
果蔬酶促褐变机理的研究进展
2021-06-03 02:39李彩云严守雷王清章
食品科学 2021年9期
李彩云,李 洁,2,3,*,严守雷,2,3,王清章,2,3
(1.华中农业大学食品科学技术学院,湖北 武汉 430070;2.环境食品学教育部重点实验室,湖北 武汉 430070;3.湖北省水生蔬菜保鲜加工工程技术研究中心,湖北 武汉 430070)
水果和蔬菜在贮运、包装或加工过程中受损后极易发生变色,这种现象被称为酶促褐变[1],一般发生在新鲜的活组织中,或者至少发生在含有活性酶的组织中。机械损伤后的褐变反应通常发生迅速且剧烈。酶促褐变极大地影响了果蔬外观和风味,使营养物质损失,降低了消费者的接受度,对食品生产商和食品加工业都产生重大的经济影响。据估计,热带和亚热带水果和蔬菜的损失超过50%是由酶促褐变造成的[2]。酶促褐变反应的 基本步骤是酚类化合物在酚酶催化下氧化成邻醌类化合物,随后醌类物质经过一系列氧化缩合聚合形成褐色产物,从而导致褐变发生。发生酶促褐变的3 个必要因素分别为酚类底物、氧气、酶[3]。国内外对食品褐变研究较多,但对于酶促褐变机理的研究比较零散,且不够深入。本文整理归纳了国内外围绕褐变机理进行的多酚氧化酶(polyphenol oxidase,PPO)和酚类底物的相关研究和报道,为酶促褐变机理的系统深入研究提供借鉴与参考。
1 PPO相关研究进展
1.1 PPO分类
PPO是酶促褐变反应的重要影响因素和必备条件,PPO是含有Cu2+的酶。国际生物化学和分子生物学联合命名委员会提出,PPO包括3 种类型[4]:第一种是单酚单氧化酶,亦称酪氨酸酶、单酚氧化酶、甲酚酶,这种酶能催化单羟基酚羟基化为邻二羟基酚;第二种是双酚氧化酶,亦称儿茶酚氧化酶、o-双酚酶,这种PPO能催化氧化邻位酚,但不能氧化间位酚和对位酚;第三种是漆酶,该酶能氧化邻位酚和对位酚,不能氧化一元酚和间位酚。现在所说的PPO一般是双酚氧化酶和漆酶的统称[5]。
1.2 PPO催化位点
研究PPO的活性中心发现,铜离子存在于酶的活性中心,大多数PPO活性中心具有两个富含组氨酸(His)残基的Cu结合区域[6]。通过核磁共振等方法得到的数据显示PPO活性中心每个铜离子与3 个His残基中的N原子形成配位键,形成了具有特定三维结构的活性部位,配位键中两个为赤道面配位键,一个为较弱的轴向配位键。这两个铜离子以赤道面与不同数目的氧原子产生配位结合而形成电子通路[7]。PPO因两个铜离子结合氧原子数的不同,产生以下3 种型态:1)含两个与一对外来氧原子连接的二价铜离子的氧化态酶;2)含两个与一个外来氧原子连接的铜离子的还原态酶;3)含两个没与任何氧连接的相互分离的一价铜离子的脱氧态酶[8-9]。氧化态酶去氧生成还原态酶,还原态酶脱氧生成脱氧态酶,而氧化态酶和脱氧态酶通过释放和结合氧气又可相互可逆转化。这3 种型态的存在和相互转化是其发挥其单酚酶和多酚酶催化能力的关键[10]。图1是酪氨酸酶通过3 种型态变化发挥双重催化的一种得到广泛认可的反应模型[11]。
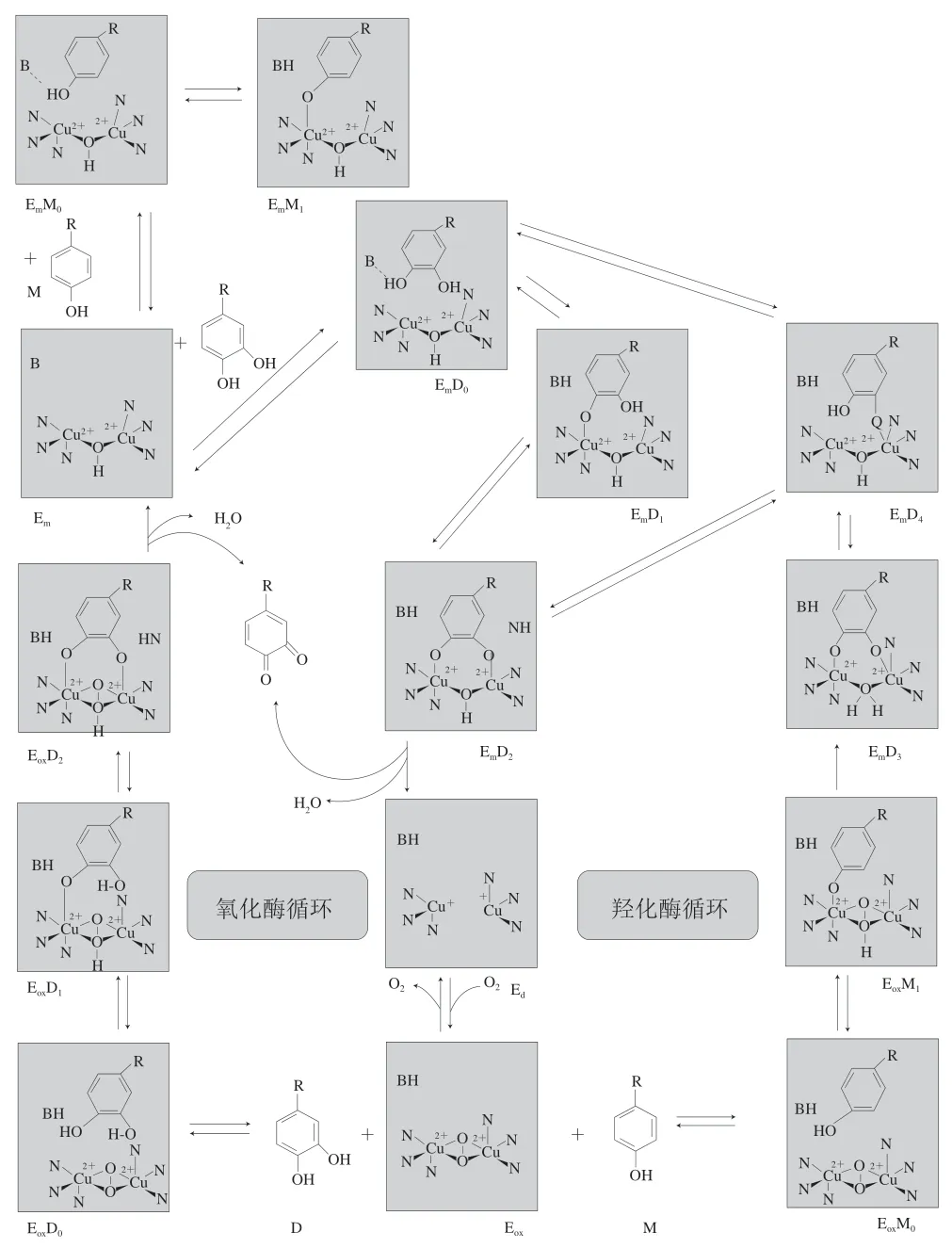
图 1 PPO催化单酚和双酚底物氧化的循环图[11]Fig. 1 Catalytic cycle for the oxidation of monophenol and diphenol substrates by PPO[11]
1.3 PPO提取、分离、纯化、纯度鉴定、活性测定的方法
PPO提取方法很多,通常根据PPO存在的植物种类来选择分离提取方法。从果蔬组织中提取分离PPO面临的一个主要问题是酚类底物和酶同时存在。在活组织中,这两种成分在细胞内单独分开存在,但在浸渍、提取和其他加工处理中,它们一旦由于组织细胞破损而接触就会产生醌和其他缩合产物,在提取时要注意去除酚类底物。传统的提取方法主要分为缓冲液提取法[12-14]和丙酮提 取法[15-16],缓冲液提取法应用较多;新型PPO提取方法有超声波辅助提取法。
PPO粗提物大都含有许多杂质,还需要对其进行分离纯化。盐析和色谱层析法等是早期纯化PPO所常使用的 方法。使用硫酸铵进行分级沉淀后,通过透析去除无机杂质,然后通过色谱法进一步分离。色谱方法有凝胶色谱、吸附色谱、亲和色谱、离子交换色谱[17-18]。在各种色谱方法中选择适当的方法取决于酶的类型、杂质、电荷、分子大小和提取物的纯度。最近几年温度诱导相分离、双水相萃取、三相分离技术等液-液萃取的技术也开始用于酶的分离纯化中。
分离纯化出的PPO一般要进行纯度鉴定,为确保纯化完全,一般鉴定方法有层析法、电泳法、高效液相色谱(high performance liquid chromatography,HPLC)、质谱法等。在整个分离纯化过程中要实时监测PPO活性,测定PPO活性的方法主要有分光光度法和氧电极法。最近Pan Tingtiao等[19]发展了一种新的测定PPO的 方法——表面增强拉曼散射法,将该方法用于苹果和马铃薯样品中PPO活性的测定,并与比色法测定结果进行了比较,发现该方法可成功地用于食品样品中PPO活力的测定。表1总结了PPO提取、分离、纯化、纯度鉴定、活性测定的方法及特点。

表 1 PPO提取、分离、纯化、纯度鉴定、活性测定的方法及特点Table 1 Methods for PPO extraction, separation and purification and determination of its purity and activity and their characteristics

续表1
2 酚类底物相关研究进展
2.1 多酚种类
酚类化合物属于植物次生代谢产物,以苯环为基本骨架,以苯环的多羟基取代为特征,具有一系列的结构和功能。酚类化合物根据结构分为酚酸类、黄酮类、 1,2-二苯乙烯和木质素类[36]。根据其碳骨架的不同可进一步细分(表2)。

表 2 根据碳骨架对植物组织酚类化合物的分类[37]Table 2 Classification of phenols compounds in plant tissues based on carbon skeleton[37]
2.2 影响植物组织多酚种类和含量变化的因素
不同品种的植物组织酚类化合物种类与含量差异较大。Khettal等[38]从产于阿尔及利亚的7 个不同品种柑橘叶片中提取多酚化合物,得到各品种水提物中总酚和总黄酮含量分别为68.23~125.28 mg/g和11.99~46.25 mg/g (以干质量计,下同)。Tanriöven等[39]测定了7 个不同品种梨汁中酚类物质的含量,得到绿原酸质量浓度为73.1~249 mg/L,表儿茶素质量浓度为11.9~81.3 mg/L,咖啡酸质量浓度为2.4~11.4 mg/L,对香豆酸质量浓度为0.0~3.0 mg/L。Yi Yang等[40]对13 个品种莲藕的酚类化合物的 含量、分布、组成进行了分析,发现藕肉、藕皮和藕节中总酚的平均含量分别为1.81、4.30 mg/g和7.35 mg/g (以鲜质量计),总黄酮的平均含量分别为3.35、7.69 mg/g和15.58 mg/g(以鲜质量计)。酚类化合物种类在不同部位上有显著差异,藕肉中酚类物质主要由没食子儿茶素和儿茶素组成;藕皮和藕节中酚类物质主要由没食子儿茶素、没食子酸、儿茶素和表儿茶素组成。
种植方式会影响酚类种类及含量。Veberic等[41]采用HPLC法测定了奥地利和斯洛文尼亚11 个有机栽培苹果品种和11 个综合生产苹果品种果皮和果肉中酚类化合物的含量,发现有机栽培苹果品种与综合生产苹果品种的 果皮酚类物质含量无显著差异;然而,有机种植的苹果果肉中酚类物质的含量高于综合生产的苹果品种。
提取方式也会影响测出的酚类种类和含量。Delgado-Torre等[42]对18 个不同葡萄品种的水醇提物进行了比较研究,分别采用过热液体提取法、微波辅助提取法和超声波辅助提取法进行提取,与其他两种方法相比,过热液体提取法具有更高的提取效率。Mai 等[43]采用多层逆流色谱法和制备型HPLC从卷心莴苣中分离出了10 种酚类化合物(咖啡酸、绿原酸、菜豆酸、菊苣酸、异绿原酸、木犀草素-7-O-葡萄糖醛酸、槲皮素-3-O-葡萄糖醛酸、槲皮素-3-O-半乳糖苷、槲皮素-3-O-葡糖苷和槲皮素-3-O-(6-丙二酰基)-葡糖苷)。紫丁香苷首次在莴苣中被鉴定,这种多酚类物质以前在菊科中没有提到过。
不同成熟期也会影响酚类含量。Lachowicz等[44]在大蒜的叶、根、茎和蒜根提取物中鉴定出了32 种山柰酚衍生物和4 种咖啡酸衍生物。3月份大蒜总酚含量为10.9~1477 mg/100 g,6月份大蒜总酚含量为6.5~1195 mg/100 g。甜菜叶在不同的成熟期褐变程度也不相同,甜菜叶从成熟3 个月到成熟8 个月,总酚含量从4.6 mg/g(以鲜质量计)增加到9.4 mg/g(以鲜质量计)。成熟8 个月的甜菜叶提取物的PPO活力是成熟3 个月叶提取物的6.7 倍[45]。Mahayothee等[46]研究了泰国两个种植区授粉后180、190 d和225 d椰子水和椰肉中的酚类物质,结果发现总酚含量在授粉后180~190 d随椰子成熟而增加,在授粉后225 d下降或保持不变。儿茶酚和水杨酸是椰子水中的主要酚类化合物,而没食子酸、咖啡酸、水杨酸和对香豆酸则存在于椰肉类中。
果蔬的酚类化合物含量由众多因素决定,如品种、成熟期和环境因素。而且酚类化合物在植物中的分布也有不同,这是由遗传和外界因素引起的[41]。HPLC法以及液相色谱-质谱联用在果蔬中酚类物质的分离、定量和鉴定方面取得了长足的进展[47]。
3 酚类化合物的酶促氧化
酶促褐变发生的确切途径一直是人们关注的焦点。现在普遍认为涉及两个反应步骤[48]。第一个反应是单酚羟基化为二酚,邻二酚羟基化为醌。酶促氧化的主要产物醌具有不同的光谱特性,其颜色取决于酚的来源和pH值。例如,酶促氧化后,绿原酸经氧化后呈暗橙色,最大吸收波长为420 nm;儿茶素经氧化后呈亮黄色,最大吸收波长为380 nm;L-多巴经氧化后呈粉红色,最大吸收波长为480 nm[49]。Rouet-Mayer等[50]研究了苹果破碎组织中的酶促褐变以及在(+)-儿茶素和绿原酸的纯溶液中酶促褐变,由二次反应形成的化合物可知,(+)-儿茶素形成的邻醌颜色更深,在绿原酸溶液和破碎的苹果中颜色则更浅。儿茶素的邻醌和绿原酸的邻醌都不稳定,较高pH值时,不稳定性增加。在pH值为4的条件下,绿原酸的邻醌比儿茶素的邻醌显色更强烈。
酚类化合物在发生氧化的同时还会发生自身聚合。酚类物质的氧化聚合对果酒(白葡萄酒、苹果酒等)的口感和稳定性均有影响。张影陆等[51]以黄酮类物质的典型代表儿茶素和酚酸类物质的典型代表绿原酸为研究对象,在模拟体系中通过液相色谱-质谱对其酶促氧化聚合进行了研究。发现单酚在酶促反应过程中确实会发生聚合,且不同的单酚表现出不同的聚合过程及不同的聚合速度。Bernillon等[52]研究了模型溶液的氧化产物与氧化苹果汁的氧化产物,发现了咖啡酰奎宁酸(caffeoylquinic acid,CQA)二聚体、CQA和儿茶素的二聚体以及CQA和二聚黄烷-3-醇的二聚体,未检测到CQA和二氢查耳酮的二聚体。
酚类化合物可以通过不同途径氧化,氧化产物相似。Makris等[53]研究了蘑菇PPO、辣根过氧化物酶和铁氰化钾3 种处理对槲皮素的氧化作用,发现槲皮素溶液经PPO、辣根过氧化物酶、铁氰化钾处理后得到的色谱图非常相似,得到的产物相同。
3.1 醌的生成途径及其次级代谢
3.1.1 醌的不同生成途径
结合不同的分析方法对确定含有一种或两种酚类的模型体系的氧化产物时特别有用。研究者将化学氧化和酶催化氧化均用于这些体系中试图阐明氧化机理[54]。不管醌是通过酶促氧化还是非酶促氧化形成的,这些可能导致形成色素的反应都是相似的[55]。这两个反应途径中唯一区别是褐变的发生速率,酶促褐变通常更快。Munoz-Munoz等[56]通过动态研究PPO、过氧化物酶以及化学氧化对绿茶儿茶素的作用,并且研究了这些化合物在自氧化过程中以及其邻醌生成过程中过氧化氢的形成。PPO/O2和过氧化物酶/过氧化氢氧化绿茶儿茶素分别产生了邻醌和半醌,(-)-表儿茶素和(+)-儿茶素在有O2参与的自氧化过程中过氧化氢生成量很低,而高碘酸盐氧化产生的邻醌的量很大,这突出了邻醌在反应过程中的重要性。
3.1.2 醌的耦合氧化反应
酚类化合物氧化形成不稳定的产物醌后,这些醌会与其他化合物反应,导致醌的稳定性以及由此产生的任何颜色的色调和强度都变化很大,这取决于酚类前体和化学环境。醌的反应分为与酚类化合物反应(图2)和与非酚类化合物反应(图3)两种。

图 2 邻醌与酚类化合物的反应[37]Fig. 2 Reactions of o-quinones with phenolic species[37]
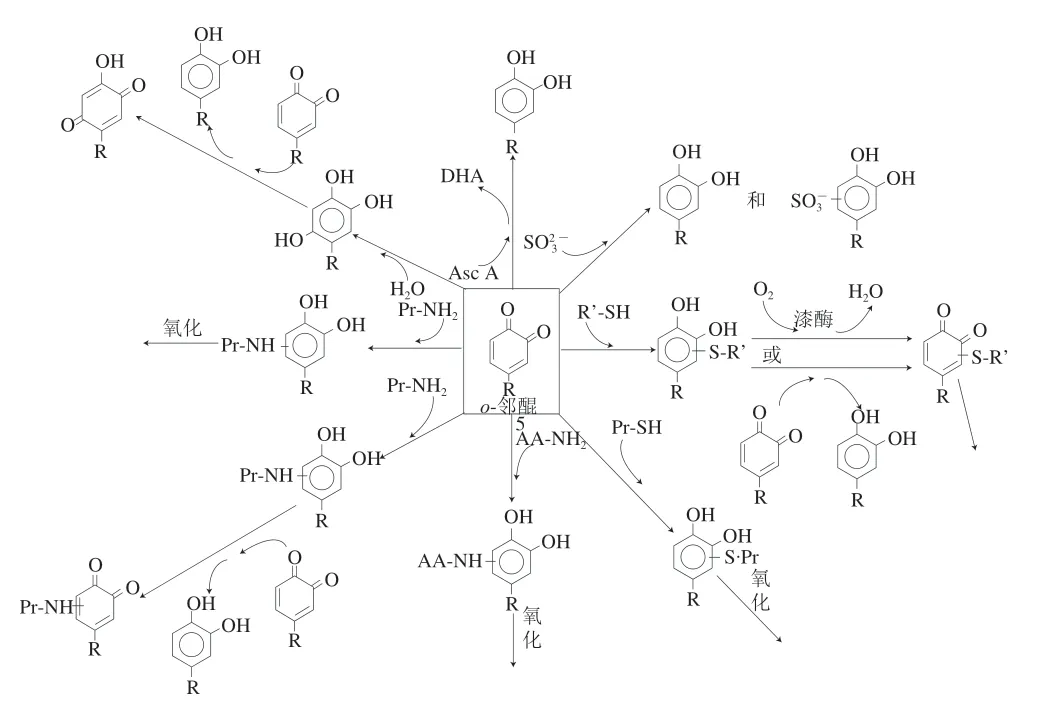
图 3 邻醌与非酚类化合物的反应[37]Fig. 3 Reactions of o-quinones with nonphenolic species[37]
3.1.2.1 醌与酚类化合物的耦合氧化
醌与酚类化合物的耦合氧化可以非常迅速,并且取决于存在的不同醌/酚对应的各自氧化还原电位[37]。这些反应导致原始苯酚的二聚体形成或苯酚的再生(图2)。产物本身经受酶或其他酚类化合物的进一步氧化,导致形成分子质量较大的低聚物。Cheynier等[57]研究了葡萄汁中酚类氧化和褐变的机理,具体为:咖啡酰酒石酸酶促氧化为 相应的咖啡酰酒石醌,然后用谷胱甘肽捕获后者;过量的 咖啡酰酒石醌可能会与黄烷醇和花青素一起进入氧化反应。Poupard等[58]在苹果汁模型溶液中研究了(-)-表儿茶素结构对绿原酸邻醌的反应性(即邻醌是否能够与多酚反应形成复合物)。考虑(-)-表儿茶素分子的两个酚部分的反应性,选择间苯三酚和4-甲基邻苯二酚分别代表A-环和B-环。从形成的氧化产物可以看出,A环和B环的反应性明显不同。A环和B环都可能参与共价键的形成,但是电子转移只发生在B环上。大多数(-)-表儿茶素氧化产物通过A/B环键(“头对尾”分子间偶联)连接。
在PPO、酚类底物和花青素存在的情况下,酶和底物反应产生的中间产物邻醌能通过氧化还原作用将花色苷转化为氧化的花色苷及降解产物。Ruenroengklin等[59]证明了荔枝PPO不能直接氧化荔枝花色苷,而是直接氧化(-)-表儿茶素(荔枝内源性底物),(-)-表儿茶素的氧化产物再催化荔枝花色苷降解,最终导致褐变反应,这可能是荔枝果实采后果皮褐变的原因。Sarni等[60]在模型溶液中研究了咖啡酰酒石酸和葡萄PPO对邻二酚(如花青素-3-葡萄糖苷)和非邻二酚(如锦葵花素-3-葡萄糖苷)花青素的氧化降解作用机理,具体为:咖啡酰酒石酸先经过葡萄PPO酶促反应形成咖啡酰酒石酸邻醌,咖啡酰酒石酸邻醌再与两种花色苷进行反应,花青素-3-葡萄糖苷的降解主要是通过与咖啡酰酒石酸邻醌耦合氧化,而锦葵花素-3-葡萄糖苷与咖啡酰酒石酸醌形成加合物。
3.1.2.2 醌与非酚类化合物的耦合氧化
第二种反应(图3)包括邻醌与非酚类化合物反应,即抗坏血酸、亚硫酸盐、硫醇、伯胺、仲胺(游离或存在于蛋白质中)和水[61]。抗坏血酸是常见的抗褐变剂,它可以通过两种不同的机制防止褐变[62]:在没有PPO底物的情况下,它使PPO不可逆地失活,可能是通过与其活性位点的结合;在存在PPO底物的情况下,抗坏血酸会还原PPO氧化的反应产物,即与氧化形成的醌发生耦合氧化,使醌被还原,从而导致苯酚的再生,从而抑制褐变[61],直到抗坏血酸被耗尽为止,随后会形成棕色。Narváez-Cuenca等[63]研究了亚硫酸氢钠对马铃薯提取物酚类化合物的影响,发现用亚硫酸氢钠处理马铃薯提取物后,酚类化合物与其反应形成了20 种硫代加合物,其结构已通过核磁共振波谱法和质谱法证实。该研究认为当亚硫酸氢钠用作抗褐变剂时,绿原酸被PPO氧化为o-邻醌,然后o-邻醌与亚硫酸氢钠反应生成磺基-o-二酚,而后者不参与褐变反应。Kuijpers等[64]以蘑菇酪氨酸酶和绿原酸组成的褐变模型为基础对NaHSO3和其他含硫化合物(半胱氨酸、谷胱甘肽、二硫苏糖醇)的抗褐变活性进行了比较,发现半胱氨酸和谷胱甘肽通过与绿原酸的邻醌形成无色加成产物来防止褐变,二硫苏糖醇通过抑制酪氨酸酶活性抑制褐变,而NaHSO3两种途径都有参与,其对褐变具有双重抑制作用。
3.1.2.3 醌与蛋白质的相互作用
醌还可以与蛋白质发生共价相互作用而改变蛋白质的理化性质和消化率。Prigent等[65]研究了绿原酸氧化产物与氨基酸侧链之间的共价相互作用,发现CQA氧化后先形成二聚体形式,二聚体再和赖氨酸、酪氨酸、组氨酸和色氨酸发生相互作用。CQA单体似乎能够与组氨酸和色氨酸反应,而未观察到赖氨酸、酪氨酸、丝氨酸和苏氨酸与CQA单体的相互作用。Schilling等[66]研究了绿原酸醌与两种氨基酸衍生物叔丁氧羰基-L-赖氨酸和N-乙酰-L-半胱氨酸之间的共价相互作用,在pH 7.0的模型系统中,证明了两种衍生物的共价加成产物的形成。氨基侧链与醌类物质的共价相互作用可能有助于降低某些食品蛋白的致敏潜力。
3.2 氧化终产物特性
酶促褐变反应产物具有抗氧化性。许多研究主要涉及天然酚类的抗氧化活性,酶促氧化产物也有抗氧化活性,因为它们仍然具有部分酚类化学结构。Cheigh等[67]研究了儿茶素模型系统中PPO氧化得到的黑色素相关产物的抗氧化活性,发现儿茶素-酶反应产物(catechinenzyme reaction products,CERP)呈不同强度的棕色,在反应的早期获得的CERP显示出比在后期更高的抗氧化活性。CERP的抗氧化活性可以通过其给出氢原子、清除自由基和抑制脂氧合酶的能力来解释。Wong-Paz等[68]用苹果中的PPO提取物氧化苹果中常见的绿原酸和(α)-表儿茶素,以产生苹果汁加工过程中形成的氧化产物,比较其与天然(-)-表儿茶素、咖啡酰奎尼酸母体分子、水溶性VE的体外1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除活性,发现氧化产物显示出比母体分子等物质的量混合物更低的抗氧化活性,但异二聚体显示出比同二聚体更高的抗氧化潜能。
酶促褐变反应产物具有抗诱变性。Seung-Shi等[69]研究了马铃薯多酚类化合物的酶促褐变反应产物(enzymatic browning reaction products,PEBRPs)对抗诱变特性的影响,发现100 mg/kg的PEBRPs未引起小鼠骨髓微核嗜多染红细胞(micronucleated polychromatic erythrocytes,MNPCEs)数量的增加,而苯并[a]芘(B[a]P)处理显著增加了MNPCEs的生成率,在小鼠暴露于100 mg/kgmbB[a]P前12 h给予PEBRPs后,MNPCEs生成率显著降低,说明马铃薯酶促褐变反应产物对B[a]P诱导的MNPCEs的生成有较强的抑制作用。
氧化产物还能抑制PPO活性。le Bourvellec等[70]从各个品种的苹果酒中纯化出聚合度分别为80、10.5和4的苹果原花青素组分,原花青素、绿原酸、(-)-表儿茶素3 种单体以及绿原酸和(-)-表儿茶素的混合物分别与绿原酸的邻醌反应被氧化形成氧化产物,评价了各组分及其氧化产物对PPO活性的抑制作用,结果发现天然原花青素 能够抑制PPO活性,抑制强度随聚合度增加而增加;氧化原花青素的抑制作用为天然原花青素的两倍,绿原酸和(-)-表儿茶素混合物的氧化产物也抑制了PPO活性。
4 褐变抑制剂控制酶促褐变的机理研究
为了抑制果蔬褐变和保持果蔬品质,国内外科研工作者进行了大量广泛而深入的研究。在传统褐变化学 抑制剂中,亚硫酸盐被广泛用作褐变的抑制剂,但它会影响人体健康。因此,亚硫酸盐的替代物如有机酸、巯基氨基酸、酚类物质、天然产物等正在被作为可能的替代物而被深入研究。
许多有机酸是天然的褐变抑制剂,如柠檬酸、水杨酸、苹果酸、曲酸、草酸[71-74]等。其抑制机理为:降低体系的pH值、络合金属离子、钝化PPO活性。抗坏血酸既是有机酸,又是还原剂,其抑制褐变机理是将氧化的醌还原为酚类物质,阻止醌类物质进一步自发聚合形成色素物质,抑制PPO活性,降低氧气含量[75-79]。
半胱氨酸、乙酰基半胱氨酸和还原型谷胱甘肽等巯基氨基酸有很强的抗褐变能力[80-83]。其抑制褐变的机制归纳起来为:一是醌类物质能与巯基化合物形成无色的复合物,中断了醌类物质聚合形成色素物质;二是巯基化合物可通过与PPO活性位点的铜离子不可逆结合而抑制酶活性,或者替代PPO活性位点的组氨酸残基;三是巯基化合物并非阻止PPO氧化酚类,而是阻止酚类的聚合[84]。
绝大多数多酚类物质本身能通过疏水键和多位点氢键与蛋白质发生结合,其多个邻位羟基可以和金属离子发生络合反应,且酚羟基中的邻位酚羟基极易被氧化,因此可以作为良好的抗氧化剂。越来越多植物组织中的多酚类提取物被证实对PPO活性具有抑制作用[85]。槲皮素、橙皮素、芹菜素、矢车菊素-3-O-葡萄糖苷都对PPO活性有较好的抑制作用[86-87]。Si Yuexiu等[88]报道橙皮素作为PPO的一种竞争型抑制剂,能够与PPO活性位点周边的His61、His85、His259氨基酸残基发生相互作用。熊志强[87]研究发现龙胆酸、芹菜素、矢车菊素-3-O-葡萄糖苷3 种多酚类物质与PPO通过非共价键和共价键的结合方式,阻断了底物邻苯二酚的进入,从而抑制PPO的催化活性。
豌豆发酵液、大豆发酵液、洋葱、萝卜、甘草的提取物等天然产物都对褐变有很好的抑制效果[89-91]。发挥其抗褐变作用的是提取物中的抗氧化物成分,比如有机酸、多酚类、黄酮类物质,其作用是保护酚类不被氧化,抑制PPO活性。周丽等[90]研究发现豌豆发酵液对马铃薯PPO活性具有良好的抑制效果,发酵液的主要成分为氨基酸、蛋白质、多糖、有机酸、多酚类、黄酮类、皂苷等物质,其中的抗褐变成分具有良好的热稳定性和酸碱稳定性,分子质量小于1000 u,为极性较大的物质。
5 结 语
酶促褐变机理研究对于控制新鲜果蔬的保鲜和加工品质具有重大意义。根据褐变机理,可以从抑制PPO活性、改变酶环境方面入手,或者从阻断酚类底物的聚合、抑制醌氧化、调节醌还原角度出发,减缓和抑制 褐变的发生。但是目前酶促褐变机理研究还存在一些瓶颈,首先,尽管已有大量研究表明PPO参与酶促褐变,但酶晶体的获得还是较为困难,目前对PPO高级结构的研究是以核酸序列、蛋白质序列为基础,利用计算机软件来模拟PPO空间结构、活性位点、底物结合区,其真实的高级结构至今仍未探明。其次,对酚类化合物复杂的代谢过程分析仍有难度,酚类化合物的种类越多,反应产物就会越复杂。再者,大多数研究通过模型体系来探讨酶促褐变机理,在真实的食品体系中研究较少,数据的获得和研究方法的可行性尚需进一步探讨。
随着基因组学技术越来越成熟,可以从基因水平研究PPO,并通过基因编辑、基因敲除、基因沉默等手段对PPO进行调控。随着代谢组学技术的更新,对果蔬中各种类型的酚类化合物在酶促反应中的代谢途径、初级产物、次级产物、反应类型等研究应进一步加强,借助现代先进分析仪器,以真实的食品体系为基础研究酶促褐变中目标成分的含量变化及代谢过程。以基因组学和代谢组学为两大抓手开展PPO和酚类化合物的深入研究将有助于进一步明晰酶促褐变机理。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
当代水产(2021年9期)2021-12-02
食品安全导刊(2021年20期)2021-08-30
中国现代医药杂志(2020年10期)2020-12-14
天然产物研究与开发(2019年1期)2019-03-01
养猪(2019年2期)2019-01-04
中成药(2017年7期)2017-11-22
中成药(2017年7期)2017-11-22
中成药(2017年10期)2017-11-16
中成药(2017年3期)2017-05-17
