
猪圆环4型病毒Cap蛋白的原核表达
2021-08-10 11:35于成东李秋璇李亭玉鲁会军金宁一
中国兽医学报 2021年7期
于成东,哈 卓,王 政,李 凯,李秋璇,孟 媛,李亭玉,于 桐,金 鑫*,鲁会军*,金宁一*
(1.延边大学 农学院,吉林 延吉 133000;2.军事科学院 军事医学研究院 军事兽医研究所,吉林 长春 130122;3.扬州大学 动物医学学院,江苏 扬州 225000;4.吉林农业大学 动物医学学院,吉林 长春 130122)
猪圆环病毒(porcine circovirus,PCV)属于圆环病毒科、圆环病毒属,为二十面体对称、无囊膜的单股、闭合、环状 DNA 病毒[1]。据报道,已鉴定出3种猪圆环病毒,其中包括PCV1、PCV2和PCV3。PCV1是由TISCHER等[1]在1974年于猪肾传代细胞系PK-15中发现并分离,对猪群没有致病性;PCV2最早是由ELLIS等[2]在1991年于加拿大发现的,对猪群有较强的致病性,是引起断奶仔猪多系统衰竭综合征(postweaning multisystemic wasting syndrome,PMWS)的主要病原;PCV3是2016年美国科研人员通过宏基因组测序技术,从患有皮炎与肾病综合征(PDNS)和繁殖障碍的母猪及流产胎儿中鉴定出的猪圆环病毒[3]。
2019年,中国学者从湖南省患有严重临床症状的猪中鉴定出了一种新型圆环病毒,命名为猪圆环4型病毒(PCV4)。PCV4基因组全长1 770 bp,与水貂圆环病毒的同源性最高。对PCV4的2个主要基因进行了分析,其中长度为891 bp的ORF1主要编码复制酶蛋白(Rep蛋白);ORF2长度为687 bp,编码衣壳蛋白(Cap蛋白)[4]。Cap蛋白是主要的结构蛋白,含有多个抗原表位,是病毒的重要免疫原区域。
据报道,中国湖南省、河南省与山西省均检测出PCV4,但在意大利与西班牙并未检测出该病毒[4-6]。PCV4病毒作为一种新发现的圆环病毒,由于发现时间较短,对其存在的时间、感染率、流行情况及致病性并不明确。猪瘟、猪伪狂犬病、猪圆环病毒病并称为世界三大猪病,每年对养猪业造成了巨大经济损失[7],而中国作为PCV4的首次检出地,面临着更加严峻的挑战,对PCV4进行分子流行病学调查,建立有效的抗原、抗体检测方法并对其进行更加深刻的基础研究,对我国养猪业新发PCV的防控具有重要意义。Cap蛋白作为PCV的主要结构蛋白,其在对PCV4的监测与抗病毒方面具有广阔的应用前景。本试验旨在构建pET28a-PCV4-Cap原核表达载体,制备PCV4 Cap蛋白,为建立PCV4血清流行病学检测方法奠定基础。
1 材料与方法
1.1 菌株、载体及主要试剂pET28a载体由本实验室保存;大肠杆菌Trans1 T1、FastPfu Fly DNA聚合酶、Blue PlusⅡ Protein Marker均购自北京全式金生物技术有限公司;表达宿主菌BL21(DE3)PLySs购自生工生物工程(上海)股份有限公司;限制性内切酶BamHⅠ与XhoⅠ,T4DNA连接酶均购自NEB公司;IPTG、DL2000 DNA Marker、λ-EcoT14Ⅰdigest Marker购自TaKaRa公司,质粒小量提取试剂盒购自美国AxyPrep公司。
1.2 基因、引物的设计与合成以本实验室扩增的内蒙PCV4毒株PCV4/CN/NM1/2017(GenBank登录号为:MT882410)的ORF2基因序列为参考,送往生工生物工程(上海)股份有限公司进行ORF2基因合成。应用Premier 5.0软件,设计1对去掉Cap蛋白N端信号肽特异性引物,分别在引物的上下游分别加入BamHⅠ与XhoⅠ酶切位点。上游引物P1:5′-CGCGGATCCCACGCGCGTTTCATGC-GTGA-3′;下游引物P2:5′-CGGCTCGAGAACCCTGTTTCGGGTAGTTAA-3′。
1.3 目的基因的扩增以合成的基因质粒为模板,对目的片段进行PCR扩增,反应体系为:ddH2O 13.5 μL;5× FastPfu Fly Buffer 5 μL;2.5 mmol/L dNTPs 2 μL;;FastPfu Fly DNA聚合酶 0.5 μL;上游引物P1 1 μL(10 μmol/L);下游引物P2 1 μL(10 μmol/L);模板 2 μL,总反应体系为25 μL。PCR扩增程序如下:95℃预变性2 min;95℃变性20 s,60℃退火20 s,72℃延伸30 s,40个循环;72℃延伸5 min。将PCR产物进行1 %琼脂糖凝胶电泳鉴定,对目的片段切胶回收备用。
1.4 重组表达质粒pET28a-PCV4-Cap的构建与鉴定将经PCR扩增后目的片段的胶回收产物与pET-28a载体,使用BamHⅠ与XhoⅠ限制性内切酶进行双酶切,胶回收双酶切产物,将回收的Cap基因与pET-28a载体,使用T4连接酶于16℃条件下连接过夜。将连接产物转化至大肠杆菌Trans1 T1感受态细胞中,挑取单克隆菌落,37℃ 震荡培养12 h,进行质粒小量提取,对所得质粒进行双酶切鉴定,鉴定正确的送往生工生物工程(上海)股份有限公司进行测序,测序结果正确的质粒命名为pET28a-PCV4-Cap。
1.5 重组蛋白的诱导表达将鉴定正确的重组表达质粒转化至BL21(DE3)pLysS 感受态细胞,挑取单克隆菌落,接种于5 mL卡那霉素抗性LB培养基中,37℃震荡培养12 h。以1∶200的比例接种于卡那霉素抗性的LB培养基中,37℃震荡培养至D600达到0.5左右时,加入 IPTG 进行诱导表达,收集菌液进行超声处理。
1.6 重组Cap蛋白的表达条件优化
1.6.1最适表达温度优化 按照1∶200的比例将保存的重组菌接种于卡那霉素抗性的LB培养基中,当D600达到0.5左右时,加入终浓度为1 mmol/L的IPTG进行诱导,于26,37℃条件下分别诱导6 h,收集菌液样本,制备蛋白样,使用12.5%的分离胶进行SDS-PAGE电泳。
1.6.2最适IPTG诱导浓度 按照1∶200的比例将保存的重组菌接种于卡那霉素抗性的LB培养基中,当D600达到0.5左右时,加入终浓度为0.2,0.4,0.6,0.8,1.0 mmol/L的IPTG,37℃诱导8 h,收集菌液,制备蛋白样,进行SDS-PAGE电泳,筛选IPTG最适诱导浓度。
1.6.3最佳诱导时间 按照1∶200的比例将保存的表达菌接种于卡那霉素抗性的LB培养基中,220 r/min 震荡培养至D600为0.5左右时,加入终浓度为0.2 mmol/L的IPTG,于37℃条件下分别诱导2,4,6,8,10,12 h,制备蛋白样,使用12.5%的分离胶进行SDS-PAGE电泳,确定最佳诱导时间。
1.7 重组Cap蛋白的表达形式验证将诱导后的菌液进行收集,5 000 r/inm离心10 min,弃去上清,将沉淀用PBS重悬后,低温条件下进行超声破碎,将破碎后的菌液5000 r/min 离心10 min,收集上清,沉淀用与上清等量的PBS重悬,并取出适量样本,加入5×SDS-PAGE上样液,沸水煮10 min,而后通过SDS-PAGE电泳进行验证。
1.8 Western blot验证将1.5中所得诱导表达的蛋白样本,进行SDS-PAGE电泳后,半干法恒压15 V转至硝酸纤维素膜,5%脱脂乳室温封闭2 h;TBST洗涤3次;使用1∶3 000稀释的鼠源His抗体,4℃孵育过夜;TBST洗涤3次;使用HRP标记的山羊抗小鼠二抗1∶5 000稀释,室温孵育45 min;TBST洗涤3次,每次10 min,而后进行显影。
2 结果
2.1 PCV4Cap基因的扩增使用设计的Cap基因特异性扩增引物,经PCR方法扩增后,进行1%琼脂糖凝胶电泳,最终得到567 bp的目的片段产物,与预期效果相同(图1)。
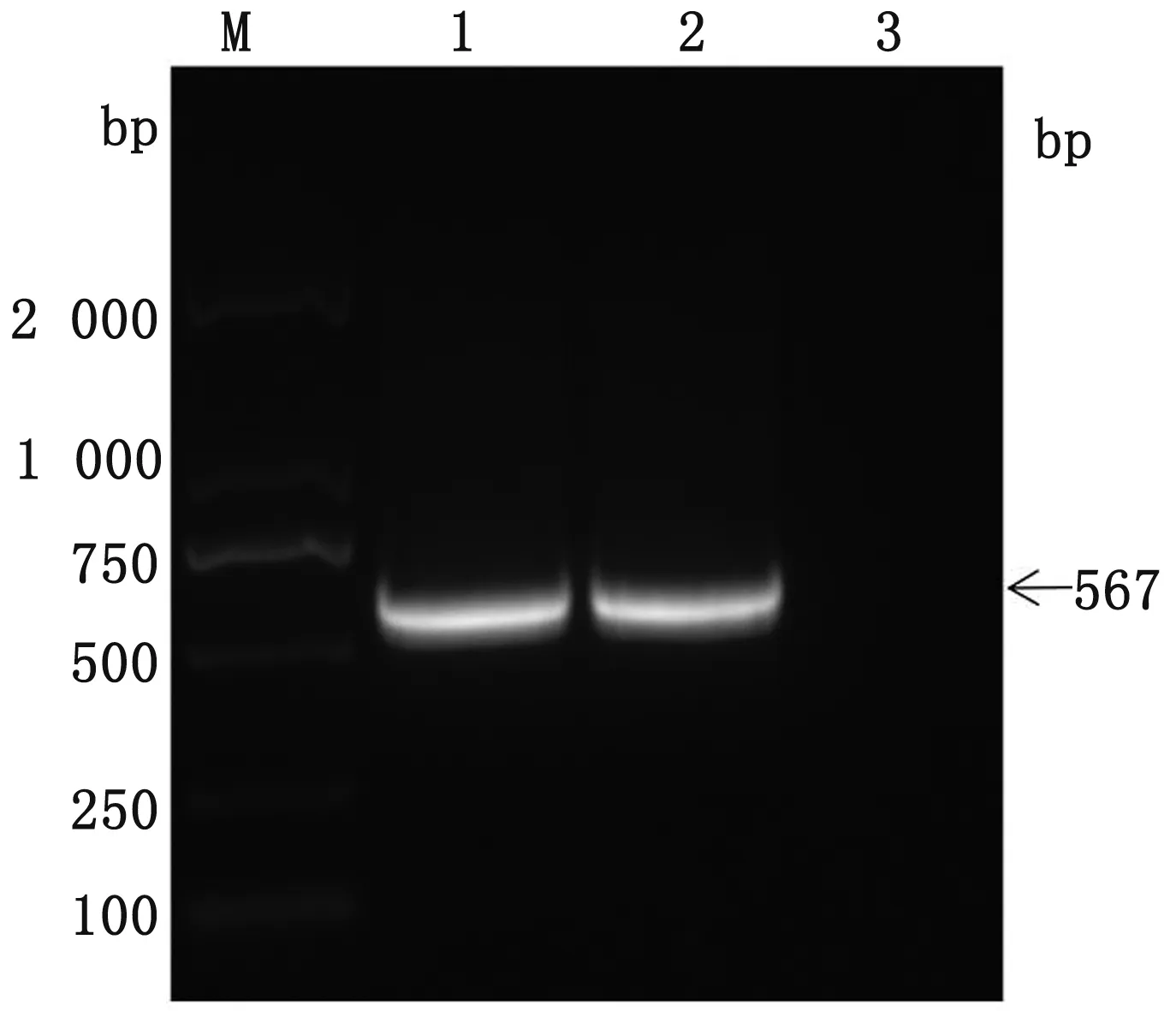
M.DL2000 DNA Marker;1,2.PCV4 Cap基因PCR扩增产物;3.空白对照
2.2 pET28a-PCV4-Cap 重组质粒的构建与鉴定将扩增的PCV4Cap基因片段与pET-28a载体连接,获得重组质粒后,将质粒经BamHⅠ与XhoⅠ限制性内切酶进行双酶切鉴定,结果显示载体与目的基因分别出现在预期位置,PCV4Cap基因成功连接至pET-28a载体(图2)。同时将重组质粒进行测序,测序结果显示,连接至载体的PCV4Cap基因,无碱基突变与缺失。
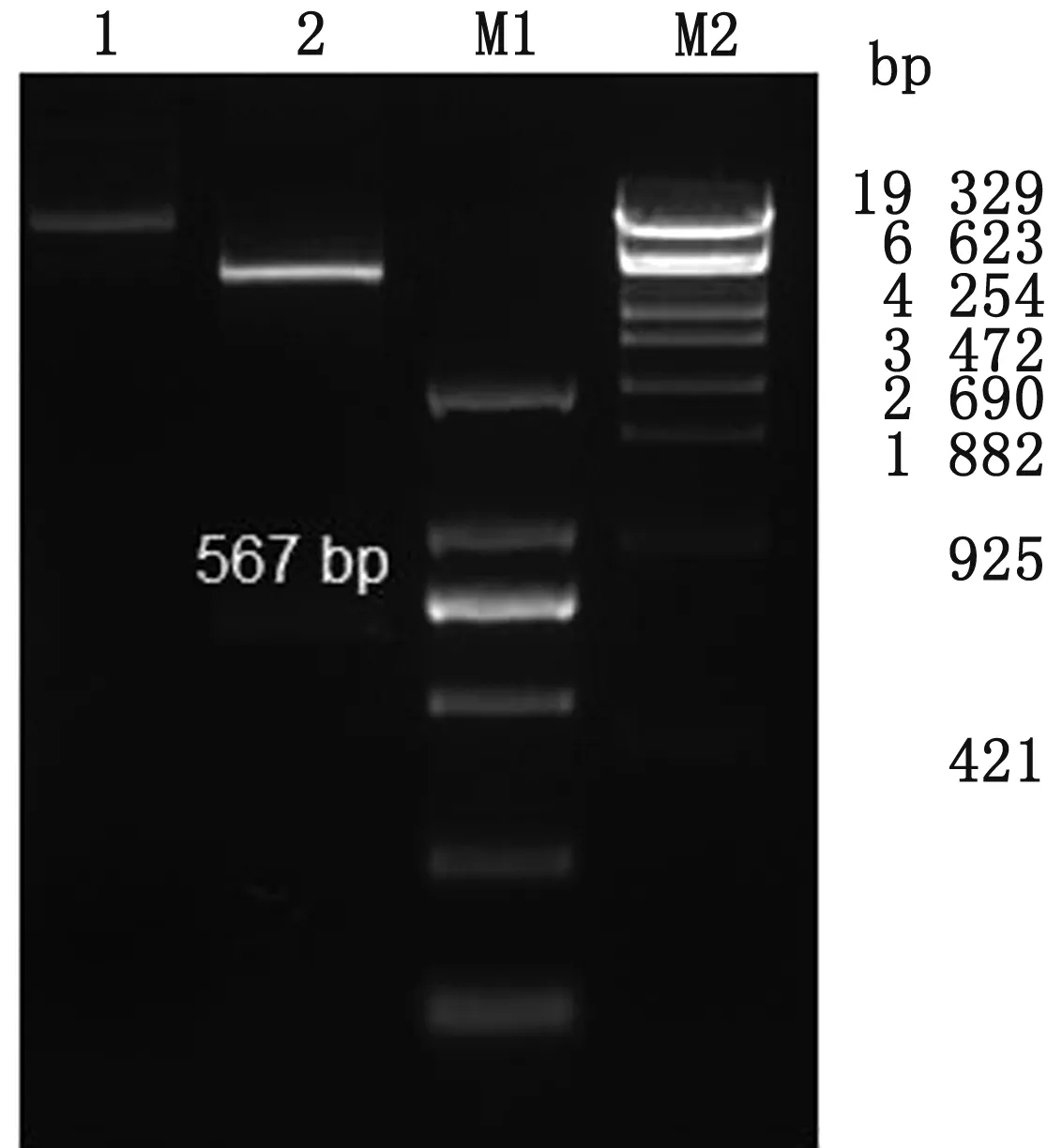
1.pET28a-PCV4-Cap重组质粒;2.pET28a-PCV4-Cap质粒酶切产物;M1.DL2000 DNA Marker;M2.λ-EcoT14 I digest Marker
2.3 pET28a-PCV4-Cap重组蛋白表达条件优化
2.3.1优化最适表达温度 将重组菌分别置于26,37℃条件下诱导表达6 h,SDS-PAGE电泳结果显示26℃几乎没有蛋白表达,pET28a-PCV4-Cap重组蛋白在37℃条件下,表达量更高,更适宜表达(图3)。
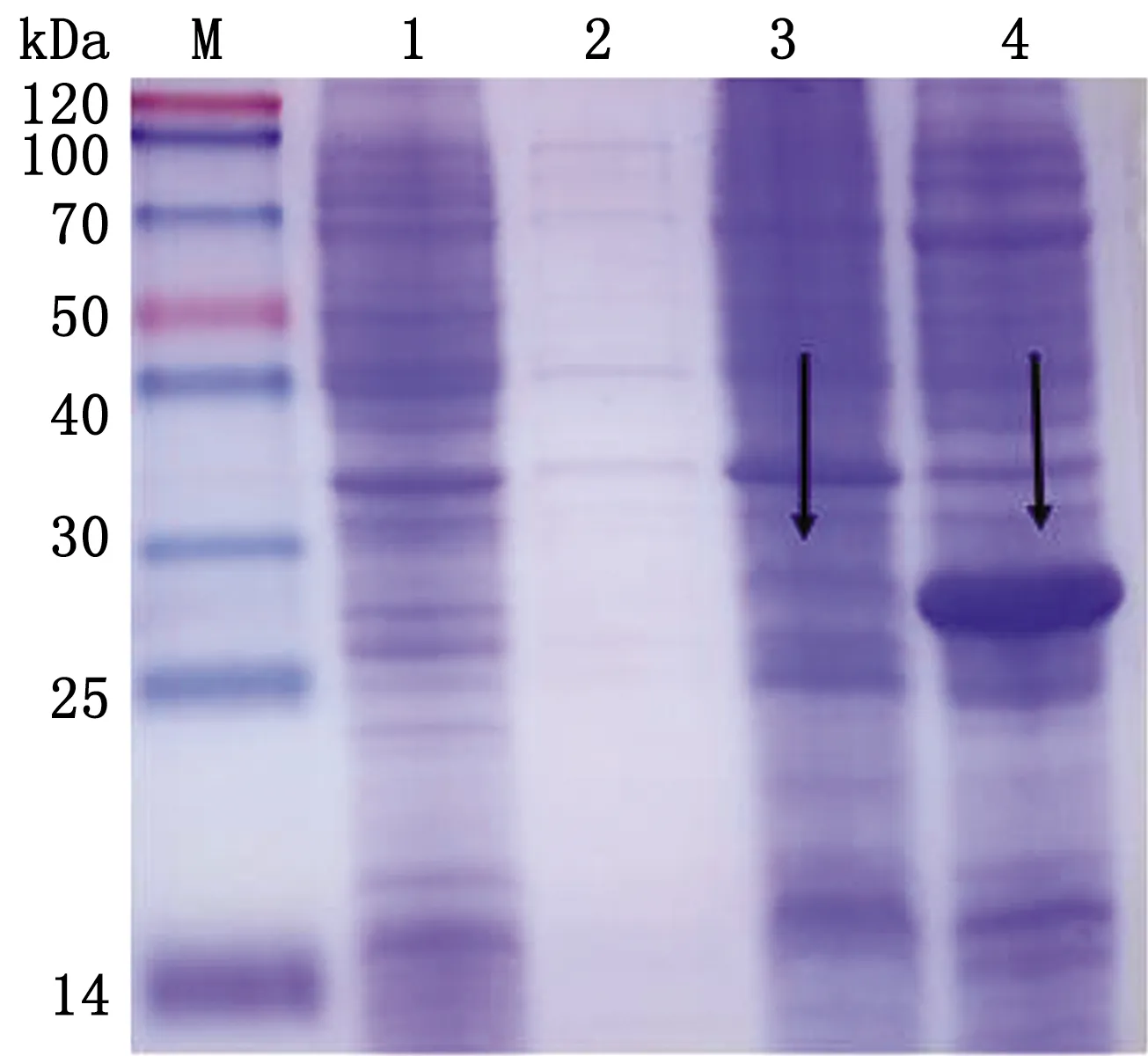
M.Blue PlusⅡProtein Marker;1.pET-28a空载体蛋白;2.重组蛋白诱导前;3.26℃诱导pET28a-PCV4-Cap蛋白表达;4.37℃诱导pET28a-PCV4-Cap蛋白表达
2.3.2优化最佳IPTG诱导浓度 将重组菌于37℃条件下,分别加入终浓度为0.2,0.4,0.6,0.8,1.0 mmol/L 的IPTG进行诱导,结果显示在0.2 mmol/L 的IPTG条件下,pET28a-PCV4-Cap重组蛋白就已经呈现很高的表达量,而后的几个IPTG浓度梯度与0.2 mmol/L浓度IPTG诱导相比并无明显变化。从节约成本的方面进行考虑,故选取0.2 mmol/L的IPTG为最佳诱导浓度(图4)。

M.Blue PlusⅡ Protein Marker;1.pET-28a空载体蛋白;2~7.诱导浓度分别为0.0,0.2,0.4,0.6,0.8,1.0 mmol/L
2.3.3优化最佳诱导时间 于37℃、0.2 mmol/L IPTG诱导条件下,分别在诱导后0,2,4,6,8,10,12 h,取不同时间段取等量菌液,制蛋白样品,通过SDS-PAGE电泳进行比较,发现在诱导后6 h时蛋白的表达量最高(图5)。
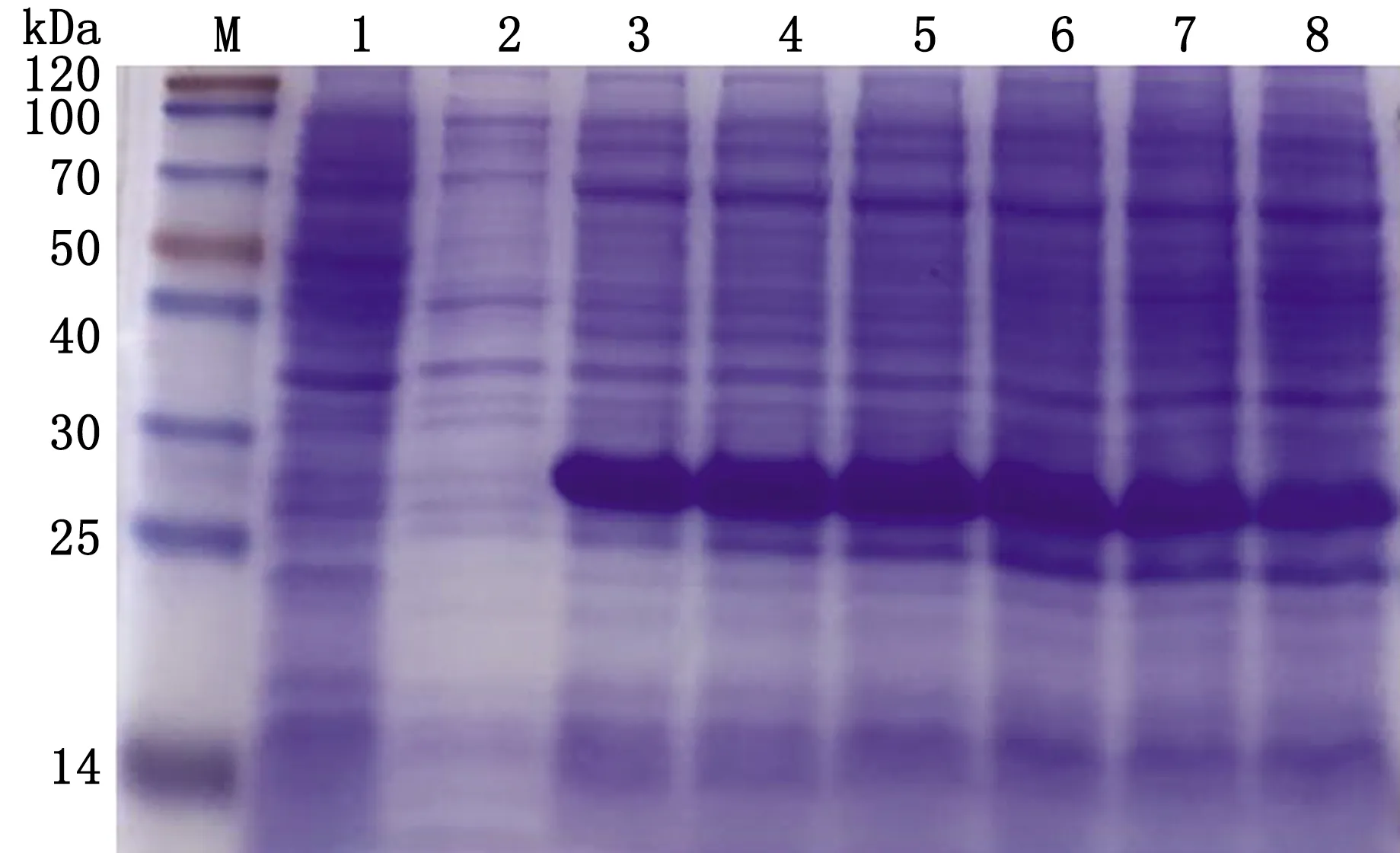
M.Blue PlusⅡ Protein Marker;1.pET-28a空载体蛋白;2~8.诱导时间分别为0,2,4,6,8,10,12 h
2.4 pET28a-PCV4-Cap重组蛋白表达形式验证收集诱导后菌体,5 000 r/min离心10 min后,用适量PBS重悬菌体沉淀,低温条件下超声,使用600 W 功率,超声5 s,间歇7 s,总计15 min。4℃条件下,5 000 r/inm 离心10 min,沉淀用与上清等量PBS重悬,分别取上清与沉淀制备蛋白样后,SDS-PAGE电泳显示,pET28a-PCV4-Cap蛋白均以包涵体形式表达(图6)。
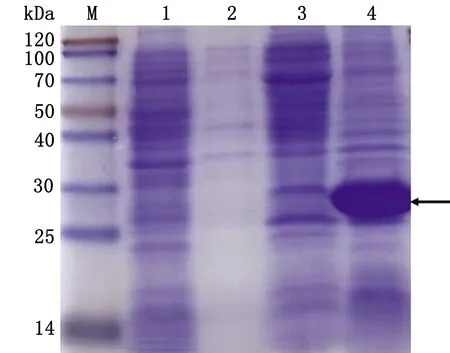
M.Blue PlusⅡ Protein Marker;1.pET-28a空载体蛋白;2.pET28a-PCV4-Cap蛋白诱导前;3.pET28a-PCV4-Cap蛋白超声上清;4.pET28a-PCV4-Cap蛋白超声沉淀
2.5 重组蛋白诱导表达Western blot验证收取诱导后菌液,进行Western blot验证。结果显示,重组蛋白在诱导前具有少量表达,通过诱导后能够出现大量表达,在约27 kDa处出现单一条带,与预期结果相同(图7)。
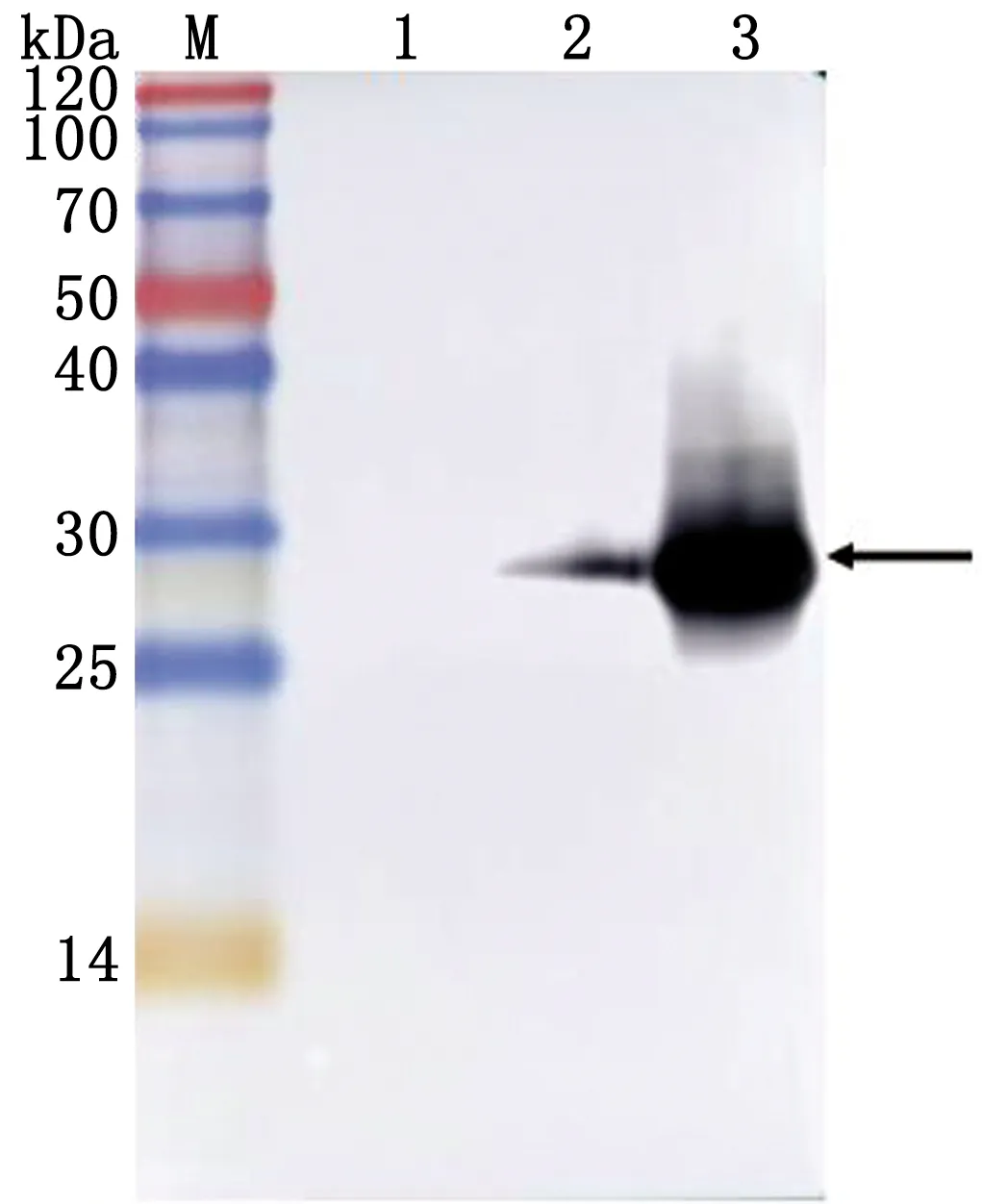
M.Blue PlusⅡ Protein Marker;1.pET-28a空载体蛋白;2.重组蛋白诱导前;3.重组蛋白诱导后
3 讨论
2016年国际病毒分类学委员会更新的圆环病毒科分类标准表明,圆环病毒属内物种划分的标准是全基因组核苷酸同源性小于80%[8]。通过Cap和Rep蛋白完整的基因组及核苷酸序列分析,进一步证明了PCV4属于单独的进化枝,是圆环病毒属中一个独立基因型。通过对PCV4 Cap与Rep蛋白的氨基酸预测,显示其与水貂圆环病毒的同源性最高,而与PCV1、PCV2、PCV3的氨基酸同源性低于50%[4]。PCV1的非致病性与PCV2的致病性已经充分被证实,在有呼吸疾病和腹泻猪中,PCV3检出率和病毒滴度都较高,预示PCV3与这些临床疾病的出现有一定的相关性,且PCV3能够在健康的动物中检测出来[9]。PCV4既可以单独检测,也可以在同时感染了PRRSV或PCV2的猪体内被检测出来。大多数被检测的猪都呈现严重的临床症状,包括呼吸系统疾病、肠炎和猪皮炎与肾病综合征。但是,PCV4 DNA也已在健康猪中检测出来[4]。在2019年中国学者通过感染性克隆,研究了PCV3的致病性,但至今为止也没有可以在细胞中进行稳定传代的PCV3毒株被分离出来。
到目前为止,PCV4的存在时间、感染率、流行情况及致病性并不明确,而Cap蛋白作为PCV的唯一结构蛋白,在病毒的感染与复制中起着至关重要的作用,所以对PCV4 Cap蛋白展开研究,对接下来PCV2、PCV3与PCV4的Cap蛋白之间是否可以提供交叉保护,通过PCV4 Cap蛋白的表达,建立PCV4抗体水平的检测方法,都具有重大意义。
原核表达系统和真核表达系统是当今研究较多的,两者各有优缺点,而目前原核表达系统研究最为透彻,其中大肠杆菌表达系统是外源基因表达的首选,但是表达的外源目的蛋白大多为包涵体[10]。原核表达系统具有试验成本较低、周期短、转化率高且能够获得大量目的蛋白的优点[11],所以本试验选择大肠杆菌表达系统对pET28a-PCV4-Cap重组蛋白进行表达,主要以包涵体的形式表达出约为27 kDa的重组蛋白,并对其表达条件进行了优化。本试验结果显示,加入终浓度0.2 mmol/L IPTG诱导6 h为最优诱导表达条件,重组蛋白表达量最高。
本试验通过成功构建重组表达质粒表达pET28a-PCV4-Cap重组蛋白,为PCV4病毒Cap蛋白的深入研究与PCV4单克隆及多克隆抗体的制备及血清学特异性检测方法的建立提供了依据。
猜你喜欢
数学年刊A辑(中文版)(2022年4期)2022-02-16
中国土壤与肥料(2021年5期)2021-12-02
现代畜牧科技(2021年4期)2021-07-21
现代畜牧科技(2021年5期)2021-07-20
疯狂英语·新悦读(2020年7期)2020-07-30
癌变·畸变·突变(2020年1期)2020-02-12
数学物理学报(2018年5期)2018-11-16
食品科学(2018年10期)2018-05-23
上海蔬菜(2015年2期)2015-12-26
西南医科大学学报(2015年1期)2015-08-22
