未成熟种仁、花蕾、根转录组数据与CoCYP78A10a 的mRNA 序列进行比对,结果(表4)表明:SSR7 具有多态性;基本重复单位为CT,具有8 种重复序列,即8 个等位位点,分别为:即(CT)9、(CT)10、(CT)11、(CT)13、(CT)14、(CT)15、(CT)17、(CT)18;花蕾中有3 个等位位点,代表油茶良种赣无1 中SSR7 的多态性;根和种仁中分别有5、3 个等位位点,其中5 个等位位点是花蕾中所没有的,由于油茶是异花授粉,根和种仁是赣无1 与其它品系杂交所得,其等位位点代表油茶群体中SSR7 的多态性。
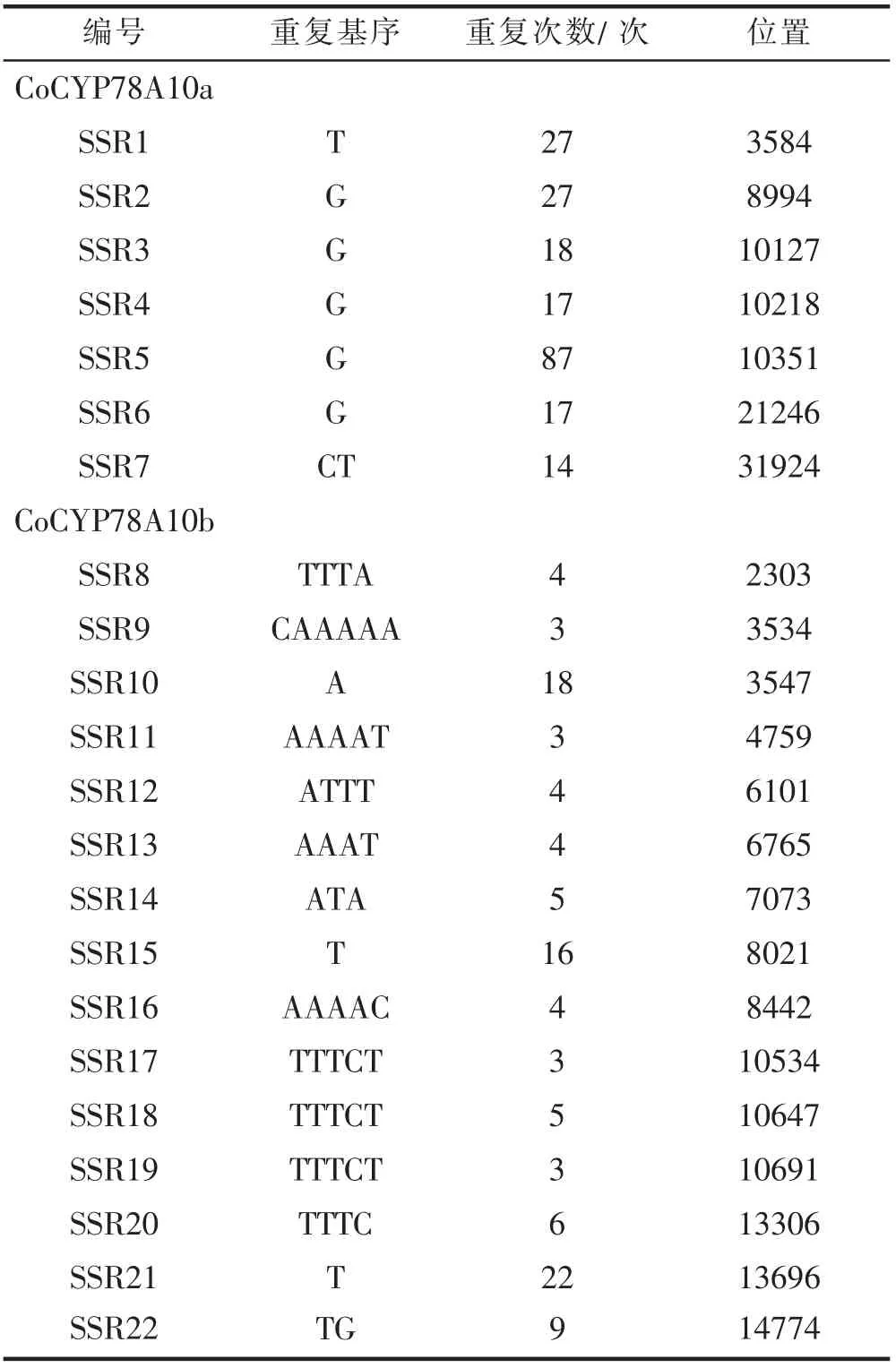
表2 CoCYP78A10a 和CoCYP78A10b 基因组序列中的SSR位点Tab. 2 SSR loci in CoCYP78A10a and CoCYP78A10b genome sequences
3 结论与讨论
本研究从油茶转录组CDS 数据库中鉴定出2 条与拟南芥CYP78A10 基因同源的CDS 序列,从油茶基因组三代测序数据中得到其完整的基因组序列及全长CDS 序 列,并分别命名 为 CoCYP78A10a、CoCYP78A10b。经与其它植物的CYP78A 基因序列进行比对分析并构建进化树,结果表明,CoCYP78A10a、CoCYP78A10b 与CYP78A10、GmCYP78A10 在一个分支,是CYP78A10 的同源基因,可能参与种子大小的调控。
对CoCYP78A10a 和CoCYP78A10b 的基因组序列进行SSR 位点信息分析,分别搜索到7、15 个SSR 位点。用赣无1 等15 个油茶良种的GBS 数据,以及油茶幼叶、未成熟种仁、花蕾、根转录组数据对SSR 位点的多态性进行分析,SSR7、SSR8、SSR17 和SSR18 具有多态性;SSR7 位于CoCYP78A10a 第1 外显子5’端的非翻译区,与翻译起始位点距离35bp;SSR18 位于CoCYP78A10b 的启动子区,与转录、翻译起始位点分别距离814bp、1009bp。
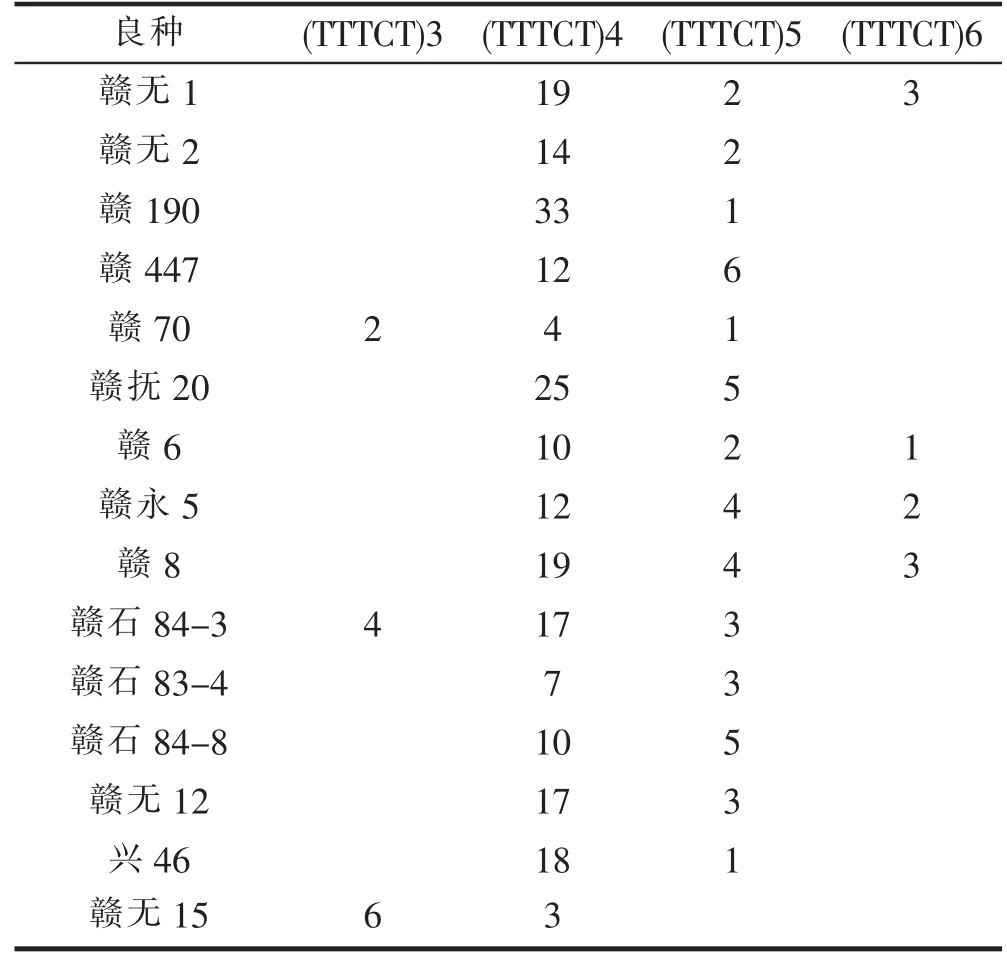
表3 SSR18 等位位点在不同良种中的测序深度Tab. 3 Sequencing depth of SSR18 alleles in different cultivars
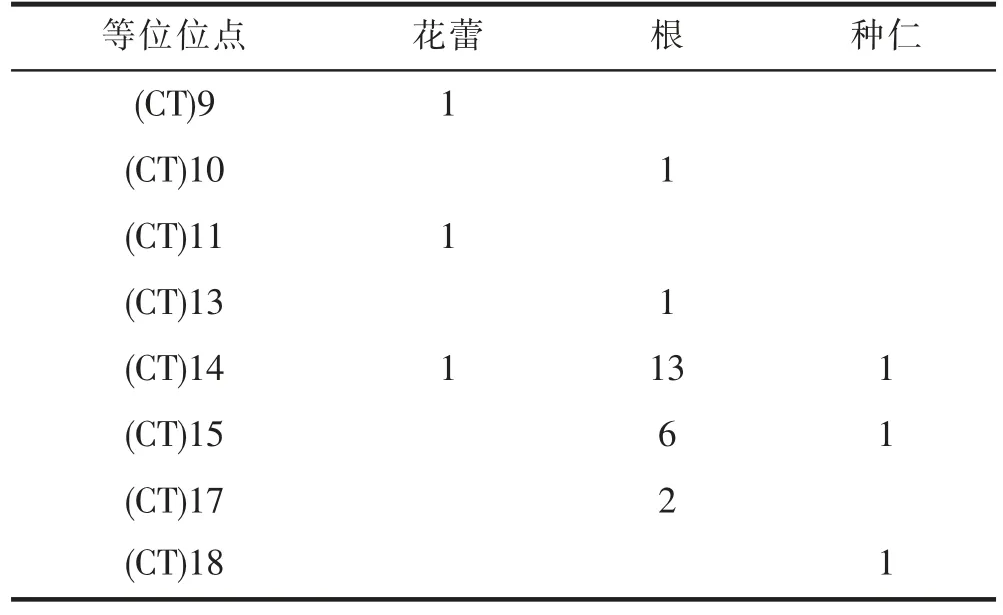
表4 SSR7 等位位点在不同组织中的测序深度Tab. 4 Sequencing depth of SSR7 alleles in different tissues
总体而言,油茶SSR 标记开发比较盲目,成功率比较低,主要原因是没有PCR 产物或PCR 产物没有多态性,如:Jia 等合成了150 对SSR 引物,检测了20 个品种,只有52 对具有多态性[23];李海波等合成了89 对SSR 引物,只有32 对能有效扩增[24];闫蕊等用163 对SSR 引物检测了5 个品种,48 对引物具有多态性[25]。利用油茶基因组数据搜索SSR 位点,结合GBS 数据和转录组数据统计分析SSR 位点的多态性,有利于提高SSR 标记开发的成功率。未来将围绕CoCYP78A10a、CoCYP78A10b 挖掘出更多的多态性位点,并转换成分子标记,以便对油茶种质资源群体进行种子大小的关联分析,研究CoCYP78A10a 和CoCYP78A10b 的功能。
猜你喜欢
今日农业(2022年2期)2022-11-16
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2022年3期)2022-04-19
作物学报(2022年6期)2022-04-08
昆明医科大学学报(2022年2期)2022-03-29
国际医学放射学杂志(2021年5期)2021-10-22
汉字汉语研究(2021年2期)2021-08-30
昆明医科大学学报(2021年2期)2021-03-29
汉字汉语研究(2019年2期)2019-08-27
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
